Effects of Dietary Fat Levels on Growth, Nutrient Digestibility,Nitrogen Utilization and Fur Quality of Growing-furring Blue Foxes
2019-10-22CaoShuxinLiuDaweiWangChaoWeiJingkunSunJiayouBaiYangandXuLiangmei
Cao Shu-xin, Liu Da-wei, Wang Chao, Wei Jing-kun, Sun Jia-you, Bai Yang, and Xu Liang-mei
Institute of Animal Nutrition, Northeast Agricultural University, Harbin 150030, China
Abstract: This study evaluated the effects of dietary fat levels on the growth, nutrient digestibility, nitrogen balance and pelt characteristics of the blue fox (Alopex lagopus). Age-matched blue foxes (12 weeks old) with similar body weights were randomly divided into six dietary groups over 98 days, during the growing-furring period. Blue foxes in the six groups were fed with experimental diets containing 10% (groupⅠ), 12% (groupⅡ), 14% (group Ⅲ), 16% (group Ⅳ), 18% (groupⅤ) and 20% (group Ⅵ)fat during the growing period (from 13 weeks to 18 weeks of age); foxes were fed with diets containing 12% (groupⅠ), 14%(groupⅡ), 16% (group Ⅲ), 18% (group Ⅳ), 20% (groupⅤ) and 22% (group Ⅵ) fat during the furring period (from 18 weeks to 26 weeks of age). The results indicated that 16%-18% dietary fat, during the growing period significantly improved nutrient utilization,and as a main energy source, significantly reduced the feed/gain ratio. A dietary fat level of 18%, during the furring period not only improved the growth and development and decreased the feed/gain ratio, but also increased nutrient digestibility and fur quality. These data, combined with data from a conic regression model, showed that dietary fat levels between 16.29% and 18.40%, during the growing period and between 17.72% and 18.72%, during the furring period enhanced the growth performance and fur quality of blue foxes.
Key words: blue foxes, fat, digestibility, nitrogen balance, fur quality
Introduction
The blue fox (Alopex lagopus) is also called the arctic fox and it belongs to the Canidae family of carnivorous mammals, which is also known as the dog branch of the carnivore family (Hersteinsson 1989;Burlikowskaet al., 2015). The blue fox is mostly found on farms and is the descendant of arctic foxes from Greenland, Alaska, Iceland and Svalbard that were trapped approximately 80 years ago (Kempe and Strandén, 2016; Laiet al., 2017; Hasselgrenet al.,2018). Blue foxes mainly eat animal feedstuff that is high in protein and fat because their digestive systems are shorter than the intestine to body length ratio of 4 : 1 and their average food passage time is 6-8 h(Risinget al., 2017). Dietary fat content and fat use is high in fur-bearing animals. Up to 44% of adipose tissue in fresh general feed has been fed to foxes without any detrimental effects (Genget al., 2011 ).Kopczewskiet al. (2001) showed that the increase in energy content in feed causes a significant increase in the body weight of foxes and improvement in their hair quality. There is no difference in the anatomical and histopathological manifestations of each group.Nevertheless, high-energy feeding causes vacuolar and fatty degeneration of liver and kidneys. Blue foxes tolerate fat well; however, the excessive dietary fat content impedes growth of blue fox. Genget al. (2012)stated that 40% dietary fat significantly increases the index of fat deposition, especially the hepatic fat rate and liver to body ratio; moreover, hyperlipidemia and hyperglycosemia are detected and the liver is badly damaged.
