植物乳杆菌基因yesN对中式发酵香肠风味的影响
2019-10-12张大革黄漫青高秀芝张红星徐文生
张大革 黄漫青 高秀芝 张红星 徐文生 *
(1农产品有害微生物及农残安全检测与控制北京市重点实验室 北京 102206
2北京农学院食品科学与工程学院 北京 102206)
中式发酵香肠又称腊肠,在我国具有悠久的制作历史,其制法约始创于南北朝,始见载于《齐民要术》。中式发酵香肠由于具有易于加工、口感鲜美、香味浓郁、存贮期长等特点而深受国人喜爱,是我国传统发酵肉制品的典型代表。我国不同地域饮食文化的差异,造就不同地域生产的香肠各有特色,然而制作方法大同小异,主要是在腊月以肉类为原料切成丁,配以糖、香辛料等辅料,灌入动物肠衣,经自然发酵、成熟干制而成的具有中国特色的肉制品。目前中式发酵香肠生产主要以传统自然发酵为主,存在生产受季节限制、产品质量不稳定等问题,严重制约了中式发酵香肠的发展。要拓展中式发酵香肠的发展空间,需要对传统的生产工艺进行现代化改良。
自Morris于1970年代开始研究发酵香肠微生物群落以来,科学家一直致力于揭示发酵香肠制备过程中微生物群落的发展规律。研究表明:乳酸菌对发酵香肠发酵、后熟和风味形成等起着主导作用[1];而且发酵香肠中微生物群落的发展也与香肠的加工工艺密切相关,如在加工工艺时间较短的香肠中常分离出乳杆菌;而在后熟时间长的发酵香肠中常分离出微球菌[2]等。植物乳杆菌、清酒乳杆菌等是常见于自然发酵的香肠中的乳酸菌[3],也常作为香肠发酵剂应用于发酵香肠的生产[4]。这些微生物对发酵香肠中乙酸、乙醇、丙酮酸等及其支链氨基酸降解产物如3-甲基丁醇、2-和3-甲基丁醇、3-甲基丁酸等发酵香肠典型风味成分的形成具有重要意义[2,5]。
植物乳杆菌是食品、医药工业重要的微生物[6-7],也是中式自然发酵香肠中最常见的微生物之一,对中式发酵香肠发酵和风味物质形成具有重要作用。发酵香肠中的乳酸菌也是引起发酵香肠的口感偏酸的主要原因,在制作时往往需要添加蔗糖等甜味剂改善口感。植物乳杆菌等乳酸菌常单独或混合作为发酵剂应用于发酵香肠的制备,然而生产的发酵香肠在风味上还无法与自然发酵生产的香肠竞争。
