Zn2+胁迫对4种草生长及抗逆生理特性的影响
2019-10-25高秉婷吴永贵陈伟
高秉婷 吴永贵 陈伟


摘要:为了筛选耐受锌冶炼废渣及土法炼锌废渣堆场原位修复的先锋草本植物,采用蛭石培养法模拟铅锌废渣在酸雨淋溶过程中释放的锌含量,测定多年生黑麦草(Premier)、高羊茅(Barrera)、草地早熟禾(Barrister)、白三叶(Haifa)4种草的株高、脯氨酸含量及丙二醛(MDA)、超氧化物歧化酶(SOD)指标,分析不同浓度梯度Zn2+胁迫对4种草生长及其抗逆性生理特性的影响。结果表明,Zn2+胁迫会对4种草植株株高、脯氨酸含量、MDA含量、SOD活性产生显著影响(P<0.05),且离子浓度越高,影响越显著。其中,多年生黑麦草、高羊茅和白三叶对Zn2+胁迫耐受性相对高于草地早熟禾,结合从黔西北某铅锌矿渣堆场采样经过改良矿渣的性质,种植4种草验证了模拟试验的结果,因此在锌浓度较高的土壤上或铅锌矿渣场原位生态修复中可以使用多年生黑麦草、高羊茅和白三叶作为先锋物种来进行生态修复的草本种植。
关键词:Zn2+胁迫;重金属污染;生态修复;抗逆性;生理特性
中图分类号:X173 文献标志码: A 文章编号:1002-1302(2019)13-0285-03
土壤是人类赖以生存的重要自然资源之一,也是人类生存环境的重要组成部分。土壤中重金属污染不仅具有隐蔽性、不可逆性等特点,而且经水、植物等介质进入人体,最终影响人类健康[1]。重金属Zn的污染在目前土壤污染中普遍存在。土壤中的Zn主要来自于矿山开采、大气沉降、垃圾和化学药品的残留等,当土壤中的Zn含量超标后,可严重抑制农作物和草坪植物的生长,同时会通过食物链影响家畜和人类健康[2-3]。徐学华的研究表明,重金属对植物产生危害后,植物生理生化特性会发生一系列反应来抵御这种危害[4]。重金属污染是国内外学者研究的重要课题,近年来许多学者开展了对重金属污染的治理研究[5]。重金属污染会导致植株的生理特征发生变化,同时也引起生态系统的恶性循环。当pH值降低到使碳酸盐溶解(如酸雨沉降)时,碳酸盐结合态的重金属容易从废渣堆中释放出来。重金属的可交换态有比其他化学相态更高的溶解性和潜在的生物可利用性。因此,土法炼锌固体废物对环境有潜在影响[6]。草本植物生长周期快,覆盖面大,生命力强,容易种植,易接触和吸收土壤中的金属离子[7],对重金属污染土壤的治理意义重大。本研究采用蛭石培养法探讨不同Zn2+浓度对草地早熟禾(Barrister)、多年生黑麦草(Premier)、高羊茅(Barrera)和白三叶(Haifa)4种草的株高、脯氨酸含量、MDA含量以及SOD活性的胁迫响应,分析了不同浓度Zn2+胁迫对多年生黑麦草、高羊茅、白三叶和草地早熟禾生长及其抗逆生理特性的影响,以期为利用草本植物修复锌冶炼过程产生Zn2+污染矿渣及土法炼锌废渣堆场的原位修复筛选先锋物种提供理论参考。
1 材料与方法
1.1 试验材料及地点
试验材料为多年生黑麦草(Premier)、高羊茅(Barrera)、白三叶(Haifa)及草地早熟禾(Barrister),由西安百绿草业有限公司提供。金属离子:ZnSO4·7H2O为AR级。
試验地点为贵州大学资源与环境工程学院实验室。
1.2 Zn2+溶液配制和浓度设置
