河套灌区不同秋浇年限对土壤细菌群落的影响
2019-10-12张晓丽张宏媛卢闯逄焕成靳存旺高喜程挨平李玉义
张晓丽,张宏媛,卢闯,逄焕成,靳存旺,高喜,程挨平,李玉义
河套灌区不同秋浇年限对土壤细菌群落的影响
张晓丽1,张宏媛1,卢闯1,逄焕成1,靳存旺2,高喜3,程挨平3,李玉义1
(1中国农业科学院农业资源与农业区划研究所,北京 100081;2五原县农牧业技术推广中心,内蒙古五原 015100;3内蒙古河套灌区义长灌域管理局义和渠管理所,内蒙古五原 015100)
【】针对内蒙古河套灌区农民自愿放弃秋浇造成干地面积逐渐增多的现象,通过调查研究不同秋浇年限对盐碱土壤细菌群落组成差异的影响,为灌区盐碱地改良与秋浇制度改革提供科学依据。选取盐荒地(CK)、一直秋浇(AUI1)、隔2—3年秋浇(AUI2)、3—4年不秋浇(AUI3)以及6—7年不秋浇(AUI4)5种类型的典型地块,采用高通量测序(Illumina HiSeq)技术,分析不同秋浇年限处理下土壤细菌群落的变化特征,并对土壤化学性质与细菌群落进行了冗余分析以及与群落组成的相关性分析。AUI1和AUI2处理对降低0—30 cm土层盐分含量最显著,分别比CK、AUI3、AUI4降低了128.82%和29.04%、108.76%和17.72%、108.44%和17.55%,在30—40 cm土层AUI1处理的盐分含量显著低于其他处理;各个土层的pH值均以AUI2处理最低,其0—40 cm土层pH平均值较CK、AUI1、AUI3、AUI4处理分别显著降低了0.28、0.32、0.16、0.88个单位(<0.05)。另外,AUI2处理0—40 cm土层微生物量碳平均含量分别较CK、AUI1、AUI3、AUI4提高了252.89%、148.59%、58.10%和60.10%,可溶性有机碳平均含量分别提高了48.41%、29.42%、6.01%和14.27%(<0.05)。AUI1与AUI2处理间的丰富度指数(ACE和Chao1)显著高于CK、AUI3和AUI4处理(<0.05)。变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)和拟杆菌门(Bacteroidetes)为不同秋浇年限处理的3大优势菌门,占所有菌门的53.93%,而AUI2、AUI3处理分别有利于增加拟杆菌门、酸杆菌门的丰度(<0.05)。相关性分析结果表明,变形菌门(Proteobacteria)和绿弯菌门(Chloroflexi)与土壤微生物量碳和可溶性有机碳均具有很强的负相关性,其中与微生物量碳的相关系数分别为(=-0.559**和-0.522*),与可溶性有机碳的相关系数分别为(=-0.795**和-0.820**);而放线菌门(Actinobacteria)、奇古菌门(Thaumarchaeota)、厚壁菌门(Firmicutes)和疣微菌门(Verrucomicrobia)均与土壤盐分呈显著负相关,与土壤微生物量碳和可溶性有机碳呈显著正相关。因子分析显示,土壤盐分、pH、微生物量和可溶性有机碳总共解释了97%的群落变化,成为土壤细菌群落组成变化的主控因子,贡献率依次为:土壤盐分>pH>微生物量碳>可溶性有机碳。隔2—3年秋浇(AUI2)处理不仅可以有效降低耕层土壤盐分并能使0—40 cm土层微生物量碳和可溶性有机碳含量显著增加,而且还可提高土壤优势菌群中拟杆菌门的相对丰度。综上所述,隔2—3年秋浇(AUI2)是河套地区兼顾盐碱土壤改良与节水的优化措施。
秋浇;土壤细菌;群落结构;多样性;盐碱土;高通量测序;内蒙古河套灌区
0 引言
【研究意义】内蒙古河套地区是我国西部地区重要的粮油基地,同时也是我国重要的生态屏障。河套地区地处生态环境脆弱区域,极高的蒸降比、封闭地形以及长期引黄灌溉造成严重的土壤次生盐渍化,盐碱化耕地比例接近50%[1-3]。“秋浇”是河套地区特定的自然气候条件下,为适应当地土壤耕作、作物种植、盐碱地改良和水资源合理利用,在长期经验积累中形成的一种有效灌溉制度[4],其目的是在于通过每年10—11月份进行的一次大规模灌溉淋洗田间盐分和调整土壤墒情,为来年春播创造良好的土壤水盐条件。但是,近年来河套地区受经济利益的驱动,种植结构发生了很大变化,特别是短生育期的向日葵种植面积大幅度增加,使得部分农民不再秋浇,或改秋浇为春灌等等[5]。目前,不秋浇的势头越来越猛,造成干地面积逐年大幅度增加。据统计,河套地区现有灌溉面积71.5万hm2,自2014年开始每年秋浇干地面积基本保持13.3万hm2左右[6]。灌溉制度改变后多年形成的水盐动态平衡打破,必然影响整个河套地区水盐平衡状况,导致排入乌梁素海的总盐量减少,而留在灌区内的总盐量增加,长此以往可能会引起耕地盐碱化程度加剧[1]。土壤微生物在一定程度上能够反映土壤质量,通过调查不同秋浇年限对土壤环境和微生物群落多样性及组成的影响,确定一个合理的秋浇间隔年限,以期为河套灌区盐碱地改良与秋浇制度改革提供科学依据。【前人研究进展】有研究表明,秋浇后的土壤温度适宜,在微生物的作用下可以加速枯枝败叶的腐烂,有利于提高土壤肥力,减轻病虫危害[6]。但是随着秋浇的发展以及灌水量的增大,使之排水不畅,造成地下水位大幅度上升以及次生盐碱化问题[3]。土壤的盐碱度变化直接影响其微生物的数量及群落结构。