Saulnier等[8]研究表明:当蔗糖/果聚糖摄取和降解相关基因表达受到抑制时,植物乳杆菌将上调其支链氨基酸摄取及相关代谢基因的表达。而支链氨基酸、芳香族氨基酸和含硫氨基酸代谢产物对发酵香肠典型风味的形成具有重要意义。蔗糖利用缺陷的植物乳杆菌菌种作为发酵剂,可改善发酵香肠的风味,降低酸度。YesN是双组分系统YesM/YesN中的应答调控子,属于参与细菌代谢、胁迫应答和毒力因子的AraC/XylS转录调控子家族。双组分系统YesM/YesN的生物学功能目前还不清楚,主要参与细菌降解植物细胞壁多糖和细胞内糖代谢的调控[9-11]。徐文生等[12]研究表明:植物乳杆菌转录调控子基因yesN参与细胞的蔗糖代谢。植物乳杆菌SNB是本实验室通过基因敲除技术构建的一株植物乳杆菌基因yesN缺陷菌株。为研究以植物乳杆菌SNB作为模式菌株应用于中式发酵香肠发酵剂的潜力,本研究以植物乳杆菌SNB为发酵剂制备香肠,检测发酵香肠挥发性风味物质成分、酸度、总游离氨基酸等指标,并与自然发酵制备香肠和以野生植物乳杆菌为发酵剂制备香肠进行比较。
1 材料与方法
1.1 材料
菌种:植物乳杆菌WT,购自中国工业微生物菌种保藏中心;植物乳杆菌SNB,本实验室构建保存的基因yesN缺陷植物乳杆菌。
1.2 方法
1.2.1 发酵剂准备 100 mL过夜培养的植物乳杆菌菌液倒入灭菌大离心管中,5 000 r/min 5 min离心后弃上清。用灭菌生理盐水重悬菌体,再次离心弃上清,重复2次。获得的菌泥加入少量灭菌的生理盐水重悬菌体后加入香肠中。
1.2.2 香肠制备[13-14]将瘦肉、肥肉切丁后按3∶1的比例混合,并加入大曲酒、盐、味精、酱油、白糖等辅料充分搅拌混匀后分成3等份。不添加植物乳杆菌直接填充肠衣的香肠样品为空白对照(CK);添加植物乳杆菌野生菌株后填充肠衣的香肠样品记为(SEB);添加蔗糖利用缺陷植物乳杆菌菌株后填充肠衣的香肠样品记为(SNB)。制备的香肠置于恒温恒湿培养箱内发酵,发酵工艺参数为发酵温度 24℃,湿度90%~95%,发酵48 h后将温度调至20℃,湿度降至70%~80%,发酵120 h后,湿度降至60%~70%进行干燥240 h后即为成品,真空包装后贮藏于室温。
1.2.3 挥发性成分的提取[15]准确称取30.0 g剪碎的发酵成熟香肠装入100 mL样品瓶中密封。80℃水浴30 min。插入涂层为聚二甲基硅氧烷、厚度75 μm,先在GC进样口220℃解吸5 min的萃取头并推出探头,平衡30 min完成吸附。后将萃取头插入GC/MS进样器中解析5 min。