Zn2+以溶液形式加入到花盆中,5个浓度梯度分别为0、500、800、1 100、1 400 mg/kg。将草种子播入以蛭石、磷石膏、有机肥等营养物质为基床、直径12 cm的花盆中,待幼苗长出2张展开叶时按照1株/cm2的标准剔除多余幼苗,根据培养基70%持水量每天注入Hoaglands(霍格兰氏)营养液,并将重金属无机盐溶液按梯度施入,当幼苗长出第4张叶片时进行生长发育指标的测定。
1.3 草坪草生长及其抗逆生理特性测定
1.3.1 株高测定 采用游标卡尺测定上述4种草株高(从地面至最高叶尖)。
1.3.2 脯氨酸含量测定 采用酸性茚三酮法[8]测定。取草坪草叶片0.2 g,剪碎后置于试管中,加入5 mL 3%磺基水杨酸,沸水浴提取10 min,冷却后以4 000 r/min进行离心 10 min。吸取上清液2 mL放入另一个带玻璃塞的试管中,加入2 mL冰醋酸和2 mL酸性茚三酮,沸水浴加热30 min。冷却后加入甲苯4 mL,摇匀后静置片刻。吸取上层脯氨酸-甲苯红色溶液于比色皿中,以甲苯为空白,在波长520 nm处比色。根据标准曲线算出2 mL测定溶液中的脯氨酸含量。
单位样品鲜质量中脯氨酸含量=(x×5/2)/(样质量×106)×100%。
1.3.3 MDA含量测定 称取叶片材料1 g左右,加入2 mL 10%三氯乙酸(TCA),研磨后,用TCA定容至10 mL,4 000 r/min 离心10 min,吸取2 mL上清液加入10 mL的具塞玻璃试管中,加入2 mL 0.6%硫代巴比妥酸,沸水浴煮沸15 min,立即冷却离心,分别测定D450 nm、D532 nm值,MDA含量按以下公式计算[9]:
MDA含量(μmol/L)=6.45×D532 nm-0.56×D450 nm。
1.3.4 SOD活性测定 采用氮蓝四唑(NBT)法[10]测定。反应总体积为3 mL,反应以50 mmol/L pH值7.8的磷酸缓冲液作为反应介质,含0.3 mL 130 mmol/L甲硫氨酸、0.3 mL 750 mmol/L 氮蓝四唑(NBT)、0.3 mL 100 μmol/L EDTA-Na2、0.3 mL 20 μmol/L核黄素、0.1 mL酶提取液(空白管加磷酸缓冲液0.1 mL),放于指形管中,反应于4 000 lx光照度下照光10 min(调零管放在暗处),然后在560 nm下进行比色。以抑制NBT光化还原50%时的酶量作为1个活力单位[11]。
1.4 数据统计与分析
运用Origin 9.0整理数据、制作相关图表,用SPSS 24.0软件进行统计分析。
2 结果与分析
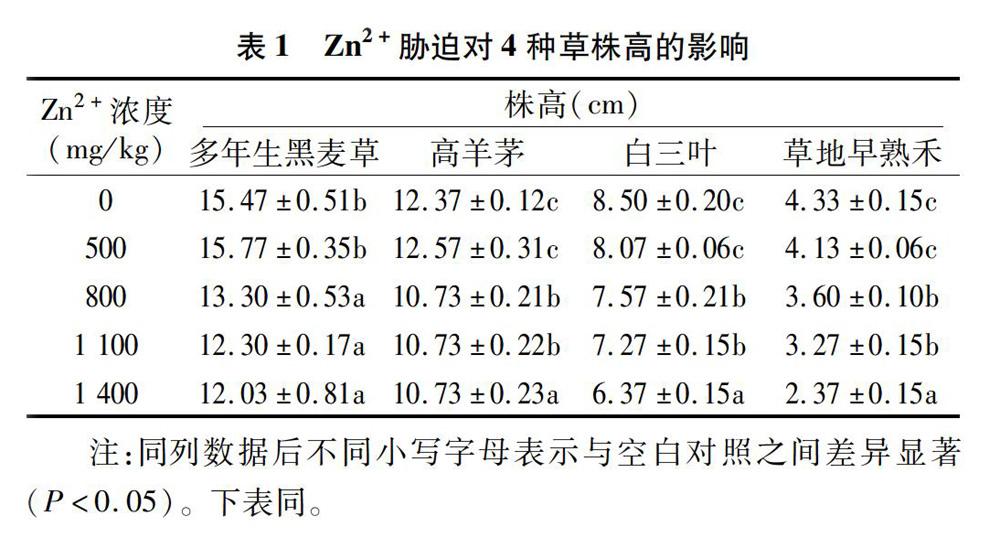
2.1 Zn2+胁迫对4种草株高的影响
从表1可以看出,对4种草来说,除了Zn2+浓度为 500 mg/kg 处理下株高与对照差异不显著外,其余各浓度处理株高均显著低于对照,并且随着重金属离子浓度的升高,株高呈逐渐下降趋势。说明低浓度Zn2+处理对4种草植株的生长影响不大,只有当Zn2+浓度达到800 mg/kg及以上时,会显著影响上述4种草正常生长,造成株高显著低于对照。但对不同草而言,Zn2+胁迫对其株高的影响又不完全一致。对于多年生黑麦草,Zn2+浓度在800~1 400 mg/kg处理下,其株高差异不显著,说明Zn2+浓度达到800 mg/kg甚至更高时,对株高的影响是一定的,没有对生长造成更加不利的影响;对于高羊茅、白三叶和草地早熟禾,Zn2+浓度为800、1 100 mg/kg时,对株高的影响是一致的,当Zn2+浓度达到 1 400 mg/kg 时,对株高影响最大。总体上来说,Zn2+胁迫会对4種草植株株高产生显著影响,且离子浓度越高,影响越显著。但对不同草的影响是不同的,其中多年生黑麦草在株高指标上对Zn2+胁迫的耐受性要高于另外3种草。