Fat supplementation improves cost effectiveness of feeding by increasing the energy concentration of the feed and thus decreasing feed consumption. In addition, as the major energy source for blue foxes,fat acts as a physiological carrier for feed flavor and fat-soluble vitamins. The appropriate increase in dietary fat may decrease N excretion, which increases the biological value of dietary protein in each developmental phase which, in turn, improves fur quality (Prestrud and Nilssen, 1992; Liuet al., 2008;Zhanget al., 2017). Fat is a major energy source and a nutrient that may be stored in adipose tissue of fox during the winter months when feed resources may be limited. Furthermore, fat provides thermal insulation and physical protection for internal organs, is the only source of essential fatty acids for prostaglandin synthesis of prostaglandins, and plays an important role in metabolic regulation (Koskinenet al. 2014).Dietary fat may improve fur quality in blue foxes.Genget al. (2012) stated that the most preferable fur quality and efficiency of ME used for gain are obtained when the diet contained appropriate fat levels in the growing-furring period.
This study focused on the effects of dietary fat levels on nutrient absorption, metabolism and fur quality in blue foxes. Hopefully, these data would provide a reliable basis for the development, selection and use of optimal dietary fat levels for the growingfurring period to improve fur quality in blue foxes.
Materials and Methods
Animals and experimental diets
A total of 60 larva blue foxes (12 weeks of age), with healthy, similar age and body weight, were randomly allocated into six groups; each blue fox was considered an experimental replicate. Littermates of the same sex were assigned to different groups. The experimental period (the growing-furring period) was divided into two phases, the growing period (from 13 weeks to 18 weeks of age) and the furring period (from 18 weeks to 26 weeks of age). Blue foxes in the six groups were fed with experimental diets containing 10% (groupⅠ),12% (groupⅡ), 14% (groupⅢ), 16% (group Ⅳ),18% (groupⅤ) and 20% (groupⅥ) fat levels of the growing period; the furring period were fed with diets containing 12% (groupⅠ), 14% (group Ⅱ), 16%(group Ⅲ), 18% (group Ⅳ), 20% (group Ⅴ) and 22% (groupⅥ) fat. All the diets met the nutrient requirements of growing-furring foxes (NRC, 1982)and other nutrients were the same except for fat level (Tables 1 and 2). The experiment included an adaptation period of one week (the 13th week) for animals to accustom the experimental diets based on dry and powdery components mixed with water, and the test lasted for 14 weeks.
Feeding management
During the experiment, blue foxes were individually housed outdoors in conventional cages (1.00 m long×0.70 m wide×0.80 m high) with natural light. The experimental diets were mixed with water (1 : 3) prior to consumption, foxes were fed waterad libitum. Each fox was given the same amount of daily feed (weighed to an accuracy of 0.001 kg) and fed twice a day at 7: 00 a.m. and 5: 00 p.m.
Growth performance
All the blue foxes fasting weights were recorded at 12,18 and 26 weeks of age. The amounts of food given to foxes and amounts leftover were recorded daily,during the experimental period. The average daily gain(ADG) and statistical feed intake by replicate units i.e., the calculated average feed intake (ADFI) for each treatment group, were calculated according to the ADG and the ADFI calculated feed conversion rate formulas.
Daily matter intake (kg)=Mass of food given-mass of food leftover recorded
Average daily gain (kg)=(Final weight-initial weight)/trial days
Feed conversion rate=(Final weight-initial weight)/Average weight gain
Daily ME intake (MJ · d-1)=Daily metabolism×feed consumption
Metabolizable energy/Gain (MJ · kg-1)=Daily ME intake/Daily gain
Digestion metabolism experiment
Six blue foxes were randomly selected from each group and used for digestibility measurements.Their urine and feces were collected separately.The digestibility measurements were conducted for 3 days at 18 and 26 weeks old, respectively. The feces and urine were collected on a 24 h cycle at 7: 00 a.m. During the period, excretions of animals were collected every day. The collected feces were preserved with 10% hydrochloric acid that accounted for 10% of the weight of fresh drains and stored at -20℃ until analysis. Urine was collected at the completion of each period and preserved with 20%sulfuric acid that accounted for 2% of the volume of fresh drains and stored at -20℃ until analysis.
