柱花草苯丙氨酸解氨酶(SgPALs)对生物胁迫与非生物胁迫的响应
2019-10-22郭鹏飞雷健罗佳佳刘攀道虞道耿罗丽娟
郭鹏飞 雷健 罗佳佳 刘攀道 虞道耿 罗丽娟


摘 要 本研究以熱研2号柱花草为材料,分析其苯丙氨酸解氨酶(PAL)活性、总酚含量、类黄酮含量、总抗氧化能力和SgPALs基因表达模式对生物胁迫(炭疽菌侵染)与非生物胁迫(干旱和盐胁迫)的响应。结果表明,在3种胁迫处理下,柱花草不同组织部位的PAL活性增加18.58%~123.56%,且叶片的总酚含量、类黄酮含量和总抗氧化能力分别增加65.11%~68.00%、51.00%~76.87%和83.00%~262.08%,差异显著。在干旱和盐胁迫处理下,根系的总酚含量、类黄酮含量和总抗氧化能力也显著提高43.77%~51.12%、45.46%~45.98%和60.45%~97.89%。随后对SgPALs的表达模式分析表明,除SgPAL4外,其他3个SgPALs受炭疽菌侵染诱导上调表达;在干旱胁迫下,根系中4个SgPALs均增强表达,但叶片中仅SgPAL1和SgPAL4上调表达;在盐胁迫下,根系中4个SgPALs也都上调表达,叶片中除SgPAL3下调表达外,其他3个SgPALs增强表达。综上所述,在遭受生物胁迫和非生物胁迫时,柱花草中的SgPALs基因表达及PAL酶活性升高,伴随着总酚与类黄酮含量的同步升高,表明其对环境胁迫的抗性升高。
关键词 柱花草;苯丙氨酸解氨酶;类黄酮;生物胁迫;非生物胁迫中图分类号 S59 文献标识码 A
Response of Phenylpropane Ammonia-lyase on Biotic and Abiotic Stress in Stylosanthes
GUO Pengfei1, LEI Jian1, LUO Jiajia1, 2, LIU Pandao2, YU Daogeng2, LUO Lijuan1*
1. Institute of Tropical Agriculture and Forestry, Hainan University, Haikou, Hainan 570228, China; 2. Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan 571101, China
Abstract Changes in PAL activity, total phenol and flavonoid content, total antioxidant capacity andSgPALsexpression pattern were analyzed to study the responses ofStylosanthesguianensiscv. Reyan2 to biotic (infectied byColletotrichum gloeosporioides) and abiotic (drought and salt) stresses. PAL activity increased by 18.58%-123.56% in different tissues under treatments of the three different stresses. The total phenol and total flavonoid content, and the total antioxidant capacity significantly increased by 65.11%-68.00%, 51.00%-76.87% and 83.00%-262.08%, respectively, in the leaf by pathogen infection, drought and salt stress, and similarly increased by 43.77%-51.12%, 45.46%-45.98% and 60.45%-97.89%, respectively, in the root by pathogen infection, drought and salt stress.PAL gene expression patterns showed that the clonedSgPALsgenes, exceptSgPAL4, were up-regulated by anthrax infection in the leaf. TheSgPALsgenes were up-regulated by drought stress inthe root while onlySgPAL1andSgPAL4were up-regulated in the leaf. Under salt stress, theSgPALsgenes were up-regulated in the root, while theSgPALsgenes, exceptSgPAL3, were down-regulated in the leaf.