我国学者在研究滨海和内陆盐碱地的微生物数量时发现,随着土壤盐碱化程度的加重,土壤细菌、真菌和放线菌数量及微生物总数呈现显著降低的趋势[7-9]。而对于盐碱地来说一个适度的灌溉量是微生物数量增长的必要条件,过少和过多的灌溉量均不利于微生物繁殖[10]。目前对于河套地区盐碱土壤秋浇的研究集中在节水潜力、脱盐效果、养分淋溶等方面[11-13],而秋浇制度变化对盐碱土壤微生物方面的研究鲜见报道。【本研究切入点】细菌是土壤微生物中最主要和最活跃的类群,直接参与土壤中物质和能量的转化与循环。目前有关不同盐碱程度对土壤细菌群落结构及组成的影响研究较多[14-15],但针对不同秋浇年限条件下土壤细菌群落多样性及其组成,以及秋浇引发的细菌群落组成差异与土壤环境因子的关系研究鲜见报道。【拟解决的关键问题】本研究以盐荒地为对照(CK),通过选择不同秋浇年限地块,采用高通量测序技术明确不同秋浇年限对土壤细菌群落多样性及组成的影响,阐明不同秋浇年限农田土壤细菌群落组成差异及其与土壤盐分、pH、微生物量碳、可溶性有机碳等的相互关系,研究结果可为河套灌区盐碱地改良与秋浇制度改革提供相应的科学支撑。
1 材料与方法
1.1 研究区概况
研究区域位于内蒙古五原县胜丰镇(41.07°N、108.00°E,海拔1 022 m),该地区属于中温带大陆性气候,全年日照时数3 263 h,年均温6.1℃,≥10℃的积温3 392℃,无霜期117—136 d。该地区位于河套平原腹地,处于黄河灌溉区,具有光能丰富、日照充足、昼夜温差大、降雨量少而集中的特点,土壤类型以粉砂壤土为主。
1.2 试验处理
2017年9月20日,作物收获后且秋浇未开始前,在农田基本理化性状一致的典型示范区中,以盐荒地为对照(CK),选取一直秋浇(AUI1)、隔2—3年秋浇(AUI2)、3—4年不秋浇(AUI3)、6—7年不秋浇(AUI4)等5种不同秋浇年限处理的地块,每块样地667 m2以上,为确保取样点的代表性,取样时包含了路边、沟渠与田埂的结合部等环境。样点分布示意图如图1所示:即不管地块是否紧邻沟渠或路边,都会采取如下等距离法进行取样,每个点用土钻取0—10、10—20、20—30和30—40 cm 4个土层的土壤样品,取完分别将1、5、7、9、11;3、6、10、13、15;2、4、8、12、14不同土层的土壤样品混在一起,视作3次重复。
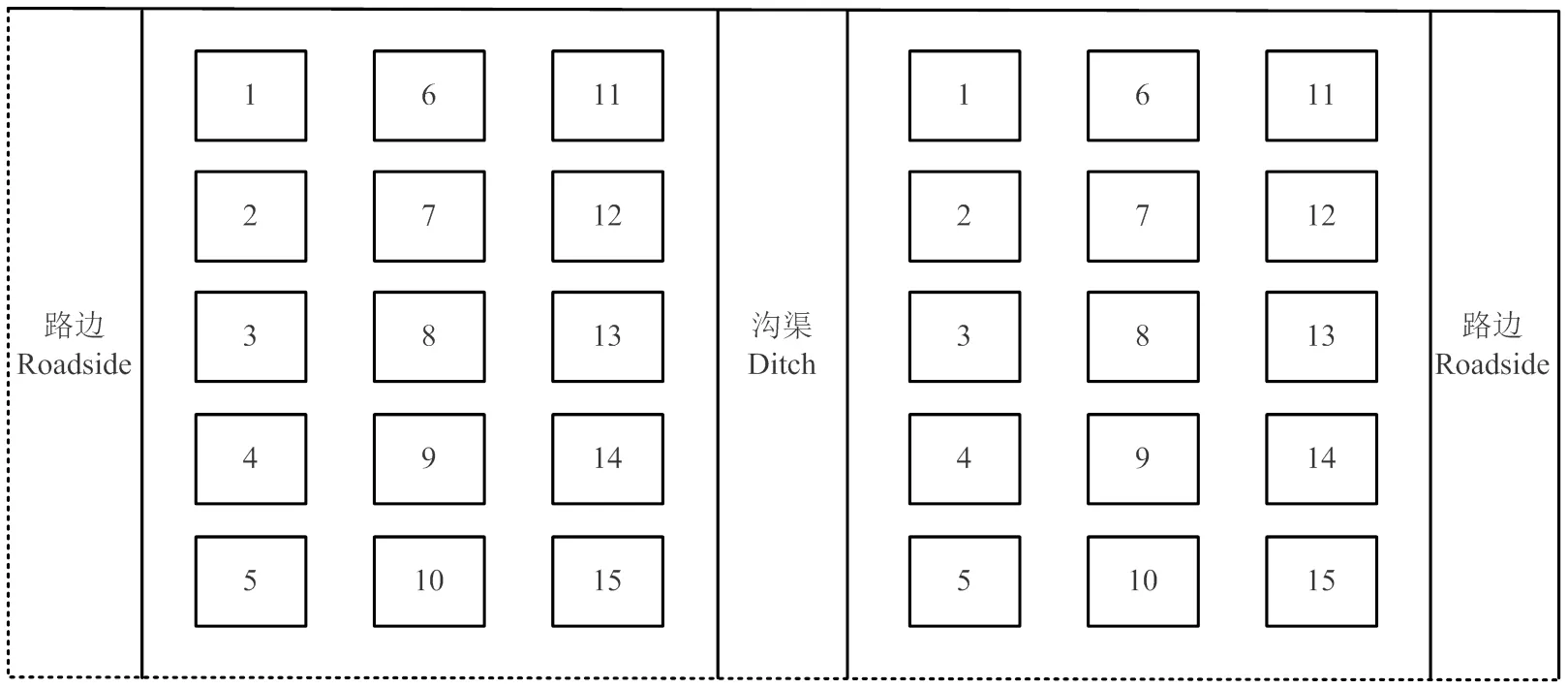
图1 样点分布示意图
1.3 取样方法与数据测定
1.3.1 土壤盐分、pH的测定 作物收获后,在两行作物中间位置(地膜下)取土进行土壤盐分、pH测定,取土层次为0—10、10—20、20—30、30—40 cm,带回实验室自然风干,磨碎,过2 mm筛,以1﹕5的土水比提取土壤溶液上清液,用pH计(FE20)和电导率仪(DDS-307)分别测定土壤pH和电导率,电导率根据经验公式换算为土壤盐分含量。
土壤盐分含量(g·kg-1)=电导率×0.064×5×10/1000
1.3.2 土壤可溶性有机碳、微生物量碳含量的测定 作物收获后,取0—40 cm土层的土壤样品,每10 cm一层,用移动保温箱带回实验室。土壤微生物量碳(MBC)和可溶性有机碳(DOC)采用氯仿熏蒸-K2SO4提取法测定[16]。以不熏蒸土样提取的有机碳作为土壤可溶性有机碳(DOC);以熏蒸土样与不熏蒸土样提取的有机碳差值除以转换系数KC(本研究取值为0.38)计算土壤微生物量碳(MBC)。
1.3.3 土壤细菌群落测定 在食葵收获后取0—30 cm土层土壤样品,将土样放入牛皮纸袋并迅速保存于4℃冷藏箱带回实验室进行土壤细菌群落的测定。称取约0.33 g经冷冻干燥处理的土壤样品,使用 Fast DNA Spin Kit for Soil DNA 提取试剂盒(MP Biomedicals,USA),参照试剂盒使用说明提取样品中的总DNA。并用1% 凝胶电泳检测提取的DNA质量,同时用Nano Drop 2000 UV-Vis Spectrophotometer(Thermo Fisher Scientific, USA)测定浓度。
细菌使用16S rRNA基因通用引物515F(5¢-GTGCCAGCMGCCGCGGTAA-3¢)和 907R (5¢-CCGTCAATTCCTTTGAGTTT-3¢)进行目的基因扩增。PCR反应条件为:98℃预变性1 min,98℃变性10 s,50℃退火30 s,72℃延伸30 s,共30个循环,最后在72℃下延伸5 min。反应产物用2%琼脂糖凝胶电泳检测。通过Hiseq 2500平台(Illumina,SanDiego,CA,USA)进行测序(诺禾致源生物信息科技有限公司,北京)。
原始序列使用 Pandaseq(V 2.7)软件进行拼接,再用 Trimmomatic(V 0.30)软件去除引物和接头序列,使用 Usearch(V 8.0)软件将拼接过滤后的序列与数据库进行比对,去除嵌合体序列(chimera sequence),最终得到有效序列数据。利用 RDP classifier 贝叶斯算法对97% 相似水平上进行操作分类单元(operational taxonomic units,OTUs)聚类[17-19],根据 OTUs聚类结果,对每个 OTUs的代表序列做物种注释,得到对应的物种信息和基于物种的相对丰度分布情况,同时,对OTUs进行相对丰度、Alpha多样性计算等分析[19],得到样品内物种丰富度和均匀度信息、不同样品或分组间的共有和特有OTUs信息等。