2.2 Zn2+胁迫对4种草脯氨酸含量的影响
脯氨酸是植物蛋白质的主要组成成分,在逆境条件下,植物为了减少自身伤害,维持正常生理功能,会在体内积累大量脯氨酸[12]。从表2可以看出,对4种草来说,Zn2+浓度为 500 mg/kg 处理时,其体内的脯氨酸含量与对照差异不显著;Zn2+浓度为800 mg/kg处理时,除草地早熟禾与对照差异显著外,其余3种草都与对照差异不显著;当Zn2+浓度为 1 100 mg/kg 处理时,除多年生黑麦草与对照差异不显著外,其他3种草皆与对照差异显著;在1 400 mg/kg处理时,4种草皆与对照组表现出一定的差异性,除多年生黑麦草外其余3种草的脯氨酸含量均显著高于对照,并且随着重金属离子浓度的不断升高,脯氨酸呈逐渐上升趋势。说明低浓度Zn2+处理对4种草脯氨酸含量影响不大,只有当Zn2+浓度达到800 mg/kg及以上时,会显著影响草正常生长激素的分泌,造成脯氨酸含量显著高于对照。但对不同草而言,Zn2+胁迫对草的脯氨酸含量的影响又不一致。对于多年生黑麦草,Zn2+浓度在500~800 mg/kg处理时,其脯氨酸含量之间差异不显著,说明Zn2+浓度达到500 ~800 mg/kg时,对其脯氨酸含量的影响具有相似性,没有对生长激素的分泌造成不利的影响;但1 400 mg/kg Zn2+浓度处理下时,4种草内的脯氨酸含量之间差异明显;而对于高羊茅、白三叶来说,Zn2+浓度为500~800 mg/kg时,对脯氨酸含量的影响一致,当Zn2+浓度达到 1 100 mg/kg 及以上时,对脯氨酸含量的影响较显著,但当Zn2+浓度达到1 400 mg/kg时,对脯氨酸的含量的影响最大;草地早熟禾在Zn2+浓度为800 mg/kg时表现出显著影响,但当Zn2+胁迫浓度在1 100~1 400 mg/kg之间时对脯氨酸含量的影响极大。总体上来说,Zn2+胁迫会对4种草植株体内的脯氨酸含量产生显著影响,且离子浓度越高,影响越显著。但对不同草的影响是不同的,其中多年生黑麦草在脯氨酸含量上对Zn2+胁迫的耐受性要高于其他3种草。
2.3 Zn2+胁迫对4种草MDA含量的影响
在逆境条件下,植物为了减少自身伤害并维持正常生理功能,细胞膜中不饱和脂肪酸产生过氧化反应而产生丙二醛(MDA),因此可以用MDA衡量细胞膜损伤程度[13]。从表3可以看出,对4种草来说,Zn2+浓度为500 mg/kg处理下,其MDA含量除白三叶与草地早熟禾与对照差异不显著外,黑麦草与高羊茅表现出显著的差异性;Zn2+浓度为800~1 400 mg/kg 处理下,4种草的MDA含量都与对照差异显著,且均显著高于对照,并且随着重金属Zn2+浓度的升高,MDA含量逐渐上升趋势。说明低浓度Zn2+处理对4种草MDA含量的影响不大,只有当Zn2+浓度达到800 mg/kg及以上时,会显著影响草体内激素MDA的产生,造成上述4种草MDA含量显著高于对照。但对不同草而言,Zn2+胁迫对草MDA含量的影响又相对不一致。对多年生黑麦草与高羊茅而言,Zn2+胁迫浓度在500~1 400 mg/kg处理条件下,其体内MDA含量与对照差异显著,说明Zn2+浓度达到500~ 1 400 mg/kg 时,对其MDA含量的影响较大。对白三叶和草地早熟禾来说,Zn2+浓度为500 mg/kg时,对MDA含量的影响与对照差异不显著,说明Zn2+浓度在达到500 mg/kg时,对白三叶与草地早熟禾MDA含量的影响不大;而当Zn2+浓度达到800~1 400 mg/kg 时,对上述2种草MDA含量产生显著影响,且离子浓度越高,影响越显著。总之,Zn2+胁迫浓度的变化对不同草的影响是不同的,其中白三叶与草地早熟禾在MDA含量上对Zn2+胁迫的耐受性在低浓度时要高于其他2种草。