Keywords Stylosanthes guianensis; phenylalanine ammonia-lyase; flavonoids; biotic stress; abiotic stress
DOI10.3969/j.issn.1000-2561.2019.09.011
全世界范围内,由于病原菌侵染、动物取食、干旱、盐害、低温等各种生物和非生物胁迫导致作物产量严重下降[1]。其中,病原菌侵染是较典型的生物胁迫之一,包括大豆疫霉病[2]、小麦纹枯病[3]、水稻稻瘟病[4]等,而盐害和干旱是限制农作物生产的典型非生物胁迫[5-6]。逆境环境下,植物物质代谢和能量代谢平衡遭受破坏,电子传递链缺少下游电子受体而被阻断,导致大量活性氧累积(reactive oxygen species, ROS),例如H2O2、O2?和OH?等,氧化损伤脂质、细胞膜、蛋白质、DNA等[7]。Mittler[8]报道,植物产生了10种抵抗ROS的策略,主要包括阻止ROS的再产生和清除已有的ROS,其中阻止ROS产生的机制有叶片卷曲、C4或景天酸代谢途径和抑制光合作用等,而清除机制包括酶系统的抗氧化酶(如CAT、POD、SOD等)和非酶系统的抗氧化次级代谢物(如多酚、类黄酮、类胡萝卜素等)。
苯丙烷代谢途径是植物重要的三大次生途径之一,其上游起始是以L-苯丙氨酸或酪氨酸为底物,在苯丙氨酸解氨酶(phenylalanine ammonia-lyase, PAL, EC 4.3.1.5)的催化下非氧化性脱氨,产生反式肉桂酸,最终生成香豆酰辅酶A,并以此为底物进入下游的类黄酮代谢、木质素代谢、香豆素代谢等分支[9]。近年来,基于基因组学技术的快速发展,大量研究从转录、蛋白以及代谢水平均阐明植物苯丙烷代谢及下游代谢物广泛参与各种逆境胁迫[10]。PAL是苯丙烷代谢的关键限速酶,其编码基因属于多基因家族成员,不同物种其拷贝数各异,从几个成员(拟南芥含4个)至几十个(西红柿含有26个)[11]。研究发现PAL转录水平受环境刺激的调控,且各同源基因具有不同分工,例如拟南芥中,缺氮能特异诱导4个同源基因的AtPAL1和AtPAL2表达,而双突变体拟南芥pal1/pal2不能合成类黄酮,证明AtPAL1和AtPAL2主要介导下游类黄酮代谢[12]。Tonnessen等[13]利用数量性状基因座(quantitative trait loci, QTL)定位到OsPAL与水稻抗病性相关,遗传证据表明OsPAL4参与水稻对细菌性枯萎病、稻瘟病和叶鞘枯萎病的防御反应。此外,类黄酮本质为多酚类物质,具有一个或多个酚羟基基团,显示较强的还原性,能消除环境胁迫产生的ROS。Feucht等[14-15]报道,高温和干旱能改变细胞核中类黄酮的含量,而核定位的类黄酮主要作用是保護处于有丝分裂阶段无核膜包围的染色体免受氧化伤害。
柱花草(Stylosanthesspp.)是重要的多年生豆科牧草,具有耐贫瘠和耐酸性土壤等特点[16],但生物胁迫(炭疽病)和非生物胁迫(干旱、盐害)严重限制了柱花草的种植和生产[17-19]。目前,对柱花草响应干旱、盐害、炭疽病胁迫在形态、生理、生化等方面已有文献报道,且抗氧化机制主要集中在酶系统方面[17-19],但其抵抗逆境胁迫的非酶系统生理及分子机制鲜有报道。本课题组前期研究发现,接种炭疽菌的柱花草叶片其胁迫响应基因SgPAL1显著上调,但其酶活及介导的下游类黄酮代谢等支路如何变化尚不清楚[20]。因此,本研究通过分析干旱、盐害和接种炭疽菌胁迫下,柱花草PAL酶活性、总酚含量、类黄酮含量和总抗氧化能力的变化以及SgPALs的表达模式,以期解析以下2个问题:(1)SgPAL介导的苯丙烷代谢及下游酚类代谢对生物胁迫和非生物胁迫的响应机制如何,是否一致?(2)逆境胁迫下,柱花草SgPAL基因家族成员的表达模式如何?本研究为柱花草抗逆品种的选育和基因改良提供理论基础。
1 材料与方法
1.1材料