1.4 数据处理
采用Microsoft Excel 2010进行数据处理与图表制作,使用DPS 6.55软件进行数据的方差比较,采用LSD法进行差异显著性检验(<0.05)。使用CANOCO4.5 软件进行土壤化学性质与细菌群落组成的主分量分析(PCA)以及对土壤化学性质和细菌群落多样性进行了冗余分析(RDA);土壤微生物群落组成与土壤理化性质之间的 Pearson 相关性分析采用 SPSS 13.0 计算。图表数据均为 3次重复的平均值±标准误。
2 结果
2.1 不同秋浇年限对土壤盐分、pH及微生物量碳与可溶性有机碳含量的影响
2.1.1 土壤盐分 由表1可以看出,AUI1处理的盐分含量整体偏低,主要聚集在土壤表层0—10 cm处,随着土层的加深盐分含量基本呈下降趋势;AUI2和AUI3处理的盐分含量随着土层的加深在逐渐增大;而CK和AUI4处理的盐分表聚性较强。
在0—10 cm土层,AUI1处理的盐分含量最低,为1.57 g·kg-1,显著低于其他4个处理,其次盐分含量比较低的是AUI2处理,而盐分含量最高的是CK和AUI4处理;在10—20 cm土层,盐分含量的高低顺序是CK>AUI4>AUI3>AUI2>AUI1处理,且AUI1处理的盐分含量分别比CK、AUI4、AUI3和AUI2处理显著降低了143.82%、176.31%、192.45%和218.40%(<0.05);在20—30 cm土层,盐分含量最低的仍为AUI1处理,其次为AUI2处理,接下来是CK和AUI4处理,而盐分含量最高的为AUI3处理,为2.29 g·kg-1,且AUI1、AUI2、AUI4和CK处理的盐分含量分别比AUI3处理显著降低了188.13%、15.21%、11.61%和8.32%(<0.05);在30—40 cm土层,盐分含量的高低顺序为AUI1<AUI4<CK<AUI3<AUI2处理,AUI1、AUI3、AUI4和CK处理的盐分含量分别比AUI2处理显著降低了313.47%、11.36%、70.79%和41.00%(<0.05)。且在每个土层各个处理间的盐分含量是有显著差异的。
总体来说,在0—30 cm土层处,AUI1和AUI2处理的盐分含量最低,而在30—40 cm土层处AUI1处理的盐分含量依然是最低的,可见除了一直秋浇(AUI1)是降低盐分最好的措施之外,隔2—3年秋浇处理(AUI2)对于降低0—30 cm土层的盐分是可行的。
2.1.2 土壤pH 收获后各处理0—40 cm土层pH值的变化如表1所示,各处理在0—10 cm表层存在显著差异,其中以AUI2处理的pH值最低,较CK、AUI1、AUI3、AUI4处理分别显著降低了0.29、0.19、0.11、0.78个单位(<0.05),AUI1与AUI3之间没有显著差异;在10—20和30—40 cm土层,除了CK与AUI1处理间的pH值无显著差异外,其他各处理间均存在显著差异,其中AUI2处理的pH值最低,分别平均较CK、AUI1、AUI3、AUI4处理分别显著降低了0.43、0.66、0.25、1.72个单位(<0.05);而在20—30 cm土层,AUI2处理的pH值较CK、AUI1、AUI3、AUI4处理分别显著降低了0.10、0.25、0.12、0.87个单位(<0.05);CK与AUI3处理间差异不显著。总的来说各个土层的pH值均以AUI2处理最低,其均值为8.33,其次为AUI3、AUI1、CK处理,而AUI4处理的pH值最高且为9.22,AUI2处理的pH值较CK、AUI1、AUI3、AUI4处理在0—40 cm土层分别显著降低了0.28、0.32、0.16、0.88个单位(<0.05)。
2.1.3 土壤微生物量碳与可溶性有机碳 由表1还可以看出,各处理土壤微生物量碳含量在0—10 cm土层依次表现为AUI2>AUI1>AUI4>AUI3>CK且AUI2处理的微生物量碳含量较AUI1、AUI3、AUI4和CK处理显著增加了13.10%、63.82%、13.83%和78.52%(<0.05);而在10—40 cm土层,AUI1处理的微生物量碳含量最高,AUI2处理次之,AUI2处理较AUI3、AUI4和CK处理分别平均显著增加了54.38%、86.41%、203.18%(<0.05);除了AUI1处理的微生物量碳含量是随着土层的加深在逐渐增大,且在20—30 cm土层处达到最大外,其他各处理的微生物碳含量都随着土层的加深在减小,并且除0—10 cm表层的AUI2和AUI4处理外,各个处理在各个土层间的微生物碳含量的差异显著。
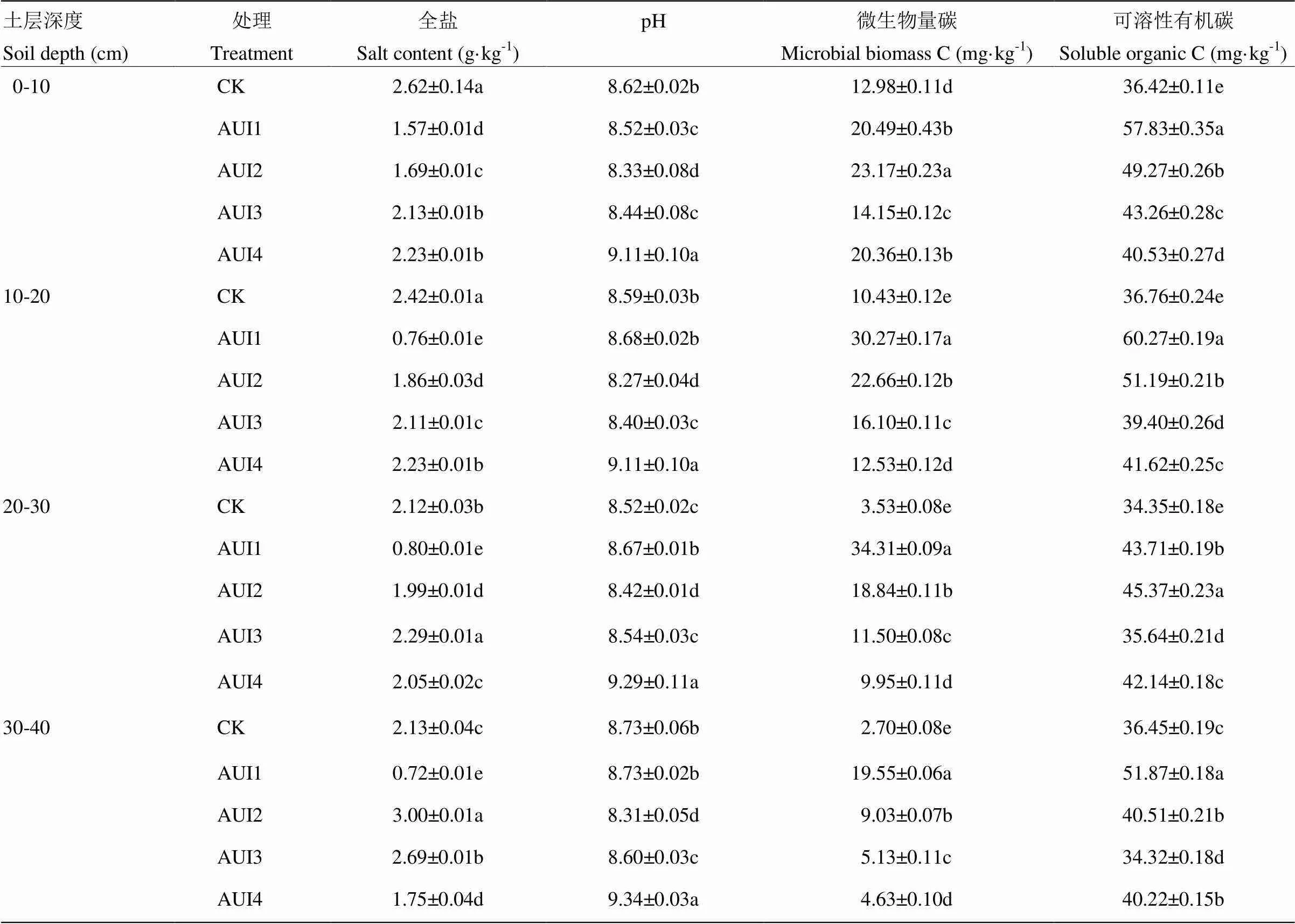
表1 收获后不同处理0—40cm土层盐分、pH及活性有机碳含量
土壤可溶性有机碳含量在0—20和30—40 cm土层表现为AUI1处理最高,AUI2处理次之,且AUI1和AUI2处理的土壤可溶性有机碳含量在0—20和30—40 cm土层平均较CK、AUI3和AUI4分别提高了55.04%和28.58%、45.29%和20.49%、38.89%和15.19%;各处理土壤可溶性有机碳含量在20—30 cm土层依次表现为AUI2>AUI1>AUI4>AUI3>CK且AUI2处理较CK、AUI1、AUI3、AUI4处理显著增加了32.09%、3.80%、27.30%和7.65%(<0.05)。