1.2.4 仪器及试验条件[13-14]气/质联用仪(GC/MS)为 Agilent 5957C-7980A。色谱柱为 DB-5(60 m×0.25 mm×0.25 μm);载气:He,流量 1 mL/min,不分流;程序升温为:初始35℃,维持5 min;升温至 100℃(升温速率 1.5℃/min),维持 5 min;升温至 200℃(升温速率 2.5℃/min),保持 5 min;最后温度升至240℃(升温速率 10℃/min),保持5 min。EI电离源,电子能量为70 eV,离子阱温度220℃;传输线温度 280℃,扫描范围:43~500 m/z,方式全扫描(Full Scan)。结果通过NIST08谱图库进行谱图成分解析;按峰面积归一化法确定发酵香肠挥发性风味物质的相对百分含量。
1.2.5 可滴定酸含量测定 取300 g剪碎后的香肠与300 mL蒸馏水置于拍打式均质袋中拍打2 min。取40 mL匀浆转移容量瓶中,70℃水浴保温45 min后定容至200 mL,过滤。取20 mL滤液加入30 mL蒸馏水,以酚酞作为指示剂,用0.1 mol/L NaOH滴定。每个样3次平行,3次重复。
1.2.6 总游离氨基酸的测定 取300 g剪碎后的香肠与300 mL蒸馏水置于拍打式均质袋中拍打2 min。取0.8 g匀浆,加入10%乙酸1 mL和6.2 mL水,混匀,冷冻离心。吸取0.5 mL上清,采用茚三酮显色法测定总游离氨基酸的含量。
1.2.7 荧光定量PCR 总RNA的提取和cDNA的合成方法分别见promega相关试剂盒使用说明书,构建16 μL RT-qPCR总反应体系包括100 ng cDNA,32 μL 引物对混合液,8 μL PCR 混合液,1.25 μL SYBR溶液,反应步骤为 95℃ 3 min;95 ℃ 10 s,58 ℃ 20 s,72 ℃ 20 s,45 个循环。检测基因scrA、sacK1和scrR的引物对分别为scrAF(TCAGCCTACATCAGGTTTG)、scrAR(GTCGGCAGTACGGACATT),sacK1F(TCGCTTCAACATCCCACTCTAC)、sacK1R(CTTTCCCCGTTCGTTTTCC)和scrRF(GTGATTTTATGTAATAGTGCGGATGA)、scrRR(CGTTGGCGAACCAGCGT)。
2 结果与讨论
2.1 蔗糖利用缺陷植物乳杆菌特性