Dry matter was quantified by drying feed or fecal samples at 120℃ for 15 minutes, then at 65℃ for 48 h to constant weight. The contents of crude protein(CP), ether extract (EE), crude ash, calcium and phosphorus in the feed, feces and urine were analyzed using the standard methods of the AOAC (2005). The gross energy (GE) of the fecal samples and feed was measured using an oxygen bomb calorimeter (Parr 6 300, USA Parr Instrument Company).

Table 1 Composition and nutrient levels of experimental diets during growing period (air-dry basis)

Table 2 Composition and nutrient levels of experimental diets during furring period (air-dry basis)
The apparent digestibility (AD) coefficient of the nutrients and energy was calculated as the following:AD(%)=(a-b)/a×100, where 'a' was the nutrient or energy intake from the feed, and 'b' was the nutrient or energy excretion in the feces.
Feed conversion rate (%)=[(Final weight-initial weight)/Feed intake]×100
Dry matter digestibility (%)=[(Dry matter intakedry matter output)/Dry matter intake]×100
Protein digestibility (%)=[(Protein intake-protein in feces)/Protein intake]×100
Fat digestibility (%)=[(Fat intake-fat in feces)/Fat intake]×100
Ca digestibility (%)=[(Ca intake-Ca in feces)/Ca intake]×100
P digestibility (%)=[(P intake-P in feces)/P intake]×100
N deposition (g · d-1)=N intake-urinary N-fecal N
Net protein utilization=(N deposition/N intake)×100
Biological value (%)=[N deposition/(N intake-fecal N)]×100
Fur quality
At the end of the experiment (26 weeks), six blue foxes were randomly selected from each group, killed by electrocution and pelted immediately by skilled workers according to the Welfare of Animals Kept for Fur Production. The final body lengths of blue foxes, accurate to 1 cm from nose-tip to tail-root, were measured immediately after they were sacrificed.Guard hair length and length of under hair were measured with a ruler from the hip region.
Statistical analysis
The data were analyzed by one-way ANOVA with SPSS 16.0 software and presented as the mean±SE. Data were considered significant atp<0.05 andp<0.01.
Results
In the growing period, the final BW2 of animals in group Ⅳ was significantly higher than that of animals in groupsⅠ, Ⅱand Ⅲ(p<0.01), but no significant difference was detected among groups Ⅳ, ⅤandⅥ (p>0.05). The ADG was increased linearly then reduced with increasing dietary fat levels, the ADG in groups Ⅳ and Ⅴ was significantly higher than that in groupsⅠ, Ⅱand Ⅲ (p<0.01), but no significant difference was detected among groups Ⅳ, Ⅴ and Ⅵ(p>0.05). The feed to gain ratio was reduced from 4.30 to 3.06 when dietary fat levels increased from 10%-20%, groups Ⅳ, Ⅴ and Ⅵ were significantly lower than groupsⅠ, Ⅱand Ⅲ (p<0.01), but no significant difference was observed among groups Ⅳ,Ⅴ and Ⅵ (p>0.05). The ME to gain ratio of foxes in groupⅠwas higher than that of foxes in groups Ⅱ, Ⅲ,Ⅳ, Ⅴand Ⅵ (p<0.05), and no significant difference was detected among other groups (p>0.05). In the furring period, the final BW3 of animals in group Ⅳwas significantly higher than that in groupsⅠ, Ⅱ,Ⅴ and Ⅵ (p<0.01), but no difference was detected among animals in groups Ⅲ and Ⅳ (p>0.05). The ADG of animals in groups Ⅲ and Ⅳ was significantly higher than that in groups Ⅴ and Ⅵ (p<0.01), but no difference was detected among groups I, Ⅱ, Ⅲand Ⅳ group (p>0.05). GroupsⅠ, Ⅱ, Ⅲ and Ⅳ had similar feed/gain and ME/gain ratios (p>0.05) that were significantly lower than those in groups Ⅴand Ⅵ(p<0.01) (Table 3).