2.2 不同秋浇年限处理的高通量测序方法分析
2.2.1 土壤细菌多样性分析 通过高通量测序,AUI1、AUI2、AUI3、AUI4、CK 5个样品原始序列条数分别为92 266、89 039、89 639、95 582、80 806,过滤掉低质量的序列后,有效序列分别为89 950、87 116、87 947、93 294、78 958。将这些序列在 97%相似度聚类为用于物种分类的 OTU,然后通过最小样本序列数对其进行抽平,再通过随机抽样的方法抽取一定数量的个体,以抽到的序列数与它们所代表的 OTU 数目构建稀释性曲线。从图2可知,OTU 数均随序列的增加而增大,不同处理之间表现为CK>AUI4>AUI1>AUI2>AUI3。随 OTU数的增大,5种处理细菌稀释曲线均基本趋于平缓,但仍未达到饱和,说明测序数据量渐进合理,更多的测序数据对发现新的OTU贡献率较小,另外,测序覆盖率均高于90%,也表明本次测序深度合理,基本能代表样本的真实情况(图2)。
微生物多样性是一个群落稳定性的标尺,反映环境的生态机制和外界胁迫对群落的影响。通过表2所示的不同处理土壤样品细菌多样性发现,各个处理的土壤细菌群落Shannon、Simpson指数差异均不显著(>0.05)。而AUI1、AUI2、AUI3处理的土壤细菌群落Chao1和ACE丰富度指数分别较CK显著提高了30.14%和29.24%、14.02%和12.51%、9.93%和8.95%,较AUI4显著提高了28.63%和28.41%、12.69%和11.79%、8.65%和8.26%(<0.05),且CK与AUI4处理间差异未达到显著水平,而AUI1、AUI2、AUI3处理间存在显著差异。
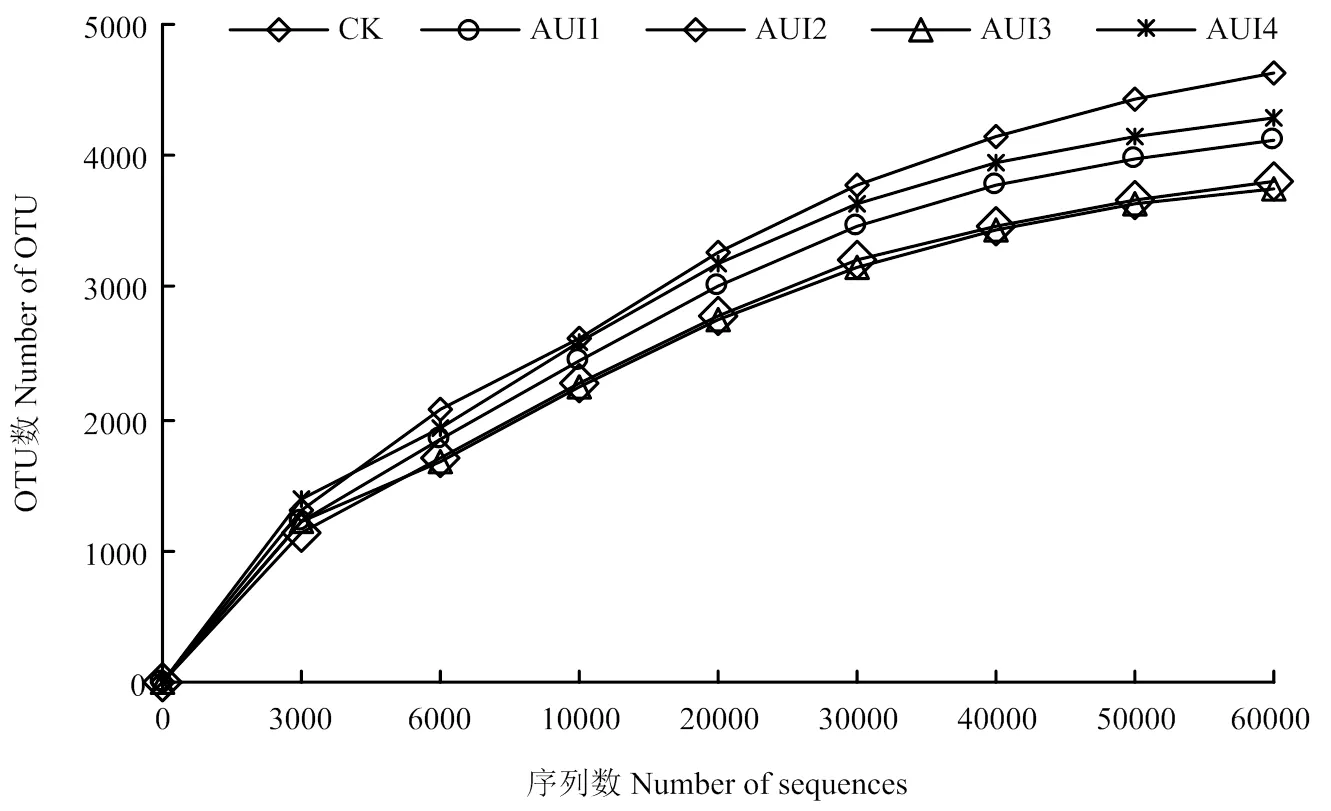
图2 细菌稀释曲线分析

表2 不同处理细菌群落多样性分析
2.2.2 土壤细菌群落组成 由门水平的细菌群落组成(图3)可知,变形菌门(Proteobacteria)相对丰度最高,占19.98%—34.01%,平均为25.57%,其次为酸杆菌门(Acidobacteria),占12.22%—23.48%,平均为15.97%,随后依次为拟杆菌门(Bacteroidetes,7.98%—12.34%)、芽单胞菌门(Gemmatimonadetes,6.53%—12.97)、放线菌门(Actinobacteria,8.02%— 10.48%)、奇古菌门(Thaumarchaeota,1.05%—8.00%)、绿弯菌门(Chloroflexi,4.05%—7.98%)、浮霉菌门(Planctomycetes,2.30%—6.55%)、疣微菌门(Verrucomicrobia,1.89%—4.52%)、厚壁菌门(Firmicutes,1.05%—2.61%)。CK处理中只有变形菌门的相对丰度均高于其他4个处理,而其余菌门在AUI1、AUI2、AUI3和AUI4处理间的响应不同。具体表现为,AUI1处理有利于增加放线菌门、奇古菌门、疣微菌门和厚壁菌门的相对丰度;AUI2处理有利于拟杆菌门和芽单胞菌门的相对丰度;AUI3处理有利于增加酸杆菌门、绿弯菌门和浮霉菌门的相对丰度;而AUI4处理不利于增加任何一门细菌的相对丰度。说明在细菌群落组成前10的菌门中,AUI3处理增加了相对丰度较大的拟杆菌门和相对丰度较小的绿弯菌门和浮霉菌门;AUI1处理增加了相对丰度居中和最小的放线菌门、奇古菌门和疣微菌门、厚壁菌门;而AUI2处理增加了相对丰度第三和第四的拟杆菌门和芽单胞菌门相对丰度。
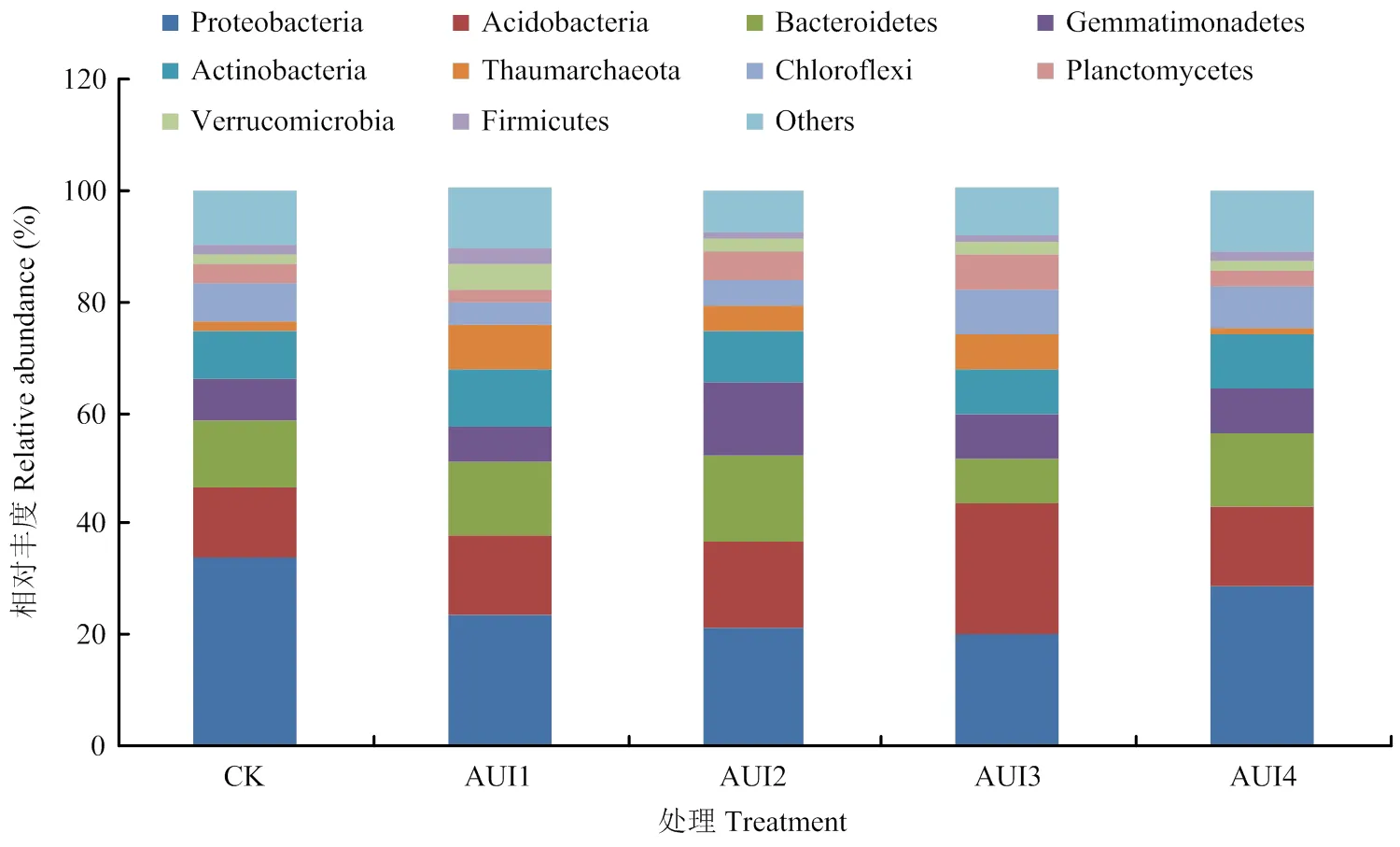
Proteobacteria:变形菌门,Acidobacteria:酸杆菌门,Bacteroidetes:拟杆菌门,Gemmatimonadetes:芽单胞菌门,Actinobacteria:放线菌门,Thaumarchaeota:奇古菌门,Chloroflexi:绿弯菌门,Planctomycetes:浮霉菌门,Verrucomicrobia:疣微菌门,Firmicutes:厚壁菌门,Others:其他
2.2.3 影响不同秋浇年限土壤细菌群落的因子分析 由土壤化学性质参数与细菌群落组成的主分量分析(图4-a)可知,相同处理的土壤都聚集在一起,且不同处理的土壤都彼此分开。这说明不同秋浇年限处理对土壤细菌群落组成的影响具有显著差异性。土壤盐分、pH、微生物量碳与可溶性有机碳4个土壤环境因子变量覆盖了78.83%的土壤环境信息,这足以说明是这些因素的共同作用影响了不同秋浇年限土壤细菌群落的组成,将AUI1、AUI2、CK、AUI4和AUI3土壤区分开,但AUI4和CK、AUI1和AUI2处理的土壤较为接近。
通过对不同秋浇年限土壤细菌群落多样性与土壤环境关系冗余分析(图4-b)发现,土壤盐分(SC)和pH对细菌群落的丰富度(chao1、ACE)指数呈现较强的负相关性,说明土壤盐分和pH的改变可以显著影响土壤细菌菌群变化。4个理化因子总共解释了97.9%的群落变化,影响顺序依次为:土壤盐分>pH>微生物量碳>可溶性有机碳,其中土壤盐分解释了91.3%,土壤pH解释了4.9%,因此土壤盐分和pH是改变不同秋浇年限土壤细菌群落多样性的主控环境因子。
