陇中旱作区不同轮作方式对土壤碳、氮含量及酶活性的影响特征
2019-09-25赵思腾师尚礼陈建纲
赵思腾, 师尚礼, 陈建纲, 陈 鑫, 何 佳
(甘肃农业大学草业学院,草业生态系统教育部重点实验室, 甘肃 兰州 730070)
陇中旱作区是我国主要的生态脆弱区,也是典型的丘陵旱作农业区,该区旱灾频繁,水土流失严重,地形破碎,且长期以马铃薯、小麦、玉米连作为主的规模化农业生产模式,自然资源过度利用,致使当地土壤养分耗竭,土壤质量退化[1]。因当地追求种植业规模化效益和粮食增产的需要,大量施用化学肥料不仅导致经济成本的增加,同时给当地脆弱的生态环境加重了压力,培肥地力构建健康的可持续发展农业生态系统成为当下面临的严峻问题。
土壤有机碳是评价土壤质量的一个重要指标[2-3],其储量大小由土壤中植物残渣降解及土壤微生物活动等决定,土壤有机碳可以为微生物的代谢提供能量和底物,在维持土壤肥力的过程中发挥重要作用[4-5],土壤微生物同样为土壤有机碳及土壤养分的循环转换提供动力,两者关系密切[6]。其中,土壤微生物碳和微生物氮常被用作评价土壤养分和土壤肥力的重要指标[7-8]。全氮作为一种大量且必要的营养元素,对土壤生态系统物质循环起着重要作用[9]。土壤酶是土壤的组成成分之一,其活性的强弱反映土壤中物质代谢的旺盛程度,是评价土壤质量的重要指标[10-11]。不同的轮作模式对土壤养分状况有不同程度的影响,虎德钰等[12]发现,有玉米和高粱参与的轮作模式其土壤酶活性比连作和有马铃薯、谷子参与的轮作模式高;赵靖静等[13]发现,连续种植10 a苜蓿轮作粮食作物后土壤总有机碳含量下降;陈丹梅等[14]发现,烤烟与不同作物轮作后提高了土壤总有机碳、微生物碳和微生物氮含量,土壤脱氢酶活性增强;谢泽宇等[15]通过对苜蓿与不同作物轮作模式土壤进行比较试验,研究发现苜蓿-玉米轮作模式土壤微生物氮含量提升明显。由此可见,因气候、作物、种植模式、土壤、施肥等因素对土壤碳、氮和土壤酶活性的影响十分复杂。玉米为陇中旱作区重要的规模化种植作物之一,在国家“粮改饲”调结构中也扮演着重要角色,高产优质,既能收获粮食,又能全株青贮作饲草,适宜于作为调整种植结构和“以种带养”的作物[16-17]。为此,本研究选择定西市安定区阎家湾村开展长期定位试验,研究了不同玉米轮作种植模式对土壤碳、氮和土壤酶活性的变化,并探究土壤碳和氮、土壤酶活性之间的交互作用,为陇中旱作区筛选最优玉米轮作模式及维持健康可持续发展的土壤生态环境提供科学有效的理论依据。
1 研究区概况与研究方法
1.1 研究区概况
试验地设在陇中黄土高原丘陵沟壑区的定西市安定区团结镇阎家湾村(35°23′ N,104°33′ E,海拔2 100 m)。该区年均太阳辐射592.9 KJ·cm-2,日照时数2 476.6 h,年均气温6.4℃,≥0℃年积温2 933.5℃,≥10℃年积温2 239.1℃;无霜期140 d。年平均降水390.9 mm,年蒸发量1 531 mm,干燥度2.53,降雨量为365 mm,属于典型的雨养旱作农业区。土壤为典型的黄绵土,黄土层较薄,土质绵软,土壤肥力均匀,贮水性能良好。试验初期0~40 cm土壤全氮含量平均为1.06 g·kg-1,有机质含量平均为15.41 g·kg-1,凋萎含水率约为7.30%,饱和含水率约为22%。
1.2 试验设计
以玉米、紫花苜蓿、小麦、马铃薯、大豆为轮作供试作物,以玉米-苜蓿、玉米-马铃薯、玉米-大豆和玉米-小麦4种轮作模式作为处理,玉米连作模式作为对照,试验设3次重复,小区面积7 m×7 m,随机区组排列,各处理详见表1。所有模式均施尿素105 kg·hm-2,过磷酸钙105 kg·hm-2,所有肥料仅在播种前施入作基肥,生育期未追肥。

表1 试验处理描述Table 1 Description of treatments
1.3 土壤样品采集与测定方法
于2018年10月用内径10 cm的土钻在各样地内随机采集3钻土样(2层,每层20 cm),将土样带回实验室,将混匀分好的土样装入自封袋。于实验室去掉植物根系、石块等,取部分土样放入4℃冰箱保存用以测定土壤酶活性以及土壤微生物碳和氮,剩余部分置于室内干燥通风处自然阴干,用于有机碳与全氮指标的测定。