2.4 Zn2+胁迫对4种草SOD活性的影响
在逆境条件下,植物体内的超氧化物歧化酶(SOD)作为一种重要的防御酶,也能间接表明植物的受害程度和抗逆活性[14]。从表4可以看出,对4种草来说,Zn2+浓度为 500 mg/kg 处理下,除草地早熟禾的SOD活性与对照差异显著外,其余3种草均与对照差异不显著;当Zn2+浓度在800~1 400 mg/kg之间时,Zn2+对4种草胁迫皆与对照表现出差异显著,草体内的SOD活性均显著高于对照,并且4种草的SOD活性随着Zn2+浓度增加呈不断上升的趋势。说明,低浓度Zn2+处理对4种草的SOD活性影响不大,只有当Zn2+浓度达到800 mg/kg及以上时,会造成上述草SOD活性显著高于对照。但对于不同草坪草来说,Zn2+胁迫对草的SOD活性的影响又不完全一致。对多年生黑麦草、高羊茅与白三叶来说,Zn2+浓度在500 mg/kg处理时,其SOD活性与对照差异不显著,说明Zn2+浓度达到500 mg/kg时,对这3种草SOD活性的影响是一致的。对于草地早熟禾,Zn2+浓度为500~1 400 mg/kg 时,与对照相比SOD活性上升显著。总体上,Zn2+胁迫会对4种草体内的SOD活性较对照组产生显著影响,且离子浓度越高,影响越显著。但对不同草的影响是不同的,其中多年生黑麦草、高羊茅和白三叶对Zn2+胁迫的耐受性在SOD活性上相对高于草地早熟禾。
3 讨论与结论
麻占威等的研究表明,铅锌矿土法冶炼废渣中富含多种重金属,其中Zn含量最高,Pb、Cu、As含量次之,Cd、Cr含量相对较低[15]。王兰等的研究表明,铅锌矿淋溶液对植物幼苗生长有一定的抑制作用,且抑制作用随淋溶时间延长而减弱[16]。付天岭等的研究表明,水溶性有机质处理能够改善冶炼废渣中的肥力水平,同时能显著提高冶炼废渣上植物的成活率、株高、生物量及覆盖率;能促进冶炼废渣中多种有害重金属的雨水淋失和水浸溶出;同时能降低植物重金属含量,而提高单位面积上重金属植物富集量[17]。研究黔西北某锌冶炼厂的废渣堆场,通过调查分析废渣堆场土壤和优势植物的重金属含量,研究优势植物对重金属的吸收、转运和富集特征,筛选出适应该地区生态修复的先锋植物。调查的优势植物均能适应废渣堆场重金属元素较高的环境,对重金属具有一定的耐性[18]。通过模拟铅锌废渣污染特征及重金属释放机制对铅锌废渣进行改良,可为修复改善废渣堆场污染情况提供参考。根据废渣中典型重金属污染特点,筛选铅锌堆场修复的先锋物种,可进行铅锌堆场生态修复。