不同秋浇年限中优势细菌群落(门水平)与土壤化学性质的相关性分析(表3)表明,除了相对丰度较高的酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)、芽单胞菌门(Gemmatimonadetes)和浮霉菌门(Planctomycetes)与4个化学性质都不具有相关性外,其他优势细菌群落对土壤化学性质的响应不同,并且每个细菌群落与土壤pH均无显著相关性。其中,相对丰度靠前的变形菌门与土壤微生物量碳和可溶性有机碳均具有很强的负相关性(分别为-0.559**和-0.522*);放线菌门、奇古菌门和疣微菌门均与土壤盐分呈显著负相关(分别为-0.620*、-0.719*和-0.679**),与土壤微生物量碳和可溶性有机碳呈显著正相关;而绿弯菌门与土壤盐分呈显著正相关(为0.765**),与土壤微生物量碳和可溶性有机碳呈显著负相关(分别为-0.795**和-0.820**);厚壁菌门仅与土壤盐分呈显著负相关(为-0.559*)。这说明土壤盐分含量的增加不利于放线菌门、奇古菌门、疣微菌门和厚壁菌门细菌菌群的生长繁殖,有利于绿弯菌门细菌菌群的生长繁殖;微生物量碳和可溶性有机碳含量的提高不利于变形菌门、绿弯菌门细菌菌群的生长繁殖,有助于放线菌门、奇古菌门和疣微菌门细菌菌群的生长繁殖。
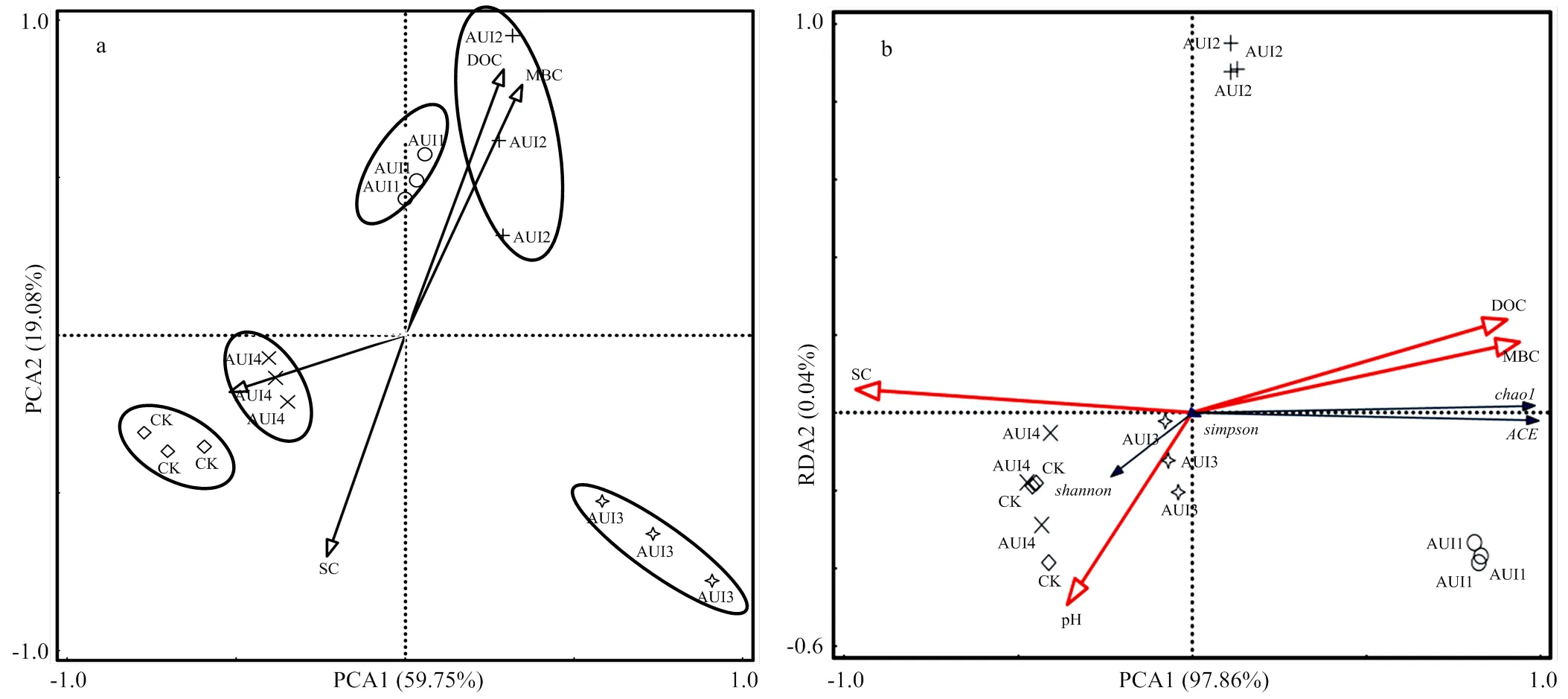
DOC:可溶性有机碳;MBC:微生物量碳;SC:盐分;CK:盐荒地;AUI1:一直秋浇;AUI2:隔2—3年秋浇;AUI3:3—4年不秋浇;AUI4:6—7年不秋浇

表3 细菌群落结构与土壤理化性质相关关系
*表示相关显著(<0.05),**表示相关极显著(<0.01) *and** indicate significance differences at 0.05 and 0.01 levels
3 讨论
3.1 不同秋浇年限对土壤理化性质的影响
有研究表明,秋浇可以充分改变盐分在土体中的空间分布,起到良好的淋盐、保墒作用,但由于该地区较小的排灌比导致秋浇仅能将耕作层中的盐分淋洗到深层土壤、地下水及盐荒地中[20],第二年经过冻融交替、整个作物生育期至秋季作物收获后盐分又返回地表[21]。而不秋浇地块由于缺乏大水压盐往往盐分分布在表层0—20 cm处,所以这使得该地区不秋浇农田的土壤盐分高于秋浇农田土壤[22],本研究中盐荒地(CK)与6—7年不秋浇(AUI4)处理在0—20 cm处秋浇盐分含量显著高于其他处理的结果也印证这一观点。本研究结果还表明,一直秋浇(AUI1)处理的土壤盐分含量随着土层的加深在逐渐减小,隔2—3年秋浇(AUI2)和3—4年不秋浇(AUI3)的土壤盐分含量随着土层的加深在逐渐增大。但一直秋浇(AUI1)和隔2—3年秋浇(AUI2)处理的土壤盐分含量在0—30 cm土层平均显著低于CK、AUI3和AUI4处理,从单纯节水角度考虑,隔2—3年秋浇是该地区淋洗耕层盐分以及减少灌溉优先考虑的措施。土壤pH值是反映盐碱土壤改良状况的重要指标,与土壤微生物的活性、有机质的分解、土壤养分的释放与固定等密切相关[23]。本研究发现隔2—3年秋浇(AUI2)处理的土壤pH值最低,且显著低于一直秋浇(AUI1)处理,并且随着不秋浇年限的增加,土壤pH值逐渐增高,同时6—7年不秋浇(AUI4)处理的pH值显著高于盐荒地(CK),由此可见,一直秋浇以及长时间不秋浇处理均对降低pH值效果不显著,太长时间不秋浇会造成pH值显著升高。综上所述,隔2—3年秋浇(AUI2)有利于降低0—30 cm土层盐分,从而达到淋洗盐分及降低土壤pH的作用。
土壤微生物量碳与可溶性有机碳碳含量与盐碱化程度有关[24],本研究所涉及的不同秋浇年限对活性有机碳的影响,归根结底是盐碱化程度不同造成的。有研究表明,随着盐渍化程度的加重土壤微生物碳和可溶性有机碳显著减少[15],本研究中结果也表明,一直秋浇(AUI1)和隔2—3年秋浇(AUI2)处理的微生物碳和可溶性有机碳显著高于3—4年不秋浇(AUI3)、6—7年不秋浇(AUI4)以及盐荒地(CK)。这反映了盐分胁迫对微生物量碳与可溶性碳具有抑制效应,与康健等[25]对不同植被类型对滨海盐碱地土壤有机碳库的影响的研究结果相似。所以一直秋浇(AUI1)和隔2—3年秋浇(AUI2)处理对提高土壤活性有机碳的效果最显著。
3.2 不同秋浇年限对土壤细菌群落多样性及组成的影响
不同秋浇年限对微生物的影响也是由于盐碱差异造成的。有研究表明,不同盐碱胁迫对土壤细菌群落多样性(Shannon和Simpson)指数和丰富度(Chao1和ACE)指数无显著影响[26],Andronov等[27]对哈萨克斯坦Akkol湖附近盐渍土壤细菌群落结构的研究中也表明细菌群落多样性(Shannon和Simpson)和丰富度(Chao1和ACE)指数不随盐度的增加而降低,这可能是因为细菌对盐渍化环境有很快的适应能力[26]。但同时也有研究表明,随着盐碱化程度的加剧,土壤细菌群落的(Shannon和Simpson)指数和丰富度(Chao1和ACE)指数都显著降低[14],孙佳杰等[9]的研究也指出土壤微生物数量随着盐碱化程度的增加而减少。而本研究发现,不同秋浇年限处理的土壤细菌多样性(Shannon和Simpson)指数无显著差异,一直秋浇(AUI1)和隔2—3年秋浇(AUI2)处理的土壤细菌丰富度(Chao1和ACE)指数显著高于其他处理,且随着不秋浇年限的增加,其丰富度(Chao1和ACE)指数逐渐减小,逐渐接近于盐荒地。可见一直秋浇(AUI1)和隔2—3年秋浇(AUI2)处理可以显著提高土壤细菌丰富度(Chao1和ACE)指数。
