浙江舟山群岛山茶所在群落物种组成及多样性
2019-10-16缪玲霞王国明陈叶平高浩杰
缪玲霞,王国明,陈叶平,高浩杰,叶 波
(舟山市农林科学研究院,浙江 舟山 316000)
山茶Camellia japonica为山茶科Theaceae山茶属常绿乔木,属仅分布在沿海地区的滨海特有植物,为浙江省重点保护野生植物。自然分布于中国山东半岛、浙江沿海、台湾北部及日本和朝鲜半岛南部,舟山群岛是中国大陆山茶自然分布中心。该种既具传统的园林、观赏、生态价值,又在油用和药用方面具有极大的开发前景。多年来由于人们对野生山茶资源破坏式利用,造成天然居群数量和规模骤减,原生地保护已十分迫切。
国内外对包括舟山群岛在内的山茶天然居群的遗传多样性研究较多[1—8],对物种多样性相关研究反而鲜有报道,仅见有关青岛沿海岛屿的山茶种群结构、分布格局等种群特征的研究[9—12]。种类组成是植物群落最基本最重要的特征之一,是群落形成的基础,其多样性不仅反映群落或生境中物种的丰富度、变化程度或均匀度,也反映群落的稳定性与动态,以及不同地理自然条件与群落的相互关系[13],研究群落的物种组成及多样性是认识植物群落结构、生态、动态的基础。本文在样地调查的基础上,分析舟山群岛山茶所在群落的种类组成、区系成分、物种多样性,旨在阐明群落的结构、稳定性和生境差异,为该物种的生物多样性保护和持续利用提供科学依据。
1 研究地区自然概况
舟山群岛位于长江口以南,杭州湾外缘的东海海域,处于29°32′~31°04′N,121°31′~123°25′E,陆域面积1439.8 km2,由1390个岛屿组成,以海拔250 m以下低丘为主,最高峰为桃花岛的对峙山,海拔544.5 m。位于西南部岛屿离大陆较近,大岛多,地势高,分布密集,东北部离大陆较远,小岛多,地势低,分布稀疏。本区属中亚热带北缘季风气候区,年均温15.6~16.6 ℃,极端最低温-7.9 ℃,≥10 ℃平均积温4784~5120.8 ℃,无霜期251~303 d,年均降水量936.3~1330.2 mm,平均蒸发量1208.7~1446.2 mm,年均干湿指数0.91~1.73。丘陵山地土壤属红壤和粗骨土,大岛面海山坡和各小岛土壤盐基饱和度偏高,属饱和红壤或中性粗骨土。海岛植被属“中亚热带常绿阔叶林北部亚地带”的“浙、闽山丘甜槠、木荷林区(ⅣAiia-2)”[14],但典型的原生植被基本上已不复存在,现有丘陵山地的植被以林相残破、林分质量差的次生阔叶林和灌木林为主,常见的有枫香Liquidambar formosana林、黄连木Pistacia chinensis林、栓皮栎Quercus variabilis林、白栎Quercus fabri林及其萌生灌丛等,具代表性的常绿阔叶林有青冈Cyclobalanopsis glauca林、苦槠Castanopsis sclerophylla林及零星分布在东南部岛屿的红楠Machilus thunbergii-普陀樟Cinnamomum japonicum var. chenii林等。
2 研究方法
2.1 样地调查
在多年野外调查的基础上,选择山茶分布较为集中的普陀山、朱家尖岛、桃花岛和六横岛的阔叶林和灌丛进行群落特征调查。样地根据地形地势、群落类型和群落的连续性设置,其中阔叶林6个,阔叶林疏林1个,灌丛1个。阔叶林样地面积设为20 m×20 m,每块样地划分为4个10 m×10 m基本格子单元,阔叶林疏林和灌丛样地面积设为10 m×10 m,每块样地划分为4个5 m×5 m基本格子单元。根据演替分析和森林经营管理需要,将林下木本植物的乔木树种和灌木树种分别分为更新层和灌木层,阔叶林群落垂直结构划分为乔木层、更新层、灌木层、草本层和藤本层,灌丛群落垂直结构划分为更新层、灌木层、草本层和藤本层。各样地的基本样方面积为:乔木层10 m×10 m,更新层、灌木层和藤本层为5 m×5 m,草本层1 m×1 m。乔木层记录所有胸径(DBH)≥5 cm树种的株数、高度、胸径和盖度,更新层记录所有种的株数、高度和盖度,灌木层、草本层和藤本层记录所有种的多度、高度和盖度。同时记录海拔、坡度、坡向、土壤类型等生境因子。样地基本情况详见表1。
2.2 重要值计算