蔗糖利用缺陷植物乳杆菌SNB是通过基因工程技术缺失植物乳杆菌ATCC14917内调控糖代谢的应答调控子基因yesN获得的突变菌株。为揭示基因yesN突变对植物乳杆菌蔗糖利用的影响,以蔗糖为唯一碳源的基础培养基中培养植物乳杆菌,其生长特性见图1。如图1所示,接种16 h后,植物乳杆菌野生菌株OD600值为0.69,而突变菌株在同样条件下OD600值仅为0.3左右。由此可见突变菌株利用蔗糖生长能力显著低于野生菌株。为确定基因yesN缺失对植物乳杆菌蔗糖利用基因座基因表达的影响,采用RT-qPCR技术检测植物乳杆菌中蔗糖代谢基因座基因scrA、scrR和sacK1的表达。结果如图2所示,相对于野生菌株,突变菌株蔗糖代谢基因座基因scrA、scrR和sacK1的表达量下降82.0%到87.0%,表明突变菌株利用蔗糖生长能力大大下降。
2.2 发酵香肠中细菌生长、总可滴定酸
中式发酵香肠发酵及其后熟均与香肠中微生物特别是乳酸菌的生长、代谢密切相关。香肠制备期间微生物指标变化见图3:第0天,样品CK和添加发酵剂的样品SEB和SNB中总细菌数大约为3.85到 4.70 lg CFU/g;到第3天,所有样品中总细菌数均达到最大水平(见图3)。乳酸菌对发酵香肠的成熟起着至关重要的作用。乳酸菌计数(见图4)显示:前3天,乳酸菌数量快速增加,从第 0天的 3.59~5.98 lg CFU/g增加到第 3天的4.95~8.65 lg CFU/g。发酵香肠中乳酸菌的生长伴随着各种有机酸特别是乳酸的生成。因此,发酵香肠中乳酸菌的快速生长常伴随着香肠酸度的增加,这样有利于快速降低香肠pH值而抑制有害微生物如肠杆菌等的生长[16]。
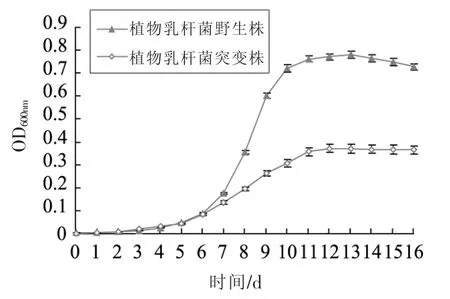
图1 植物乳杆菌野生菌株和突变菌株利用蔗糖生长特性Fig.1 The growth curve of L.plantarum WT and ΔyesN mutant strains
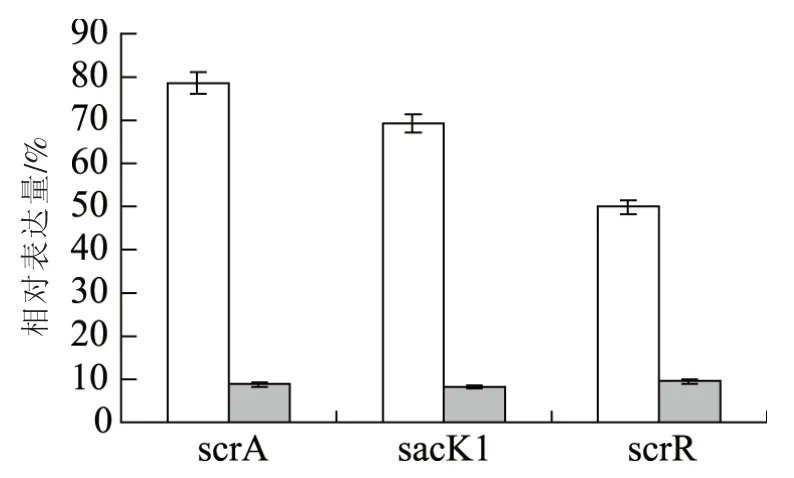
图2 植物乳杆菌野生菌株和突变菌株蔗糖利用基因座基因表达Fig.2 The expression profiles of sucrose utilization locus gens in L.plantarum WT and L.plantarum ΔyesN
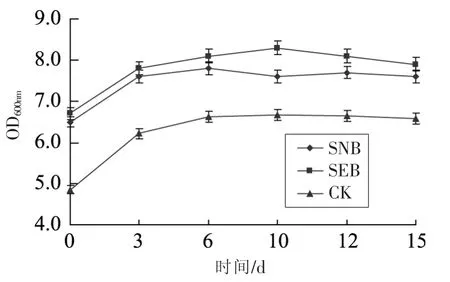