通过对不同秋浇年限处理的10大优势菌门分析(图3)发现,变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)和拟杆菌门(Bacteroidetes)是不同秋浇年限处理中相对丰度最高的3个菌门,占所有优势菌门的53%以上,这表明变形菌、酸杆菌和拟杆菌在盐碱土壤中的生长能力较强。变形菌门在盐碱土壤中相对丰度最高与大部分研究类似[28—29],这是由于变形菌门具有较好的耐盐性[30],以及其包括多种病原菌和固氮菌,可以利用有机物分解产生的氨气、甲烷等营养物质进行生长代谢活动[28],这与本研究中所表明的变形菌门在CK和AUI4处理中相对丰度最高的结果一致。而本研究中酸杆菌门和拟杆菌门相对丰度的提高可能与不同秋浇年限造成的盐碱程度有关。一些研究表明拟杆菌门细菌相对丰度与土壤盐度呈显著正相关,对高盐环境具有较强的抗性,是盐碱土中的优势种群,属于中度嗜盐菌[26]。而本研究也发现拟杆菌门在AUI2处理中的相对丰度最高,在CK、AUI1和AUI4处理中相对丰度居中,而在AUI3处理中的相对丰度最低,可见拟杆菌门细菌的生长是有一个适宜的盐分范围的,盐分过高或过低均不利于其生长繁殖。但就本研究而言,拟杆菌门最适宜在隔2—3年秋浇(AUI2)处理中繁殖。而酸杆菌门是嗜酸性细菌,碱性土壤环境不利于其生长,所以在碱性环境中酸杆菌门细菌丰度相对较低[31],但本研究结果却表明酸杆菌门细菌是仅次于变形菌门细菌的第二优势种群,这可能是因为酸杆菌门可以生长在土壤养分较缺的环境中[32]。
3.3 土壤化学性质与细菌群落多样性的冗余分析以及与群落组成的相关性分析
细菌群落多样性与土壤化学性质参数的冗余分析(图4)表明,土壤盐分对细菌群落多样性的影响最显著,这与丁新景等[33]的研究结果一致。而土壤pH、微生物量碳和可溶性有机碳也对细菌群落有一定的影响,其中土壤pH对细菌群落多样性指数(Simpson和Simpson)呈现较强的负相关性,土壤微生物量碳对细菌群落的丰富度指数(Chao1和ACE)呈现比较强的正相关性,其中土壤盐分解释了91.3%的群落变化,土壤pH解释了4.9%的群落变化,而土壤微生物量碳和可溶性有机碳总共解释了1.7%的群落变化,这说明土壤盐分和pH是引起土壤细菌群落多样性变化的重要因子。有研究发现拟杆菌门与土壤盐分呈显著正相关,浮霉菌门与土壤pH呈显著负相关,而变形菌门具有较好的耐盐性,是盐碱土壤中的优势种群[26]。本研究对优势细菌群落(门水平)与土壤化学性质的相关性分析(表3)表明,除了酸杆菌门、拟杆菌门和芽单胞菌门与土壤化学性质无显著相关性外,其他优势菌门与土壤化学因子都有一定的相关性。其中放线菌门、奇古菌门和疣微菌门与土壤盐分呈显著负相关关系,与土壤可溶性有机碳呈显著正相关关系;绿弯菌门与土壤盐分呈极显著正相关关系;变形菌门、绿弯菌门与土壤微生物量碳和可溶性有机碳呈显著负相关关系;这表明土壤盐分可能是影响放线菌门、奇古菌门、绿弯菌门、疣微菌门和厚壁菌门丰度的重要因子;微生物量碳和可溶性有机碳可能是影响变形菌门、放线菌门、奇古菌门、绿弯菌门和疣微菌门丰度的重要因子;而土壤pH对任何一种菌门的丰度都无显著影响。因此,不同秋浇年限处理可能是通过影响土壤盐分、微生物量碳和可溶性有机碳这些化学性质与土壤细菌菌群的相互作用来改变其群落组成的。
4 结论
4.1 隔2—3年秋浇可以促进耕层盐分淋洗,显著地降低0—40 cm土层的pH值,在一定程度上调节土壤酸碱度,同时提高0—40 cm土层微生物量碳和可溶性有机碳含量。
4.2 不同秋浇年限处理的细菌门水平相对含量及组成发生了一定的变化,其中隔2—3年秋浇可显著增加拟杆菌门(Bacteroidetes)、芽单胞菌门(Gemmatimonadetes)、浮霉菌门(Planctomycetes)细菌的相对丰度。
4.3 变形菌门与土壤微生物量碳和可溶性有机碳呈显著负相关;放线菌门、奇古菌门和疣微菌门与土壤盐分呈显著负相关,与微生物量碳和可溶性有机碳呈显著正相关;绿弯菌门与土壤盐分呈极显著正相关,与微生物量碳和可溶性有机碳呈极显著负相关;而厚壁菌门仅与土壤盐分呈显著负相关。
4.4 不同秋浇年限明显改变了土壤细菌多样性与群落组成,土壤盐分、pH、微生物量碳和可溶性有机碳是改变土壤细菌群落变化的重要环境因子,其中土壤盐分和pH又是提高土壤细菌群落丰富度的主控环境因子。
综上所述,隔2—3年秋浇(AUI2)处理不仅可以有效降低耕层土壤盐分并能使0—40 cm土层微生物量碳和可溶性有机碳含量增加,而且还可提高土壤优势菌群中拟杆菌门的相对丰度。综合考虑节水、改土等效果,隔2—3年秋浇(AUI2)是河套地区兼顾盐碱土壤改良与节水的优化措施。
[1] 冯保清, 崔静, 吴迪, 管孝艳, 王少丽. 浅谈西北灌区耕地盐碱化成因及对策. 中国水利, 2019(9): 43-46.