调查后分别计算各层次的重要值,其中乔木层重要值(IV) = (相对密度+相对胸高断面积+相对频度)/3;更新层重要值(IV) = (相对密度+相对盖度+相对频度)/3;灌木层、草本层、藤本层重要值(IV) = (相对盖度+相对频度)/2。相对密度= 某种的个体数/所有种总个体数×100%;相对胸高断面积= 某种的胸高断面积/所有种总胸高断面积×100%;相对盖度= 某种的盖度/所有种总盖度×100%;相对频度=某种的频度/所有种的频度之和×100%。

表1 样地基本情况表Table 1 Tree plot characteristics
2.3 物种多样性指数计算
采用物种丰富度指数、物种多样性指数和群落均匀度指数衡量物种多样性。

式中,S为每一样地每一层次中物种总数,Pi是种i的重要值Ni占所有种重要值N的比例,即Pi=Ni/N。文中采用重要值作为多样性指数的计算依据[15—16]。
3 结果与分析
3.1 物种组成
3.1.1 种类组成 在调查的8 个样地中,共记录到维管束植物65 科118 属147 种(含种下等级,下同),其中蕨类植物5 科8 属14 种,裸子植物1 科1 属1 种,双子叶植物53 科98 属121 种,单子叶植物6 科11属11 种。含4 种以上的科只有13 科,占总科数的 18.5%,其种数占总种数的44.9%;含2~3 种的科共有21 科,占总科数的33.9%,其种数占总种数的34.0%;含1 种的科多达31 科,占总科数的47.7%,其种数占总种数的21.1%(表2)。
按生活型统计,共有乔木树种39 种,包括常绿乔木24 种,落叶乔木15 种;灌木树种46 种,包括常绿灌木20 种,落叶灌木26 种;草本(含蕨类植物)30 种,包括多年生草本29 种,一年生草本1 种;藤本31 种,其中木质藤本(含攀援灌木)26 种,草质藤本(含蕨类植物)5 种,木质藤本中包括常绿藤本15 种,落叶藤本11 种;竹类1 种。各样地物种组成及其生活型见表3。

表2 8 个样地维管束植物科、属、种组成Table 2 Composition of species, genera and families of vascular plants in the eight plots
3.1.2 分布区类型 根据吴征镒等[17]对中国种子植物属分布区类型的划分,8 个样地中110 属野生种子植物的分布区类型统计见表4。结果表明,在15 种分布区类型中,仅缺乏热带亚洲至热带非洲分布型、温带亚洲分布型、中亚分布型和中国特有分布型,说明构成群落的地理成分具有多样性和复杂性,以及与世界各大洲植物区系的广泛联系,其中泛热带分布型与北温带分布型一起构成了舟山群岛山茶所在植物群落的主体。
属的分布区类型中,各种热带区系成分(2~7 项)共56 属,占54.90%(不含世界分布属)。其中以泛热带分布型占优势,计21 属,占热带成分总属数(下同)的37.50%,常见代表属有胡椒属Piper、朴属Celtis、木防已属Cocculus、榕属Ficus、黄檀属Dalbergia、花椒属Zanthoxylum、算盘子属Glochidion、冬青属Ilex、紫金牛属Ardisia、山矾属Symplocos、菝葜属Smilax 等;其次为热带亚洲分布型13 属,占23.21%,代表属有为青冈属Cyclobalanopsis、润楠属Machilus、蚊母树属Distylium、南五味子属Kadsura、山茶属Camellia、鳝藤属Anodendron 等;旧大陆热带分布型居第三位,有9 属,占16.07%,代表属有海桐花属Pittosporum、野桐属Mallotus、扁担杆属Grewia、蒲桃属Syzygium、栀子属Gardenia等;热带美洲和热带亚洲间断分布型7 属,占12.50%,为樟属Cinnamomum、楠属Phoebe、柃木属Eurya、泡花树属Meliosma 等;热带亚洲至热带大洋洲分布型6 属,占10.71%,为山龙眼属Helicia、新木姜子属Neolitsea、崖爬藤属Tetrastigma、柘属Cudrania 等。