供试柱花草种质为热研2号(Stylosanthes guianensiscv. Reyan2),种子来自中国热带农业科学院热带作物品种资源研究所牧草业研究室。
1.2方法
1.2.1 试验设计 本试验在海南大学热带农林学院科研基地的大棚进行。热研2号种子经80 ℃热水浸泡2 min,冷却至室温后,育苗盒播种,以腐殖土∶蛭石按1∶1混匀为基质,幼苗正常培养4~5周。分别进行接种炭疽菌、干旱、盐害处理。每种处理均设置3个生物学重复。
接种炭疽菌处理。接种所用菌株为实验室课题组前期从海南岛采集,分离并纯化的胶孢炭疽菌菌株[21]。参考Wang等[20]方法,制备接种的炭疽菌孢子悬浮液,即配置PDA培养基,划板接种胶孢炭疽菌,置于28 ℃暗培养约10 d,无菌水洗脱孢子。使用血球计数板,控制孢子浓度在106/mL,并加入0.2% Tween20。同样,实验设置2个处理,用100 mL孢子悬浮液喷洒热研2号柱花草直到叶片上凝成水珠,对照用含有0.2% Tween20的100 mL无菌水处理。接种60 h后,收获材料。
干旱处理。选取长势基本一致正常的柱花草幼苗,浇灌500 mL的含10% PEG-6000霍格兰氏营养液,模拟干旱条件,对照用500 mL霍格兰氏营养液浇灌。处理7 d后收获样品。
盐害处理。取土培的柱花草幼苗,浇灌500 mL的含0.5% NaCl霍格兰氏营养液,对照用500 mL霍格兰氏营养液浇灌。处理3 d后收获样品。
1.2.2 PAL酶活性测定 采用南京建成生物工程研究所的试剂盒(中国,货号为A137),PAL可以催化L-苯丙氨酸裂解为反式肉桂酸,反式肉桂酸在290 nm处有最大吸收值,通过测定吸光值变化值计算PAL活性。具体操作步骤参考该试剂盒的说明书。
1.2.3 总抗氧化能力(T-AOC)测定 采用南京建成生物工程研究所的试剂盒(中国,货号为A015),植物体内的抗氧化物质,可以将Fe3+还原成Fe2+,Fe2+可与菲啉类物质形成络合物,通过比色测出抗氧化能力。具体操作步骤参考试剂盒的说明书,单位按每毫克蛋白中的酶活力单位数计算(U/mg)。
1.2.4 总酚含量测定 参考Chang等[22]方法,采用Folin-Ciocalteu法进行比色分析,即通过磷酸缓冲液提取总酚后,加入Folin-Ciocalteu显色剂,在765 nm波长处测定吸光度,同时以没食子酸绘制标准曲线,定量总酚含量。
1.2.5 类黄酮含量测定 采用Uarrota等[23]方法,略有改动。采用磷酸缓冲液提取类黄酮后,配置2 mL的反应体系:100 μL提取液,0.5 mL 95%乙醇、0.1 mL 1 mol/L乙酸鉀、0.1 mL 10% AlCl3溶液,以及1.2 mL蒸馏水。混匀后,反应30 min,检测415 nm波长处的吸光度,并以槲皮素绘制标准曲线,进行定量。
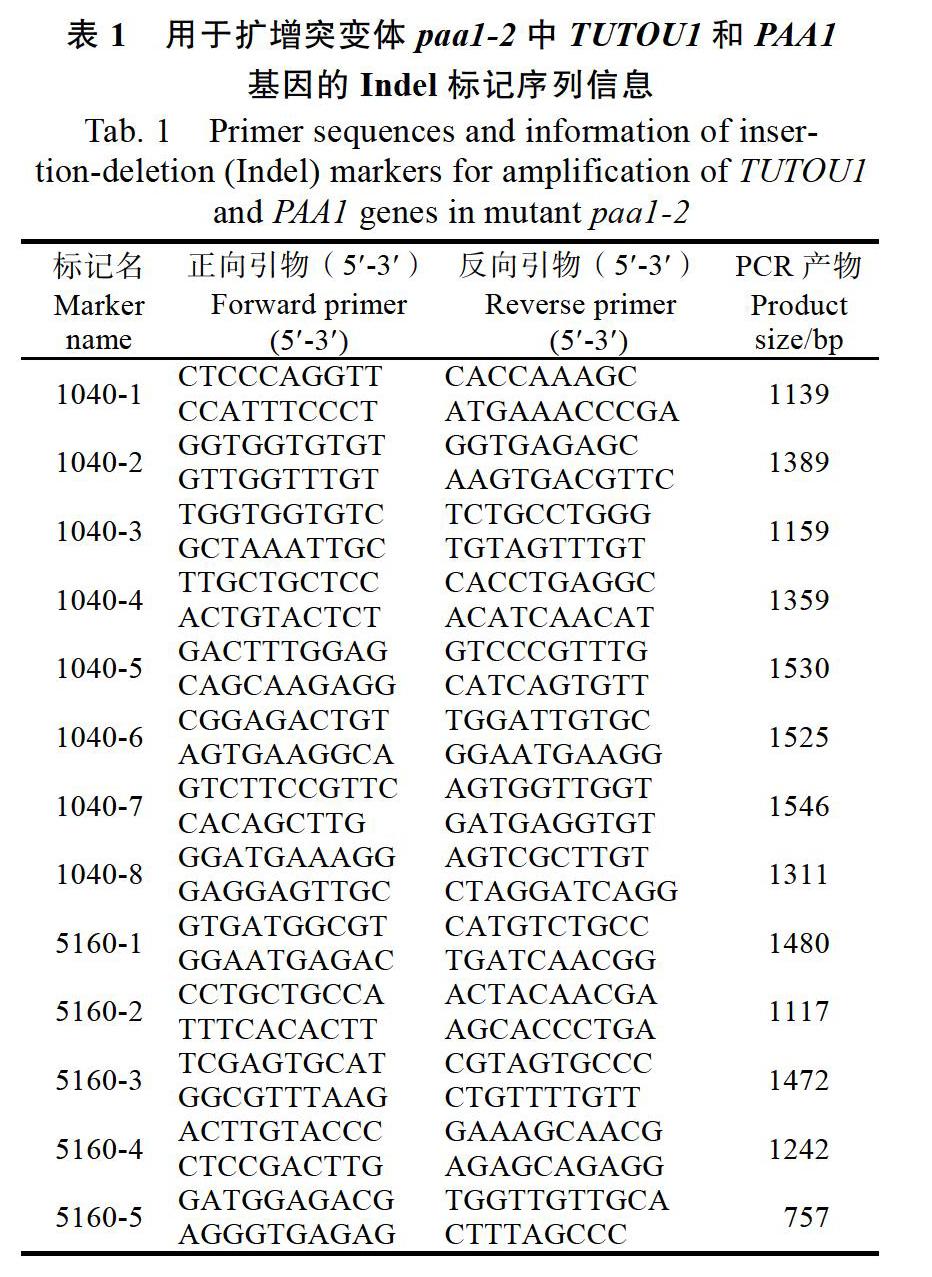
1.2.6SgPALs的克隆和Real-time PRC分析 参照TRIzol(Invitrogen Inc,美国)方法提取柱花草总RNA,并用TaKaRa公司的PrimeScript? RT reagent Kit with gDNA Eraser试剂盒合成cDNA第一链,?20 ℃保存备用。根据柱花草转录组测序结果[24],利用Oligo 7软件设计SgPAL基因全长引物和定量引物(表1),以cDNA为模板,采用高保真聚合酶Phusion(Thermo)扩增SgPAL的全长片段。PCR扩增产物经琼脂糖凝胶电泳检测,并进行切胶回收纯化的目的片段,连接到pEASY-T1(TaKaRa,日本)载体上,转化大肠杆菌DH5α感受态细胞,测序分析后获得4个SgPAL基因全长cDNA序列。此外,参考Liu等[25]方法,以柱花草看家基因SgEF-1α为内参,运用SYBR? Premix Ex Taq? II(TaKaRa,日本)定量试剂盒进行Real-time PCR分析,用Rotor-Gene Q(Qiagen, Hilden,德国)进行反应。20 μL的反应体系为:10 μL 2×SYBR Green PCR master mix,6.4 μL Mili-Q无菌水,0.8 μL 10 μmol/L引物,2 μL浓度为200 ng/μL的cDNA模板。反应程序为:95 ℃ 1 min,94 ℃ 15 s,60 ℃ 15 s,72 ℃ 30 s,循环40次。相对表达量=目的基因表达量/看家基因(SgEF-1α)表达量,定量引物如表1所示。