FENG B Q, CUI J, WU D, GUAN X Y, WANG S L. Preliminary studies on causes of salinization and alkalinization in irrigation districts of northwest China and countermeasures., 2019(9): 43-46. (in Chinese)
[2] 郭姝姝, 阮本清, 管孝艳, 王少丽, 李云鹏. 内蒙古河套灌区近30年盐碱化时空演变及驱动因素分析. 中国农村水利水电, 2016(9): 159-162, 167.
GUO S S, RUAN B Q, GUAN X Y, WANG S L, LI Y P. Analysis on spatial-temporal evolution of soil salinity and its driving factors in Hetao Irrigation District during recent 30 Years., 2016(9): 159-162, 167. (in Chinese)
[3] 杨劲松, 姚荣江, 王相平, 谢文萍. 河套平原盐碱地生态治理和生态产业发展模式. 生态学报, 2016, 36(22): 7059-7063.
YANG J S, YAO R J, WANG X P, XIE W P. Research on ecological management and ecological industry development model of saline- alkali land in the Hetao Plain, China., 2016, 36(22): 7059-7063. (in Chinese)
[4] 彭振阳, 伍靖伟, 黄介生. 内蒙古河套灌区局部秋浇条件下农田水盐运动特征分析. 水利学报, 2016, 47(1): 110-118.
PENG Z Y, WU J W, HUANG J S. Water and salt movement under partial irrigation in Hetao Irrigation District, Inner Mongolia., 2016, 47(1): 110-118. (in Chinese)
[5] 杨丽清, 刘燕凤, 刘坤. 浅谈义长灌域春灌面积增大的成因. 内蒙古水利, 2017(4): 66-67.
YANG L Q, LIU Y F, LIU K. Talking about the causes of the increase of spring irrigation area in Yichang irrigation area., 2017(4): 66-67. (in Chinese)
[6] 刘媛超. 内蒙古河套灌区秋浇灌水作用及秋浇节水潜力浅析. 内蒙古水利, 2017(5): 51-52.
LIU Y C. Analysis of autumn irrigation and irrigation water potential and water saving potential of autumn irrigation in Hetao Irrigation District of Inner Mongolia., 2017(5): 51-52. (in Chinese)
[7] 李凤霞, 王学琴, 郭永忠, 许兴, 杨建国, 季艳清. 宁夏不同类型盐渍化土壤微生物区系及多样性. 水土保持学报, 2011, 25(5): 107-111.
LI F X, WANG X Q, GUO Y Z, XU X, YANG J G, JI Y Q. Microbial flora and diversity in different types of saline-alkali soil in Ningxia., 2011, 25(5): 107-111. (in Chinese)
[8] 牛世全, 龙洋, 李海云, 达文燕, 胡山, 李渭娟, 朱学泰, 孔维宝. 应用IlluminaMiSeq高通量测序技术分析河西走廊地区盐碱土壤微生物多样性. 微生物学通报, 2017, 44(9): 2067-2078.
NIU S Q, LONG Y, LI H Y, DA W Y, HU S, LI W J, ZHU X Q, KONG W B. Microbial diversity in saline alkali soil from Hexi Corridor analyzed by Illumina MiSeq high-throughput sequencing system., 2017, 44(9): 2067-2078. (in Chinese)
[9] 孙佳杰, 尹建道, 解玉红, 杨永利, 舒晓武, 刘保东. 天津滨海盐碱土壤微生物生态特性研究. 南京林业大学学报(自然科学版), 2010, 34(3): 57-61.
SUN J J, YIN J D, XIE Y H, YANG Y L, SHU X W, LIU B D. Study on microbial ecological characteristics of coastal saline-alkali soil in Tianjin., 2010, 34(3): 57-61. (in Chinese)
[10] 樊金萍, 张建丽, 王婧, 逄焕成, 李玉义, 李二珍, 靳存旺. 节水灌溉对盐渍土盐分调控与土壤微生物区系的影响. 土壤学报, 2012, 49(4): 835-840.
FAN J P, ZHANG J L, WANG J, PANG H C, LI Y Y, LI E Z, JIN C W. Effects of water saving irrigation on salinity control and soil microbial., 2012, 49(4): 835-840. (in Chinese)
[11] 张云, 张相柱. 适宜河套灌区的四种田间节水灌溉技术. 内蒙古水利, 2018(10): 33-34.
ZHANG Y, ZHANG X Z. Four field water-saving irrigation techniques suitable for Hetao Irrigation District., 2018(10): 33-34. (in Chinese)
[12] 张晓红, 侯利, 刘永河, 郝爱枝. 河套灌区坝楞试区葵花灌溉前后水盐变化研究. 内蒙古水利, 2018(6): 8-10.
ZHANG X H, HOU L, LIU Y H, HAO A Z.Water and salt changes before and after sunflower irrigation in the dam test area of Hetao irrigation district., 2018(6): 8-10. (in Chinese)
[13] 王美荣, 史海滨, 闫建文, 李仙岳, 窦旭, 丁宗江. 水氮调控对休耕期土壤养分迁移及翌年土壤肥力的影响. 水土保持学报, 2019, 33(2): 146-154.
WANG M R, SHI H B, YAN J W, LI X Y, DOU X, DING Z J. Effects of water and nitrogen regulation on soil nutrient transfer during fallow period and soil fertility in the following year., 2019, 33(2): 146-154. (in Chinese)
[14] 李新, 焦燕, 代钢, 杨铭德, 温慧洋. 内蒙古河套灌区不同盐碱程度的土壤细菌群落多样性. 中国环境科学, 2016, 36(1): 249-260.
LI X, JIAO Y, DAI G, YANG M D, WEN H Y. Soil bacterial community diversity under different degrees of saline-alkaline in the Hetao area of Inner Mongolia.2016,36(1): 249-260. (in Chinese)
[15] 景宇鹏, 李跃进, 年佳乐, 宋瑶. 土默川平原不同盐渍化程度土壤微生物生态特征研究. 生态环境学报, 2013, 22(7): 1153-1159.
JING Y P, LI Y J, NIAN J L, SONG Y. Study on ecological characteristics of microbes under different soil salinization degrees in Tumochuan plain., 2013, 22(7): 1153-1159. (in Chinese)
[16] VANCE E D, BROOKES P C, JENKLMON D S. An extraction method for measuring soil microbial biomass C., 1987, 19(6): 703-707.
[17] BOKULICH N A, SUBRAMANIAN S, FAITH J J, GEVERS D, GORDON J I, KNIGHT R. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing., 2012, 10(1): 57-59.
[18] QUAST C, PRUESSE E, YILMAZ P, GERKEN J, SCHWEER T, YARZA P. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools., 2013, 41(D1): D590- D596.
[19] SCHLOSS P D, WESTCOTT S L, RYABIN T, HALL J R, HARTMANN M, HOLLISTER E B. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities., 2009, 75(23): 7537-7541.
[20] 刘显泽, 岳卫峰, 倪宝锋, 陈爱萍. 秋浇对内蒙古义长灌域地下水动态的影响. 北京师范大学学报(自然科学版), 2016, 52(3): 380-386.
LIU X Z, YUE W F, NI B F, CHEN A P. Impact of autumn irrigation on groundwater dynamics in Yichang Irrigation Sub-district, Inner Mongolia., 2016, 52(3): 380-386. (in Chinese)
[21] 倪东宁, 李瑞平, 史海滨, 苗庆丰, 梁建财. 秋灌对冻融期土壤水盐热时空变化规律影响及灌水效果评价. 干旱地区农业研究, 2015, 33(4): 141-145.