各种温带、亚热带成分(8~15 项)共46 属,占总属数的45.10%(不含世界分布属),低于热带成分的比例。在各种温带成分中,以北温带分布型占优势,计19 属,占各种温带、亚热带成分总属数(下同)的41.30%,常见代表属有松属Pinus、桑属Morus、蔷薇属Rosa、绣线菊属Spiraea、胡颓子属Elaeagnus、黄杨属Buxus、杜鹃属Rhododendron、乌饭属Vaccinium、梣属Fraxinus、天南星属Arisaema等;其次为东亚和北美间断分布型12 属,占26.09%,代表属有石栎属Lithocarpus、枫香属Liquidambar、蛇葡萄属Ampelopsis、爬山虎属Parthenocissus、红淡比属Cleyera、络石属Trachelospermum 等;东亚(东喜马拉雅-日本)分布型12 属,占26.09%,代表属有化香树属Platycarya、檵木属Loropetalum、野珠兰属Stephanandra、野鸦椿属Euscaphis、大吴风草属Farfugium、山麦冬属Liriope 等;地中海、西亚至中亚分布型2 属,占4.35%,为黄连木属Pistacia、常春藤属Hedera;旧大陆温带分布型1 属,占2.17%,为女贞属Ligustrum。
世界分布型共有8 属,占野生种子植物总属数的7.27%,如铁线莲属Clematis、悬钩子属Rubus、堇菜属Viola、鼠李属Rhamnus、苔草属Carex 等。
分析表明,山茶所在植物群落的种子植物区系成分复杂,来源于各种地理成分,各种热带成分大于温带、亚热带成分,反映出植物区系具中亚热带区系,并由亚热带向暖温带过渡的性质。与整个舟山群岛植物区系相比,舟山群岛野生种子植物属的分布区类型中,各种热带成分占47.9%,温带、亚热带成分占52.1%,后者略占优势[18],而山茶所在植物群落的种子植物区系在属级水平上具有更为明显的热带亲缘特征,与这些群落多分布在舟山群岛东南部海边山坡或悬水小岛,热量条件相对优越有关。
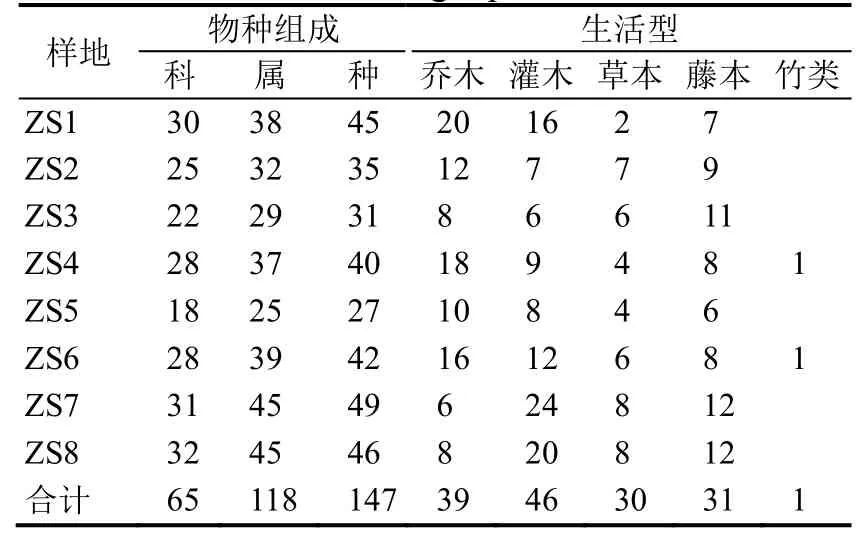
表3 8 个样地物种组成及其生活型Table 3 Floristic composition and life-form in the eight plots

表4 种子植物属的分布区类型Table 4 Areal-type of seed plant genus
3.2 物种多样性
根据各群落分层多样性指数测定结果(表5),除样地ZS8 缺乏乔木层外,乔木层最小值出现在ZS7,丰富度只有5,物种多样性指数1.320;最大值出现在ZS4,丰富度19,物种多样性指数2.497;其次是ZS6,丰富度16,物种多样性指数2.425;其他样地丰富度介于9~13,多样性指数1.783~2.216。更新层最大值出现在ZS1,丰富度为14,物种多样性指数2.185;ZS5、ZS3、ZS7 较小,丰富度4~6,物种多样性指数1.301~1.604;其他样地丰富度介于7~9,多样性指数1.420~1.843。同时ZS7、ZS8灌木层的丰富度和物种多样性指数最大,分别为(23,2.937)和(20,2.443);ZS2、ZS3、ZS5 最小,丰富度6~8,物种多样性指数1.573~1.747;其他样地丰富度介于10~16,多样性指数1.934~2.565。ZS1、ZS3、ZS5 草本层的丰富度和物种多样性指数最小,介于2~4,0.621~1.245,其他介于6~8,1.583~2.015。ZS7、ZS8、ZS3 藤本层的丰富度和物种多样性指数最大,分别为(13,2.388)、(12,1.948)、(11,2.228),其他样地丰富度介于6~9,多样性指数1.507~2.069。山茶所在的常绿落叶阔叶混交林群落(ZS4、ZS5、ZS6),乔木层的丰富度和物种多样性指数大于常绿阔叶林(ZS1、ZS2、ZS3),但在垂直结构中,更新层的丰富度和物种多样性指数明显小于乔木层和灌木层;山茶所在的灌丛(ZS8)和疏林(ZS7)群落,灌木层的丰富度和物种多样性指数显著大于其他林分,藤本层也较大。

表5 8 个样地各层次物种多样性指数Table 5 Indices of species diversity in all layers of the eight plots