Table 3 Effect of dietary fat levels on growth performance of growing-furring blue foxes
In the growing period, EE and ME increased linearly with increasing dietary fat; the digestibility of EE in group Ⅵ was significantly higher than that in groupsⅠ, Ⅱ and Ⅲ (p<0.01), and the digestibility of ME was significantly higher than that in groupsⅠ, Ⅱ,Ⅲ and Ⅳ (p<0.01). The digestibility of CP in groupⅠwas obviously lower than that in other groups (p<0.01), but no difference was detected among groups Ⅱ,Ⅲ, Ⅳ, Ⅴ and VI (p>0.05). The digestibility of DM and Ga in groupⅠwas significantly lower compared with that in other groups (p<0.01). Groups Ⅱ, Ⅲ, Ⅳ,Ⅴ and Ⅵ had similar DM and Ga digestibility values,which were positively correlated with the dietary protein levels (p>0.05). The digestibility of P in groupⅠwas significantly lower compared with that in other groups (p<0.01). In the furring period, an increased EE was observed in groups Ⅳ, Ⅴ and Ⅵ (p<0.01). The digestibility of CP in groupⅠwas significantly lower than that in other groups (p<0.01). The digestibility of ME and Ca showed no significant differences in all the groups (p>0.05). In group Ⅳ, the digestibility of DM was obviously higher than that in groups Ⅲ andⅥ (p<0.01), but no difference was detected among groupsⅠ, Ⅱ, Ⅳ and V (p>0.05). The digestibility of P in groupsⅠand Ⅱ was significantly higher compared with that in groups Ⅲ, Ⅳ and Ⅵ (p<0.01) (Table 4).

Table 4 Effect of dietary fat levels on apparent digestibility rate of nutrients of growing-furring blue foxes (%)
During the growing period, the fecal nitrogen level of groupⅠwas significantly higher than that in groupsⅣ andⅤ (p<0.01), but no difference was detected among groupsⅡ, Ⅲ and Ⅵ (p>0.05). The urine nitrogen level in groupⅠwas the highest (p<0.05).Group Ⅳ and V had higher nitrogen retention and BV than groupⅠ(p<0.05), and a significantly higher NPU than groupⅠ(p<0.01); no difference was detected among groups Ⅱ, Ⅲ and Ⅵ (p>0.05). During the furring period there was no significant difference in fecal nitrogen (p>0.05) among the groups. The urine nitrogen of animals in group Ⅵ was significantly higher than that in groups Ⅱ, Ⅲ, Ⅳ and Ⅴ(p<0.05).The nitrogen retention in group Ⅳ was significantly higher than that in group I (p<0.05) and no difference was found among groups Ⅱ, Ⅲ, V and Ⅵ (p>0.05).The NPU in group VI was significantly higher than that in groupⅠ(p<0.01). The BV of groups Ⅳ and Ⅴwas significantly higher than that in groupⅠ(p<0.05)(Table 5).

Table 5 Effect of dietary fat levels on nitrogen metabolism of growing-furring blue foxes
Pelt skin length and under hair length were significantly lower in groupⅠand group Ⅳ compared with those in other groups (p<0.01). The value of pelt width in group IV was greatly and significantly lower than that in groupⅠ(p<0.01), but no difference was detected among groups Ⅱ, Ⅲ, V and VI (p>0.05).Guard hair length did not significantly differ among the six treatment groups (p>0.05). However, guard hair length was slightly higher in group Ⅳ. The skin wide, as judged by all the above parameters, was greatly and significantly lower in group Ⅳ (p<0.01)(Table 6).

Table 6 Effect of dietary fat levels on pelt characteristics of growing-furring blue foxes (cm)
During the growing period, the optimal dietary fat levels were between 16.29% and 18.40%, according to the body weight at the end of the growing period, digestibility of phosphorus, net retention and net protein utilization, and reductions in net urine net, feed to gain and ME to gain. During the furring period, the optimal dietary fat levels were between 17.72% and 18.72%,which depended on the final body weight at pelting,net protein utilization, skin length, guard hair and under hair lengths and reduced skin wide (Table 7).