NI D N, LI R P, SHI H B, MIAO Q F, LAING J C. Temporal and spatial variation of soil water-salt-heat during freezing and thawing period and the irrigation effect under the autumn irrigation., 2015, 33(4): 141-145. (in Chinese)
[22] 王文达, 兰敬芳. 内蒙古河套灌区耕地与盐荒地盐分变化规律研究. 现代农业科技, 2017(3): 156-158.
WANG W D, LAN J F. Salinity change rule of cultivated land and salt wasteland in Hetao irrigation district in Inner Mongolia., 2017(3): 156-158. (in Chinese)
[23] 王军, 顿耀龙, 郭义强, 窦森. 松嫩平原西部土地整理对盐渍化土壤的改良效果. 农业工程学报, 2014, 30(18): 266-275.
WANG J, DUN Y L, GUO Y Q, DOU S. Effects of land consolidation on improvement of salinity soil in Western Songnen Plain., 2014, 30(18): 266-275. (in Chinese)
[24] 朱阳春, 张振华, 赵学勇, 连杰, 童非, 张娜. 河套灌区土壤有机碳和总碳的空间异质性及相关性分析. 江苏农业学报, 2017, 33(6): 1294-1300.
ZHU Y C, ZHANG Z H, ZHAO X Y, LIAN J, TONG F, ZHANG N. Spatial heterogeneity and relationship between soil total organic carbon and total carbon in the Hetao irrigation district., 2017, 33(6):1294-1300. (in Chinese)
[25] 康健, 孟宪法, 许妍妍, 栾婧, 隆小华, 刘兆普. 不同植被类型对滨海盐碱土壤有机碳库的影响. 土壤, 2012, 44(2): 260-266.
KANG J, MENG X F, XU Y Y, LUAN J, LONG X H, LIU Z P.Effects of different vegetation types on soil organic carbon pool in coastal saline-alkali soils of Jiangsu Province., 2012, 44(2): 260-266. (in Chinese)
[26] 张慧敏, 郭慧娟, 侯振安. 不同盐碱胁迫对土壤细菌群落结构的影响. 新疆农业科学, 2018, 55(6): 1074-1084.
ZHANG H M, GUO H J, HOU Z A. Effects of saline and alkaline stress on soil bacterial community structure., 2018, 55(6): 1074-1084. (in Chinese)
[27] ANDRONOV E E, PETROVA S N, PINAEV A G. Analysis of the structure of microbial community in soils with different degrees of salinization using T-RFLP and real-time PCR techniques., 2012, 45(2): 147-156.
[28] NULL N. Correction: salinity and bacterial diversity: to what extent does the concentration of salt affect the bacterial community in a saline soil?, 2014, 9(11): e114658.
[29] CAMPBELL B J, KIRCHMAN D L. Bacterial diversity, community structure and potential growth rates along an estuarine salinity gradient., 2013, 7(1): 210-220.
[30] ZHENG W, XUE D, LI X, DENG Y, RUI J, FENG K. The responses and adaptations of microbial communities to salinity in farmland soils: a molecular ecological network analysis., 2017(120): 239-246.
[31] GRIFFITHS B S, PHILIPPOT L. Insights into the resistance and resilience of the soil microbial community., 2013, 37(2): 112-129.
[32] 丁新景, 黄雅丽, 敬如岩, 马风云, 安然, 田琪, 陈博杰. 基于高通量测序的黄河三角洲4种人工林土壤细菌结构及多样性研究. 生态学报, 2018, 38(16): 5857-5864.
DING X J, HUANG Y L, JING R Y, MA F Y, AN R, TIAN Q, CHEN B J. Soil bacterial structure and diversity of four artificial forests in the Yellow River Delta based on high-throughput sequencing., 2018, 38(16): 5857-5864. (in Chinese)
[33] 丁新景, 敬如岩, 黄雅丽, 陈博杰, 马风云. 黄河三角洲刺槐根际与非根际细菌结构及多样性. 土壤学报, 2017, 54(5): 1293-1302.
DING X J, JING R Y, HUANG Y L, CHEN B J, MA F Y. Bacterial structure and diversity of rhizosphere and bulk soil offorests in Yellow River Delta., 2017, 54(5): 1293-1302. (in Chinese)
Effects of the Different Autumn Irrigation Years on Soil Bacterial Community in Hetao Irrigation District
ZHANG XiaoLi1, ZHANG HongYuan1, LU Chuang1, PANG HuanCheng1, JIN CunWang2, GAO Xi3, CHENG AiPing3, LI YuYi1
(1Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081;2Wuyuan County Agricultural and Animal Husbandry Technology Promotion Center, Wuyuan 015100, Inner Mongolia;3Yihe Canal Center of Yichang Experimental Station of Inner Mongolia Hetao Irrigation District, Wuyuan 015100, Inner Mongolia)
【】Considering the phenomenon that farmers voluntarily gave up the water irrigation in autumn, We investigated the effects of different autumn irrigation years on soil bacterial community composition in Inner Mongolia Hetao irrigation district, so as to provide corresponding scientific support for the improvement of saline-alkali land and reformation of autumn irrigation system.【】By selecting five types of typical plots, including wasteland (CK), always irrigation in autumn (AUI1), 2-3 years with autumn irrigation (AUI2), 3-4 years without autumn irrigation (AUI3) and 6-7 years without autumn irrigation (AUI4), the characteristics of soil bacteria in different autumn irrigation years were analyzed by high-throughput sequencing (Illumina HiSeq), and redundantly and correlation analysis of the soil chemical properties with bacterial communities and with community composition were followed.】The results showed that the salt content of AUI1 and AUI2 treatments were significantly reduced in 0-30 cm soil layer, compared with CK, AUI3 and AUI4, which were reduced by 128.82% and 29.04%, 108.76% and 17.72%, 108.44% and 17.55%, respectively. The salt content of AUI1 treatment was significantly reduced than that of other treatments in 30-40cm soil layer; the pH of AUI2 treatment was the lowest in each soil layer, and compared with CK, AUI1, AUI3 and AUI4 treatments, which reduced by 0.28, 0.32, 0.16 and 0.88 units, respectively (<0.05). Compared with CK, AUI1, AUI3 and AUI4 treatments, the soil microbial biomass carbon content of AUI2 treatment were increased by 252.89%, 148.59%, 58.10% and 60.10%, respectively, and the soluble organic carbon content increased by 48.41%, 29.42%, 6.01% and 4.27%, respectively (<0.05). The abundance index (ACE and Chao1) of AUI1 and AUI2 treatments were significantly higher than CK, AUI3 and AUI4 treatments (<0.05). Proteobacteria, Acidobacteria and Bacteroidetes were the three dominant bacteria that were treated in different autumn years, accounting for 53.93% of all bacteria, while AUI2 and AUI3 treatment are beneficial to increase the Bacteroidetes and Acidobacteria relative abundance (<0.05). Correlation analysis showed that Proteobacteria and Chloroflexi were significantly negatively correlated with soil microbial biomass carbon and soluble organic carbon, and the correlation coefficient with microbial biomass carbon were (=-0.559** and -0.522*), the correlation coefficients with soluble organic carbon were (=-0.795** and -0.820**), respectively; Actinobacteria, Thaumarchaeota, Firmicutes and Verrucomicrobia were significantly negatively correlated with soil salinity and significantly positively correlated with soil microbial biomass carbon and dissolved organic carbon. Factor analysis showed that soil salinity, pH, microbial biomass and soluble organic carbon were main environmental factors on the soil bacterial community structure, total explaining 97% of the community changes. The order of contribution rate was soil SC>pH>MBC>SOC.【】Comprehensive consideration, the 2-3 years with autumn irrigation (AUI2) treatment could not only effectively reduce the soil salinity and significantly increase the microbial biomass carbon and soluble organic carbon content in 0-40cm soil layer, but also improve the relative abundance of the dominant bacteria. In summary, the autumn irrigation (AUI2) was an optimization measure for the improvement of saline soil and water conservation in Hetao area.
autumn irrigation; soil bacterial; community structure; bacterial diversity; saline-alkali soil; IlluminaMiSeq; Inner Mongolia Hetao Irrigation District
10.3864/j.issn.0578-1752.2019.19.009

2019-03-25;
2019-06-10
国家自然科学基金(31871584,31471455)、国家重点研发计划项目(2016YFC0501302)
张晓丽,E-mail:1695952120@qq.com。
李玉义,E-mail:liyuyi@caas.cn
(责任编辑 李云霞)