1.2.7SgPALs的生物信息学分析 运用DNAMAN进行多序列比对分析,采用MEGA(v5.50)进行系统进化树分析,利用NCBI网站进行保守结构域分析。
1.3数据处理
采用Microsoft Office 2016软件对本研究相关数据进行分析并绘制图表,以平均值和标准误表示检测结果,并使用IBM SPSS Statistics 20(SPSS Institute,美国)软件进行统计分析,对同一组织不同处理的样品运用独立样本T检验分析差异的显著性。
2 结果与分析
2.1相关生理指标测定
2.1.1 炭疽菌侵染对柱花草PAL活性、总酚含量、类黄酮含量和总抗氧化能力的影响 由图1可知,接种炭疽菌后,柱花草叶的PAL活性、总酚含量、类黄酮含量和总抗氧化能力与对照相比均极显著提高(P<0.01),分别提高了41%、68%、51%和83%。在柱花草茎中,炭疽菌侵染后仅PAL活性和总抗氧化能力分别极显著提高48%和51%(P<0.01),但总酚含量和类黄酮含量无显著变化。以上结果表明,柱花草的叶片和茎通过增强抗氧化能力抵抗炭疽菌的侵染,叶片PAL参与调控总酚和类黄酮代谢对炭疽病的响应。
2.1.2 干旱胁迫对柱花草PAL活性、总酚含量、类黄酮含量和总抗氧化能力的影响 如图2所示,与对照相比,干旱胁迫下(PEG处理)柱花草叶、茎和根中的PAL活性、总酚含量、类黄酮含量和总抗氧化能力均显著提高(P<0.05)。其中,在PEG处理下,叶、茎和根的PAL活性分别提高了46.12%、85.21%和123.56%;总酚含量分别提高了65.11%、123.33%和51.12%;类黄酮含量分别提高了51.83%、133.68%和45.46%;总抗氧化能力分别提高了119.15%、31.25%和60.45%。以上结果显示,柱花草PAL调控下游的总酚和类黄酮代谢,通过提高抗氧化能力来适应干旱胁迫。
2.1.3 盐胁迫对对柱花草PAL活性、总酚含量、类黄酮含量和总抗氧化能力的影响 如图3所
示,与对照相比,当遭受盐胁迫时(NaCl处理),柱花草的PAL活性和类黄酮含量均显著提高(P<0.05)。其中,叶、茎和根的PAL活性分别提高了45.44%、18.58%和45.51%,类黄酮含量分别提高了76.87%、18.51%和45.98%。NaCl处理下,总酚含量和总抗氧化能力仅在叶和根中显著提高,而对茎无影响。以上结果表明,PAL介导的代谢途径参与了柱花草对盐胁迫的响应。
2.2SgPALs序列的克隆及生物信息学分析
根据柱花草4个PAL基因序列,并设计了全长引物(表1),以cDNA为模板,扩增出4个SgPAL基因的开放阅读框(open reading frame, ORF)序列,经测序分析发现所有基因全长序列与转录组结果完全一致,SgPAL1、SgPAL2、SgPAL3、SgPAL4這4个基因的大小依次为2145、2073、2133、2199 bp(图4),分别编码715、691、711、733个氨基酸。