通过相关性分析,不同群落各层次的丰富度指数与物种多样性指数极显著相关(r=0.908,P<0.01),而与均匀度指数的相关性不显著(r=0.011)。其中乔木层波动幅度较大,以ZS1、ZS2(0.744、0.783)最低,源于红楠、青冈或台湾蚊母树Distylium gracile 的优势特别突出,ZS3 分布最均匀(0.959)。更新层波动幅度也较大,以ZS4、ZS6(0.730,0.748)最低,二样地都因舟山新木姜子Neolitsea sericea 幼苗分布密集、优势突出,ZS5 物种分布较为均匀(0.938)。其他各样地相应层次波动幅度均较小,样地间均匀度的差异小于乔木层和更新层,稳定性相对较高。
4 讨论
4.1 物种组成特点
山茶所在群落的3200 m2样地共记录维管束植物65 科118 属147 种,其中优势科有鳞毛蕨科Dryopteridaceae、桑科Moraceae、樟科Lauraceae、蔷薇科Rosaceae、山茶科等,单种科占47.7%。各层次的优势种明显,分别为:乔木层红楠、青冈、沙朴Celtis sinensis 等,更新层山茶、红楠、舟山新木姜子等,灌木层硃砂根Ardisia crenata、茶Camellia sinensis、柃木Eurya japonica、天仙果Ficus erecta var. beecheyana 等,草本层刺头复叶耳蕨Arachniodes exilis、两色鳞毛蕨Dryopteris bissetiana、苔草Carex sp.等,藤本层络石Trachelospermum jasminoides、菝葜Smilax china、紫藤Wisteria sinensis、菱叶常春藤Hedera rhombea 等。此外还有一些特有成分,如海岛特有的普陀樟、冬青卫矛Euonymus japonicus、全缘冬青Ilex integra、小叶蜡子树Ligustrum ibota var. microphyllum、日本女贞Ligustrum japonicum、日本珊瑚树Viburnum odoratissimum var. awabuki 等,滨海特有的全缘贯众Cyrtomium falcatum、风藤Piper kadsura、舟山新木姜子、海桐Pittosporum tobira、日本野桐Mallotus japonicus、山茶、柃木等[18],反映出舟山群岛山茶所在植物群落的海岛特殊性。
4.2 分布区类型与群落生境的关系
与整个舟山群岛温带、亚热带成分略占优势的情况不同,山茶所在群落种子植物属的分布区类型热带成分高于温带、亚热带成分。其原因除调查生境单一、调查面积局限外,样地所在地一般位于舟山岛东南部岛屿的海边山坡或悬水小岛,这些生境热量条件比大岛丘陵山坡相对优越,但其他条件又相对恶劣,一些抗海风、耐盐雾的热带成分,如素馨属Jasminum、密花树属Rapanea、海桐花属、新木姜子属、山龙眼属、蚊母树属、鳝藤属等往往侵居于此,导致热带成分比例有所上升;反之像分布在舟山岛等大岛丘陵山坡的牛鼻栓属Fortunearia、鹿蹄草属Pyrola、糯米条属Abelia、万寿竹属Disporum、蜻蜓兰属Tulotis 等温带、亚热带成分不会出现在海边山坡或悬水小岛。
4.3 物种多样性与群落演替、生境的关系
物种多样性能反映植物群落的稳定性和生境的差异[19]。舟山海岛植被属“中亚热带常绿阔叶林北部亚地带”,顺行演替应从落叶阔叶林、常绿落叶阔叶混交林向常绿阔叶林演替,且渐趋稳定,一般认为物种多样性指数随演替进行而增加,但在随后的演替阶段物种多样性指数又有降低趋势。调查可见,山茶所在的常绿落叶阔叶混交林乔木层的多样性指数大于常绿阔叶林,表明群落越趋稳定,至少乔木层的多样性指数越小。王俊峰等[20]研究了福建三明常绿阔叶次生林物种多样性及群落演替之间的关系,得出3 个演替阶段(拟赤杨林、黄樟林和枫香林、栲类林)的物种多样性呈现先增加后降低的趋势;也与浙江古田山、天台山常绿阔叶林演替至稳定的甜槠林时物种多样性指数反而降低的结果相似[21—22]。同时灌木层和藤本层物种多样性也反映出植物群落生境的差异,林冠郁闭度是影响林下植物物种组成的主要环境因子[23],处于岛屿较高海拔(>300 m),且接近山顶的灌丛和稀疏矮林,由于常年强劲海风制约了乔木树种的生长及分布,林中光照充足,有利于灌木及藤本的蔓延,其物种多样性指数显著大于其他阔叶林的相应层次。