图3 不同发酵剂对发酵香肠菌落总数的影响Fig.3 The total plate count profiles of fermented sausage with different starter culture
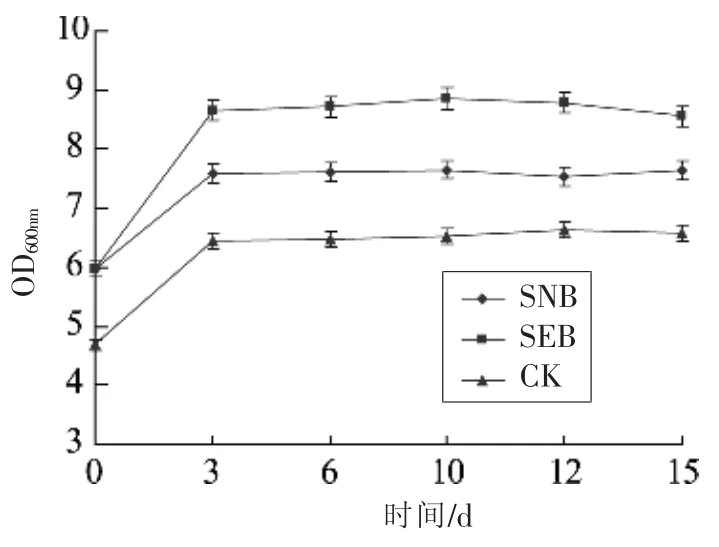
图4 不同发酵剂对发酵香肠乳酸菌总数的影响Fig.4 The total plate count of lactic acid bacterium profiles of fermented sausage with different starter culture
发酵香肠有机酸的生成可以通过检测总可滴定酸的量来分析。试验结果表明总可滴定酸的含量在发酵期间快速增加(见图5),第3天时达到0.35~0.50 mg/100 g,其中样品SNB与样品CK总可滴定酸的含量比较接近,与样品SEB中总可滴定酸含量差异显著。发酵香肠由于乳酸菌产生大量乳酸等有机酸一般口感偏酸,需要大量添加蔗糖等作为微生物的碳源和甜味剂。蔗糖等甜味剂的使用既可以促进乳酸菌等有益微生物的快速生长,也可以改善香肠的口感。但蔗糖的大量使用已经不符合现代食品低糖、低脂的发展方向。因此,相对于使用野生菌株作为发酵剂,使用模型菌株更有利于降低蔗糖的使用。
中式发酵香肠的总游离氨基酸含量见图6,发酵香肠总游离氨基酸含量从第1天的19.80 mg/100 g快速增长到第3天的66.75~84.67 mg/100 g,到第 15 天增加至 79.89~120.69 mg/100 g之间。发酵香肠的生产工艺对发酵香肠的总游离氨基酸含量有着一定的影响,如有些学者报道总游离氨基酸的含量在发酵阶段快速增加,而有些研究则报道总游离氨基酸在后熟阶段快速增加[16]。肉类蛋白的水解能力主要来源于内源性或微生物蛋白酶,不同的乳酸菌拥有不同的蛋白裂解能力[17-18],在发酵阶段总游离氨基酸的快速增加意味着在这个阶段相关酶活性最高。在本研究中,样品SNB和SEB中蛋白裂解能力显著低于样品CK(P<0.01)。而样品SNB中总游离氨基酸的含量也显著高于样品 SEB(P<0.01)。
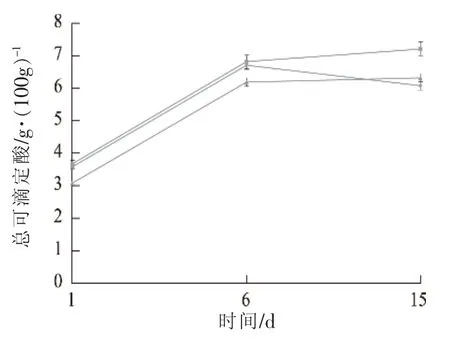
图5 不同发酵剂对发酵香肠总可滴定酸的影响Fig.5 The total titratable acid profiles of fermented sausage with different starter culture