基于在线NABI数据库的Conserved Domains分析(https://www.ncbi.nlm.gov/cdd/)发现,4个SgPALs均属于苯丙氨酸解氨酶家族(PLN02457)成员,均具有2个保守的脱氨作用位点,即苯丙氨酸/组氨酸氨裂解酶蛋白标签。多序列比对分析表明,4个SgPAL包含PAL家族蛋白的保守活性位点:G-[STG]-[LIVM]-[STG]-[AC]-S-G- [DH]-L- x-P-L-[SA]-x(2)-[SAV],其中内部三肽A-S-G是自催化环化和脱水形成4-甲基-5-咪唑基团(4-methyl diene-imidazol-5-one, MIO)的关键活性位点,而MIO可直接消除α-氨基。此外,将4个SgPALs基因的假定氨基酸序列与模式植物拟南芥、水稻以及验证功能的大豆PALs构建系统进化树,发现PALs蛋白可以分为2个类群即单子叶植物(水稻)和双子叶植物(拟南芥、大豆、柱花草)(图5),柱花草SgPAL1、SgPAL3、SgPAL4属同小分枝,且SgPAL1与SgPAL4两者序列相似性较高为87.18%,其中SgPAL1与大豆GmPAL-2.1(NP_001236956.2)相似性最高为87.59%,SgPAL3、SgPAL4与大豆GmPAL(Glyma03g 33880)相似性最高分别为84.66%、88.95%。同样,SgPAL2与大豆GmPAL(Glyma03 g33880)氨基酸序列相似性最高为69.92%。
2.3 SgPALs基因的表达模式分析
2.3.1 炭疽菌对SgPALs表达的影响 如图6所示,接种炭疽菌后,柱花草叶和茎中的SgPAL1、SgPAL2和SgPAL3的表达量与对照相比提高45.65%~1061.25%,差异显著,但SgPAL4在叶中的表达显著下调72.37%(图6D)。
2.3.2 干旱对SgPALs表达的影响 由图7可知,与对照相比,干旱胁迫显著上调了4个SgPALs在根系中的表达,增加了88.11%~1024.63%;在茎中,仅SgPAL4受干旱胁迫诱导上调表达;在叶片中,SgPAL1和SgPAL4受干旱胁迫诱导上调表达。
2.3.3 盐胁迫对SgPALs表达的影响 由图8可知,在盐胁迫下,柱花草4个SgPALs基因在根系中均显著上调表达,增加了103%~479%;柱花草的SgPAL1、SgPAL2和SgPAL3在茎中受盐胁迫诱导显著上调表达,其中上调幅度最大的基因是SgPAL3,达3.63倍;在叶片中,SgPAL1、SgPAL2和SgPAL4受盐胁迫诱导显著上调表达。
3 讨论
炭疽病是由炭疽菌侵染植物引起的真菌病害,严重抑制植物生长,包括山葵、辣椒、柱花草等[20,26-28]。干旱和盐害是典型的非生物胁迫逆境因素,在胁迫下植物体内活性氧过量累积,导致膜脂过氧化、细胞膜透性增强、细胞组分改变,严重制约植物的生长和产量[5,17-18]。柱花草是重要的热带豆科牧草,起源于中南美洲及加勒比海地区,具有适应性强、耐酸瘦土壤、耐旱等优点,是研究植物抗生物胁迫(炭疽病)和非生物胁迫(干旱和盐害)的理想材料[17-18,20]。
黄酮类物质是苯丙烷代谢途径的一大类下游产物,在植物体内具有多种功能,可作为生长发育的调节因子和信号分子,能清除紫外线,具有抗菌和抗氧化的功效等,广泛响应环境对植物的刺激,其中研究较为透彻的是其作为抗氧化剂,吸收自由基、淬灭单线态氧、分解过氧化物,调节活性氧和自由基在植物体内的稳态平衡,降低植物的氧化损伤[29]。PAL是连接初级代谢(莽草酸途径)和次级代谢(苯丙烷途径)的关键诱导酶,其活性和浓度控制着下游代谢产物的产生效率,进而影响植物的抗逆策略,研究发现大量逆境因素(如高温、盐害、紫外线等)均能提高PAL的酶活性,转录水平也发现大量PAL的同源基因显著上调表达[30]。本研究克隆了4个SgPALs基因(图4),多序列比对分析发现它们均属于苯丙氨酸解氨酶家族(PLN02457)成员。