结果表明,Zn2+胁迫会对上述4种草植株生长产生显著影响,且离子浓度越高,影响越显著。但对不同草的影响是不同的:(1)Zn2+胁迫会对4种草植株体内的脯氨酸含量产生显著影响,且离子浓度越高,影响越显著,其中多年生黑麦草对Zn2+胁迫的耐受性要高于其他3种草;(2)白三叶与草地早熟禾在MDA含量上对Zn2+胁迫的耐受性在低浓度时要高于其他2种草;(3)Zn2+胁迫会对4种草植株体内的SOD活性产生显著影响,且离子浓度越高,影响越显著,其中多年生黑麦草、高羊茅和白三叶对Zn2+胁迫的耐受性要相对高于草坪草草地早熟禾。根据Zn2+对上述4种草生长及抗逆生理特性的影响研究结果筛选出先锋植物,通过模拟铅锌废渣中Zn2+的污染和現场采铅锌废渣进行改良种植的4种草的长势及生理指标的测定结果来看,可以选用多年生黑麦草、白三叶、高羊茅作为先锋草本植物进行铅锌冶炼废渣的原位生态修复。
参考文献:
[1]金文芬,方 晰,唐志娟. 3种园林植物对土壤重金属的吸收富集特征[J]. 中南林业科技大学学报,2009,29(3):21-25.
[2]王宏信. 重金属富集植物黑麦草对锌、镉的响应及其根际效应[D]. 重庆:西南大学,2006.
[3]陈同斌,韦朝阳,黄泽春,等. 砷超富集植物蜈蚣草及其对砷的富集特征[J]. 科学通报,2002,47(3):207-210.
[4]徐学华. 北方地区7种主要绿化树种对铅镉胁迫的生理生态响应[D]. 保定:河北农业大学,2010.
[5]吴新民,李恋卿,潘根兴. 南京市不同功能区土壤中重金属Cu、Zn、Pb和Cd污染特征[J]. 环境科学,2003,24(3):105-111.
[6]吴 攀,刘丛强,杨元根,等. 炼锌固体废渣中重金属(Pb、Zn)的 存在状态及环境影响[J]. 地球化学,2003,32(2):139-145.
[7]郑袁明,余 柯,吴泓涛,等. 北京市城市公园土壤铅含量及其污染评价[J]. 地理研究,2002,21(4):418-424.
[8]邹 琦. 植物生理学实验指导[M]. 北京:中国农业出版社,2000.
[9]李子芳,吴锡冬. 植物丙二醛含量测定试验设计方案[J]. 天津农业科学,2016,22(9):49-51.
[10]朱海生,刘建汀,陈敏氡,等. 丝瓜铜锌超氧化物歧化酶 Cu/Zn-SOD基因家族的克隆与表达分析[J]. 中国农业科学,2017,50(17):3391-3404.
[11]徐学华. 北方地区7种主要绿化树种对铅镉胁迫的生理生态响应[D]. 保定:河北农业大学,2010.
[12]燕平梅,章艮山. 水分胁迫下脯氨酸的累积及其可能的意义[J]. 太原师范专科学校学报,2000(4):27-28.
[13]袁祖丽. Cd、Pb污染对烤烟生理特性及生长发育的影响[D]. 郑州:河南农业大学,2006.
[14]Ruley A T,Sharma N C,Sahi S V. Antioxidant defense in a lead accumulating plant,Sesbania drummondii[J]. Plant Physiology and Biochemistry,2004,42(11):899-906.
[15]麻占威,吴永贵,付天岭,等. 不同粒径铅锌冶炼废渣中重金属分布特征[J]. 环境科学与技术,2015(4):149-154.
[16]王 兰,刘 方,王 俭,等. 铅锌矿渣酸性淋溶下重金属释放特征及其对植物幼苗生长的影响[J]. 生态学杂志,2010,29(6):1121-1126.
[17]付天岭,吴永贵,曾 艳,等. 水可溶性有机物在铅锌废渣生态修复中的应用[J]. 环境工程学报,2017,11(2):1178-1184.
[18]朱光旭,肖化云,郭庆军,等. 锌冶炼渣堆场优势植物的重金属累积特征研究[J]. 生态环境学报,2016,25(8):1395-1400.