2.3 风味成分
通过顶空固相微萃取技术从香肠中提取的挥发性风味成分见表1,共检测到包括7种酸、18种醛、3种酮、7种醇、10种酯等46种挥发性风味成分,其中醛和乙酯类成分占主导地位。脂肪氧化产生的各种挥发性成分如各种饱和脂肪醛(己醛、庚醛、壬醛),不饱和脂肪醛(2-己醛、2,4-庚二醛、2-辛烯醛、2-壬烯醛、2,4-壬二烯醛、2-癸烯醛等),酮(4-羟基丁酮、3-羟基丁酮),醇(1-辛烯-3-醇、3-甲基丁醇)等是发酵香肠风味形成的重要基础,这些物质对发酵香肠挥发性风味特征的形成具有重要意义,也是发酵香肠不良风味的重要来源[11,19-20]。一般而言,添加植物乳杆菌发酵香肠与传统发酵香肠在挥发性风味成分上存在显著差异,特别是产生的不良风味物质相对于传统工艺发酵香肠显著减少[21]。
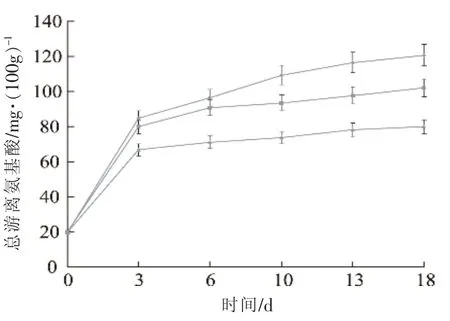
图6 不同发酵剂对发酵香肠总游离氨基酸的影响Fig.6 The profiles of total free amino acid in fermented sausage with different starter culture
2.3.1 来源于氧化作用的风味成分 醛类成分能够形成广泛的风味且具有较低的感觉阈值,是发酵香肠中来源于脂肪氧化的最受关注的一类风味成分[22-24]。不同生产工艺制备的发酵香肠中,醛类的种类和占总挥发性风味物质的含量有所不同[25-27],含有6~8个碳原子的脂肪醛,如庚醛、壬醛和己醛常作为脂质氧化的标志,是发酵香肠中重要的挥发性醛类[25]。苯甲醛和苯乙醛是亮氨酸和苯丙氨酸代谢的标志性产物。苯甲醛有烤花生的香味,苯乙醛产生的香味与山楂相似[27-29]。己醛具有一种难闻的青草味,由不饱和脂肪酸氧化降解形成醛类[26],庚醛拥有典型的土豆味而壬醛则拥有典型的水果香味。
酸类在3种样品中大约占到总离子峰的2.98%~6.32%。长链(C14-C18)和中链(C6-C12)脂肪酸在3种发酵香肠中均有分布,它们主要是从甘油三酯、磷脂或油脂降解形成[20]。长链或中链脂肪酸本身对提高发酵香肠风味没有作用,主要是作为风味成分的前体物质,通过进一步降解形成不同风味物质影响发酵香肠的口感或香味[30]。短链脂肪酸(C<6)一般检测阈值较低且具有浓郁的干酪味,是影响中式发酵香肠的主要成分之一。在本研究中,样品SEB中短链脂肪酸的含量显著高于(P<0.01)样品CK和SNB。有些研究者发现在发酵香肠中存在乙酸[30],但本研究样品中没有发现。