研究发现植物在抵抗病原菌侵染时,主要由局部防御和系统防御2种措施,其中系统防御措施主要依靠在远离感染部位的组织中大量合成抗菌复合物并转运到受伤部位抑制微生物,包括多酚、木质素、异黄酮-植保素等[2]。同时,类黄酮已被证明具有抗菌活性,能参与保护植物抵抗病原菌的侵染[30]。如在芒果中,含花青素和类黄酮较高的红色芒果具有更强的抗炭疽菌能力[3]。Zhang等[31]研究表明,西红柿突变体Delia体内花青素累积,导致ROS减少,降低灰霉病的易感性。本研究也得到一致的结果,柱花草接种炭疽菌后,与对照相比,其叶片的PAL酶活性、总酚含量、类黄酮含量及总抗氧化能力均显著提高,而茎的PAL酶活性和总抗氧化能力显著增强,结果表明炭疽菌侵染柱花草叶片诱导总酚和类黄酮等苯丙烷代谢途径的下游代谢物大量产生,以防御炭疽菌的侵害,而柱花草茎可能通过上调苯丙烷代谢途径的其他代谢物提高抗氧化能力,以抗炭疽病。此外,本研究发现柱花草叶和茎均有3个基因显著上调表达,其中SgPAL3在叶和茎中的上调幅度均最高(图6)。进化树分析发现SgPAL3与大豆GmPAL(Glyma03g33880)相似性最高,且Shine等[32]研究发现GmPAL(Glyma03g33880)能调控水杨酸的累积,抵抗大豆疫霉病,因此推测SgPAL3可能具有影响柱花草抗炭疽病的功能。
正常条件下,植物体内ROS的生成和清除处动态平衡状态,当植物遭遇干旱和盐害等非生物胁迫时,稳态被破坏,导致ROS迅速累积,氧化损伤植物[8]。而类黄酮代谢物具有较强的抗氧化活性,能清除ROS,保护植物。研究发现,干旱胁迫能诱导黄酮类累积,提高抗氧化能力,包括小茴香、拟南芥、葡萄等[33]。同时,在植物的叶、根中,盐胁迫能诱导多酚、类黄酮等抗氧化代谢物的累积,消除ROS,包括丹参、玉米、龙葵等[34]。以上研究表明苯丙烷代谢途径的类黄酮等酚类化合物对植物抗旱和抗盐害起重要作用。同样,本研究也发现,干旱胁迫下的柱花草叶、茎、根和盐胁迫下柱花草的叶、根的PAL酶活性、总酚含量、类黄酮含量以及总抗氧化能力均显著升高(图2,图3),但盐胁迫下柱花草茎的总酚和总抗氧化能力无显著变化(图3)。RT-qPCR分析表明,干旱和盐害胁迫下均有SgPALs基因不同程度地显著上调,其中干旱胁迫下,SgAPL4在叶和茎中的上调幅度最大(图7D),SgAPL2在根中的上調表达倍数达10.24(图7B)。盐胁迫下,柱花草叶、茎、根中显著上调幅度最大的基因分别是SgPAL3、SgPAL3、SgPAL4。同时,进化分析表明SgPAL3、SgPAL4均与GmPAL(Glyma03g33880)相似性最高,而实验也表明GmPAL(Glyma03g33880)能调控下游水杨酸代谢[32],因此推测SgPAL3、SgPAL4可能调控下游代谢物的合成,抵抗非生物胁迫。
綜上所述,本研究发现所检测的不同组织和胁迫下,柱花草PAL酶活均显著升高,除盐胁迫和接种炭疽菌下柱花草茎以外,其余总抗氧化能力及贡献抗氧化作用的总酚、类黄酮也均提高。表达模式分析结果显示,生物胁迫(炭疽病)和非生物胁迫(干旱和盐害)下,柱花草均有SgPALs基因不同程度地显著上调。结果表明,接种炭疽菌(叶)、干旱(叶、茎、根)和盐害(叶、根)下的大部分柱花草组织以提高SgPAL的表达和PAL的活性,促进下游总酚和类黄酮的代谢,以提高总抗氧化能力响应生物和非生物胁迫。
参考文献
[1] Sewelam N, Kazan K, Schenk P M. Global plant stress signaling: reactive oxygen species at the cross-road[J]. Frontiers in Plant Science, 2016, 7(187): 187.
[2] Zhang C, Wang X, Zhang F,et al. Phenylalanine ammonia-lyase2.1 contributes to the soybean response towardsPhytophthora sojaeinfection[J]. Scientific Reports, 2017, 7(1): 7242.