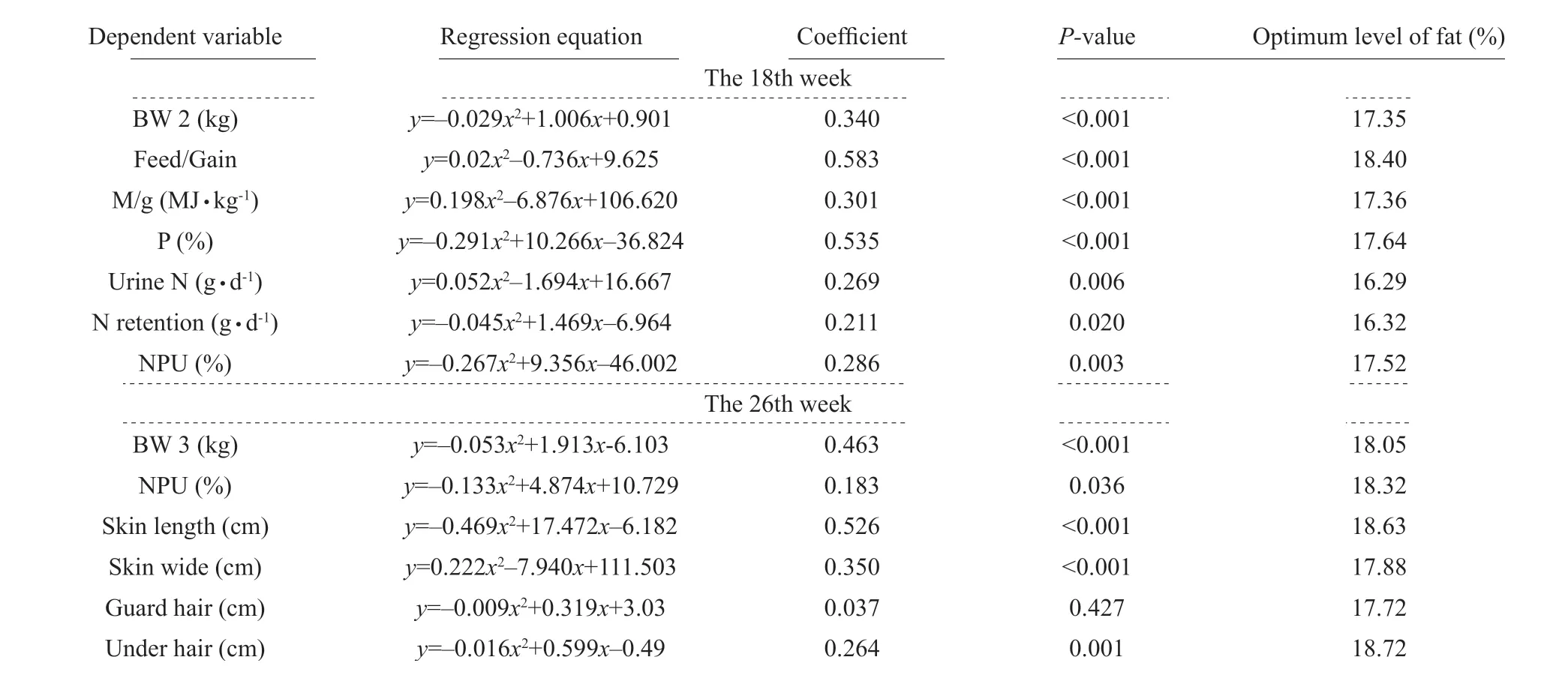
Table 7 Optimal fat levels in growing-furring blue foxes on fitting curve
Discussion
In this study, the appropriate dietary fat level efficiently accelerated growth of blue foxes and increased nutrient digestibility. Guoet al. (2014) demonstrated that the live body weight and fur length was positively correlated with BW in the fur-growing period. Gugoleket al. (2010) reported that blue foxes grow faster in the growing period (before 20 weeks of age) and grow slower during the furring period (after 20 weeks of age) which agreed with this findings. The reason was that blue foxes needed to grow during the growing period and needed to hoard fat to resist the cold during the furring period. The accumulation of body fat had dual purposes, which was crucial for animals living in the wild: to insulate the body and to serve as an energy reserve in situations of food scarcity. Kempeet al. (2009) showed that final body weight, grading size and animal length was positively correlated with fatness, most of the farmed blue foxes pelted after the growth season were either heavy or extremely fat, indicating an increased risk of obesity and related health problems. In this study, the body weights of foxes in group IV were the highest in all the groups in both the growing and furring periods but it did not find any related health problem. Blue foxes had a high tolerance to fat. Kopezewskiet al. (2001) found that the growth performance of blue foxes was improved when they were fed high fat (40%) diets. Zhanget al.(2017) reported that high fat diets decreased the feed to gain ratios of minks. Genget al. (2012) reported that the lowest feed to gain ratio was found in blue foxes fed a 54% fat diet. A diet with excessive fat led to increased energy levels, which reduced feed intake and intake of other nutrients, such as protein, which resulted in slow animal growth (Khampaet al., 2012).This study found that feed efficiency was the highest at the beginning of the experiment and the lowest at the end of the experiment. During the growing period,the feed to gain ratio of foxes fed 18% fat was lower than that in other groups and during the furring period,the group of foxes fed 16% fat had the lowest feed to gain ratio. The ME/gain ratio was increased and then decreased as increasing dietary fat levels. Difference in the feed to gain ratio and ME to gain ratio might be related to changes in body composition caused by greater fat accumulation in different growth phases.