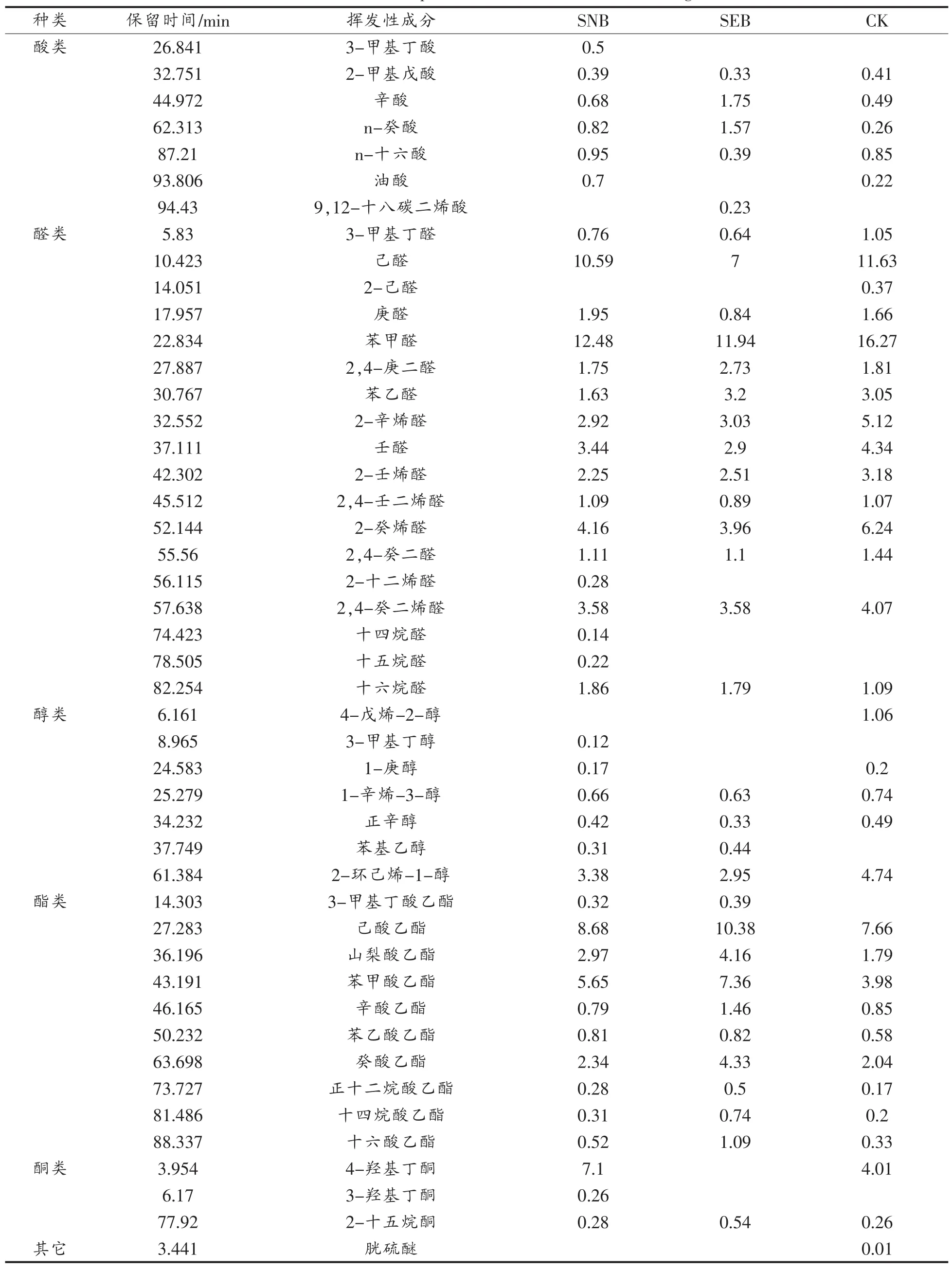
表1 发酵香肠风味成分分析Table 1 Volatile component of Chinese fermented sausage
2.3.2 来源于氨基酸降解的挥发性风味物质 醛类(苯甲醛、苯乙醛)、醇类(3-甲基丁醇,苯乙醇)、酸类(3-甲基丁酸)、酯类(3-甲基丁酸乙酯、苯酸乙酯)均检测到。3-甲基丁醛、3-甲基丁醇、3-甲基丁酸、3-甲基丁酸乙酯均来源于支链氨基酸降解。本研究在样品SNB中,来源于支链氨基酸降解的风味成分的相对含量显著高于样品SEB和CK,其机理可能与在植物乳杆菌中蔗糖能够抑制支链氨基酸摄取和降解代谢基因表达有关[8,31]。发酵香肠中添加氨基酸并不能显著增加发酵香肠的典型风味,氨基酸特别是支链氨基酸、芳香族氨基酸和含硫氨基酸通过微生物的降解代谢,才能够形成发酵香肠中浓郁香味[16]。
2.3.3 来源于微生物代谢的挥发性成分 微生物代谢活性在发酵香肠中与氨基酸降解、碳源发酵、酯类的β-氧化和酯酶活性有关[32]。本研究检测发酵香肠中来源于微生物代谢碳源的成分主要有4-羟基丁酮和3-羟基丁酮。微生物代谢形成的挥发性风味成分的浓度受微生物的显著影响[27,33],在样品CK和SNB中,碳源代谢产物的离子峰面积显著高于样品SEB。酮类成分主要来源于微生物的β-氧化,样品SNB中酮类的相对含量显著高于样品 SEB(P<0.01)。
酯类是发酵香肠中第二丰富的挥发性风味成分[20]。由于酯类具有较低的检测阈值和水果香味,酯类被认为是中式发酵香肠挥发性风味的重要组成成分[25-27]。许多乙酯类成分如己酸乙酯、辛酸乙酯在3种样品中均有分布,且在其它发酵香肠中也有发现[12,30,34-35]。而来源于氨基酸代谢的 3-甲基丁酸乙酯则仅在样品SNB和SEB中发现。
3 总结
总之,添加植物乳杆菌基因yesN缺陷菌株制备香肠在检测到的挥发性风味成分,特别是典型挥发性风味成分方面,明显优于传统发酵工艺生产的发酵香肠,而在总可滴定酸、总游离氨基酸等指标方面则明显优于添加野生菌株的植物乳杆菌菌株。虽然突变菌株涉及基因修饰微生物而未进行品评试验,但研究表明蔗糖利用缺陷植物乳杆菌具有作为改良中式发酵香肠发酵工艺的模式菌株,应用于筛选中式发酵香肠发酵菌株的潜力。