[3] Sivankalyani V, Feygenberg O, Diskin S, et al. Increased anthocyanin and flavonoids in mango fruit peel are associated with cold and pathogen resistance[J]. Postharvest Biology and Technology, 2016, 111: 132-139.
[4] Gong Q, Yang Z, Chen E,et al. A phi-class glutathione S-transferase gene for Verticillium wilt resistance inGossypium arboreumidentified in a genome-wide association study[J]. Plant and Cell Physiology, 2017, 59(2): 275-289.
[5] Supratim B, Venkategowda R, Anuj K,et al. Plant adaptation to drought stress[J]. F1000 Research, 2016, 5: 1554.
[6] Marcum K B. Salinity tolerance mechanisms of grasses in the subfamily chloridoideae[J]. Crop Science, 1999, 39(4): 1153-1160.
[7] Chen Y, Lin F, Yang H,et al. Effect of varying NaCl doses on flavonoid production in suspension cells ofGinkgo biloba: relationship to chlorophyll fluorescence, ion homeostasis, antioxidant system and ultrastructure[J]. Acta Physiologiae Plantarum, 2014, 36(12): 3173-3187.
[8] Mittler R. Oxidative stress, antioxidants and stress tolerance[J]. Trends in Plant Science, 2002, 7(9): 405-410.
[9] Fraser C M, Chapple C. The phenylpropanoid pathway in Arabidopsis[J]. The Arabidopsis Book, 2011, 9: e0152.
- Treutter D. Significance of flavonoids in plant resistance: a review[J]. Environmental Chemistry Letters, 2006, 4(3): 147-157.
- Chang A, Lim M H, Lee S W, et al. Tomato phenylalanine ammonia-lyase gene family, highly redundant but strongly underutilized[J]. Journal of Biological Chemistry, 2008, 283(48): 33591-33601.
[12] Rohde, A. Molecular phenotyping of the pal1 and pal2 mutants ofArabidopsis thalianareveals far-reaching consequences on phenylpropanoid, amino acid, and carbohydrate metabolism[J]. The Plant Cell Online, 2004, 16(10): 2749-2771.