Many factors might influence the digestibility of protein, fat, energy and other nutrients in the diets of blue foxes. The appropriate dietary fat level might improve utilization efficiency of other nutrients in diet.Fat use was generally high in fur-bearing animals,particularly with respect to unsaturated fatty acids,such as those found in raw poultry offal, poultry meals and fat. Previous studies had suggested that the average digestibility of dry matter, protein, fat and other nutrients in dogs was similar to that in blue foxes(Ahlstrom and Skrede, 1998; Vhilet al., 2005). A similar effect of decreasing dietary fat levels affecting nutrient digestibility in dogs was presented by Romsoset al. (1976) who reported that the energy digestibility values ranged from 84% to 90% when dietary fat levels ranged from 13% to 76% (Schaufet al.,2017). The digestibility of protein in the standard diet of blue fox ranged from 78% to 86%, and usually exceeded 80% (Valajaet al., 2000; Dahlmanet al.,2002; Ahlstrømet al., 2003). Additionally, studies conducted by Dahlmanet al. (2002) and Ahlstromet al.(2003) showed that digestibility coefficients of fat in blue foxes exceeded 90%. The data of this study were consistent with all the findings related to blue foxes (Ahlstromet al., 1995; Dahlmanet al., 2002;Zhanget al., 2014). Both the energy and fat digestibility tended to increase with increasing dietary fat levels. One of the reasons was that blue fox had a simple gastrointestinal tract, a relatively short intestine and a fast passage rate. Thus, to satisfy its living and reproductive needed and to develop highquality fur, it needs high energy and protein diets.During the growing period, protein digestibility was improved as fat levels increased in diets, and in the furring period, protein digestibility was increased then decreased as increasing dietary fat levels. This change in protein digestibility might be due to the high dietary fat levelsand inhibited the digestion and absorption of dietary protein in blue foxes, which resulted in excretion of protein in their feces.