[13] Tonnessen B W, Manosalva P, Lang J M,et al. Rice phenylalanine ammonia-lyase geneOsPAL4 is associated with broad spectrum disease resistance[J]. Plant Molecular Biology, 2015, 87(3): 273-286.
[14] Feucht W, Treutter D, Dithmar H,et al. Loss of nuclear flavanols during drought periods inTaxus baccata[J]. Plant Biology, 2013, 15(3): 462-470.
[15] Feucht W, Treutter D, Jürgen P. Flavanols in nuclei of tree species: facts and possible functions[J]. Trees: Structure and Function, 2012, 26(5): 1413-1425.
[16] 陳志坚, 罗佳佳, 彭玺如, 等. 不同柱花草种质的抗旱性分析[J]. 热带作物学报, 2017, 38(10): 1790-1795.
[17] 薛 瑞, 周广奇, 胡新文, 等. 柱花草种质抗旱性综合评价[J]. 中国农学通报, 2009, 25(11): 224-233.
[18] 刘一明, 冯 宇, 丁西朋, 等. 67份柱花草耐盐性综合评价[J]. 热带作物学报, 2017, 38(11): 2040-2049.
[19] 易克贤. 柱花草炭疽病及其抗病育种进展[J]. 中国草地学报, 2001, 23(4): 59-65.
[20] Wang H, Chen Z, Liu G, et al. Alterations of growth, antioxidant system and gene expression inStylosanthes guianensisduringColletotrichum gloeosporioidesinfection [J]. Plant Physiology & Biochemistry, 2017, 118: 256.
[21] Jia Y X, Chen Z J, Yang M X,et al. First report of anthracnose onStylosanthes guianensiscaused byColletotrichum carstiiin China[J]. Plant Disease, 2017, 101(4): 630.
[22] Chang C C, Yang M H, Wen H M,et al. Estimation of total flavonoid content in propolis by two complementary colorimetric methods[J]. Journal of Food and Drug Analysis, 2002, 10(3): 178-182.
[23] Uarrota V G, Moresco R, Coelho B,et al. Metabolomics combined with chemometric tools (PCA, HCA, PLS-DA and SVM) for screening cassava (Manihot esculentaCrantz) roots during postharvest physiological deterioration[J]. Food Chemistry, 2014, 161: 67-78.
[24] Jiang C, Liu L, Li X,et al. Insights into aluminum-tolerance pathways inStylosanthesas revealed by RNA-Seq analysis[J]. Scientific Reports, 2018, 8(1): 6072.
[25] Liu P D, Xue Y B, Chen Z J,et al. Characterization of purple acid phosphatases involved in extracellular dNTP utilization inStylosanthes[J]. Journal of Experimental Botany, 2016, 67(14): 4141-4154.
[26] Zhang X, Liu C J. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids[J]. Molecular Plant, 2015, 8(1): 17-27.
- Macdonald J L, Punja Z K. Occurrence of botrytis leaf blight, anthracnose leaf spot, and white blister rust on Wasabia japonica in British Columbia[J]. Canadian Journal of Plant Pathology, 2017, 39(1): 60-71.
- Diao Y Z, Zhang C, Liu F, et al. Colletotrichum species causing anthracnose disease of chili in China[J]. Persoonia Molecular Phylogeny & Evolution of Fungi, 2017, 38: 20-37.
- Vera Saltos M B, Naranjo Puente B F, Milella L, et al. Antioxidant and free radical scavenging activity of phenolics from Bidens humilis[J]. Planta Medica, 2015, 81(12/13): 1056-1064.
[30] Treutter D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis[J]. Plant Biology, 2010, 7(6): 581-591.
- Zhang Y, Butelli E, De Stefano R, et al. Anthocyanins double the shelf life of tomatoes by delaying overripening and reducing susceptibility to gray mold[J]. Current Biology, 2013, 23(12): 1094-1100.
- Shine M B, Yang J W, El-Habbak M, et al. Cooperative functioning between phenylalanine ammonia lyase and isochorismate synthase activities contributes to salicylic acid biosynthesis in soybean[J]. New Phytologist, 2016, 212: 627-636.
- Alinian S, Razmjoo J, Zeinali H. Flavonoids, anthocynins, phenolics and essential oil produced in cumin (Cuminum cyminum L.) accessions under different irrigation regimes[J]. Industrial Crops and Products, 2016, 81: 49-55.
- Hamada A E, Gaurav Z, Hegab M M, et al. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs[J]. Frontiers in Plant Science, 2016, 7(580): 276.