The nitrogen metabolism index reflected the ratio of dietary protein utilization, some dietary proteins were digested, absorbed and used by the body for growth and development, while unabsorbed dietary proteins were excreted in urine and feces. Up to now, the effects of dietary fat levels on nitrogen metabolism in blue foxes were barely understood. Genget al. (2012)showed that the appropriate increase in the level of dietary fat decreased N excretion and the biological value of protein increased with increasing dietary fat levels during the growing-furring phase. The results of Magarethet al. (1996) showed that dietary fat supplementation improved the utilization of proteins in diets and nitrogen deposition was increased as protein digestibility increased. The findings of this study agreed to these data. Deficient and excessive dietary fat levels might increase fecal and urinary nitrogen excretion and decrease nitrogen deposition. The data from a previous study indicated that improvements in growth were due to increased fat utilization and nitrogen retention (Wuet al., 2014). At this stage,the blue foxes had reached their mature body length and had exceeded their peak hair development during the fur cycle in winter, and their demand for proteins was low. An oversupply of proteins, as well as an imbalance of amino acids, probably caused a large amount of urinary nitrogen excretion and a low nitrogen utilization rate (Mayntzet al., 2009;Damgaardet al., 2012). This study demonstrated that during the growing and furring periods of blue foxes, when the fat levels ranged from 16% to 18%,the protein digestibility was the highest. As protein digestibility was increased, protein excretion was decreased i.e., decreased excretion of fecal nitrogen and increased nitrogen deposition. Blue foxes in the growing period deposited nitrogen mainly for growth and development. Blue foxes in the furring period mainly deposited fat to cope with cold and deposit nitrogen to promote the growth of their furs. Excessive or inadequate dietary fat contents led to protein waste which reduced nitrogen deposition.
The body and dry pelt lengths in group IV were higher than those in other groups. Fur quality was the key factor that influenced fur prices. Fur prices were mostly determined by size, color, luster, density, length of guard hairs, the dry weight and cleanliness of the pelt (Liuet al., 2012). Hair growth and hair properties of fur depended on the type of dietary protein supplementation from 22 weeks to final body weight at pelting, independent of the supply of the previous period (Rasmussen and Børsting, 2000). Romsoet al. (1976) reported that most desirable fur quality was obtained when foxes were fed a diet with 27%protein and 8% fat content. Genget al. (2012) reported that animals were fed a diet with 12%-26% fat, skin size trended to increase; however, a 40% fat diet had negative effects on skin size and fur quality.Kumenoet al. (1970) suggested that pelletized feed must contain more than 15% of fat, but when foxes were fed a high-fat diet, the fur quality tended to be poor, which was similar to the results of this study.Genget al. (2012) found that foxes fed diets with 40% and 54% fat content had lower fur quality. It was found that good fur quality was achieved with dietary fat levels between 17.72% and 18.72%. Blue foxes fed diets with between 17.72% and 18.72% fat had longer skin lengths, under hair, guard hair and better fur quality in relation to other groups. Too much fat intake might lead to certain functional disorders.However, the animals in this study showed no signs of illness or maldigestion. Taken together, considering environmental protection and pelt properties, the data suggested that the fur quality of blue foxes was improved by supplementation of optimal fat level.
Conclusions
This study demonstrated that the optimal dietary fat levels for blue foxes were between 16.29% and 18.40%in the growing period and between 17.72% and 18.72%in the furring period. These fat levels promoted growth,increased the nutrient digestibility and efficiency of nitrogen use, reduced urinary and fecal nitrogen levels and improved the fur quality of blue foxes.
杂志排行

Journal of Northeast Agricultural University(English Edition)的其它文章
- Differential Responses of Phytophthora sojae to Seed Exudates of Host Soybean and Non-host Maize
- Identification of QTL and Analysis QTL with Tolerance to Sclerotinia sclerotiorum in Soybean
- Screening of Transgenic Soybean Materials for Salt Tolerance
- Development, Reproduction and Body Size Variation of Aphis glycines Matsumura Fed on Different Plants
- Effects of Bacillus subtilis on Degradation of Cellulose
- Phylogeny and Homologous Recombination Occurring in Classical Swine Fever Viruses