土壤总有机碳(Total-organic carbon,TOC)含量的测定采用重铬酸钾外加热法[18];全氮(Total-nitrogen,TN)含量的测定采用凯氏定氮法[18]。土壤微生物碳(Microbial-biomass carbon,MBC)含量的测定采用氯仿熏蒸—硫酸钾浸提法[19];土壤微生物氮(Microbial-biomass nitrogen,MBN)含量的测定采用氯仿熏蒸浸提法[19]。蔗糖酶(Sucrase-glucose,SG)活性的测定采用3,5-二硝基水杨酸比色法[20];脲酶(Urease,UR)活性的测定采用靛酚比色法,以NH3-N mg·g-1(37℃,24 h)表示[20];过氧化氢酶(Catalase-gallicin,CG)活性的测定采用高锰酸钾滴定法[20]。
1.4 数据处理与分析
采用Microsoft Excel 2010软件进行数据整理,采用SPSS 19.0软件进行单因素方差分析(ANOVA)和最小显著差异(LSD)检验,Pearson双尾检验法进行相关性分析,Origin 9.1软件进行聚类分析和热图(heatmap)分析。
2 结果与分析
2.1 不同轮作模式对土壤碳含量的影响
由图1可知,不同轮作模式土壤总有机碳含量随土层深度的增加呈降低趋势,且不同土层间差异显著(P<0.05),变化范围为14.07%~16.38%。在0~20 cm土层,土壤总有机碳含量以C-A轮作模式最高,C-C最低,其中C-A,C-P,C-S,C-W有机碳含量显著高于C-C处理(P<0.05),分别高出27.52%,13.14%,21.93%和14.37%;在20~40 cm土层,土壤有机碳含量表现与0~20 cm土层基本一致,C-A,C-P,C-S,C-W与C-C差异显著(P<0.05),分别高出26.69%,14.16%,22.39%和16.75%。与试验开展前土壤相比,玉米连作模式土壤有机碳含量下降最为明显,说明玉米轮作模式更有利于土壤有机碳的积累,且不同轮作模式对土壤有机碳含量影响表现出差异性。
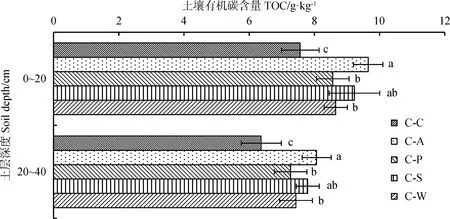
图1 不同轮作模式土层间土壤有机碳含量Fig.1 Soil organic carbon content in different soil layers
由图2可知,土壤微生物碳含量随土层深度的增加呈降低趋势,且不同土层间差异达显著水平(P<0.05),变化范围为16.52%~25.79%。在0~20 cm土层,微生物量碳含量表现为C-A最高,C-P最低,C-A,C-S,C-W显著高于C-C处理(P<0.05),分别高出18.11%,17.19%和6.75%;在20~40 cm土层,微生物碳含量表现与0~20 cm土层基本一致,C-A,C-S和C-W处理的微生物碳含量显著高于C-C处理(P<0.05),分别比C-C处理提高32.86%,29.36%和17.78%。
2.2 不同轮作模式对土壤氮含量的影响
由图3可知,土壤全氮含量随土层深度的增加呈降低趋势,不同土层间差异达显著水平(P<0.05),变化范围为4.35%~15.59%。在0~20 cm土层中,土壤全氮含量排序为C-S最高,C-C最低,C-S,C-A,C-W和C-P显著高于C-C处理(P<0.05),分别提高出57.61%,18.47%,58.69%和20.65%,说明轮作较连作模式有利于维持土壤全氮含量;在20~40 cm土层中,土壤全氮含量表现为C-A最高,C-C处理最低,相比于C-C处理,C-A,C-S提高显著(P<0.05),C-P,C-W无显著差异(P>0.05),C-A和C-S分别比C-C处理提高48.86%和40.91%。较初期土壤比,C-C模式土壤全氮含量有所降低,而C-A,C-S模式土壤全氮含量呈明显上升趋势。
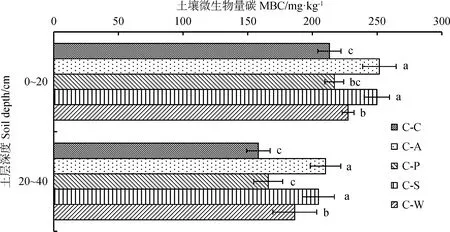
图2 不同轮作模式土层间土壤微生物碳含量Fig.2 Soil microbial biomass carbon content in different soil layers
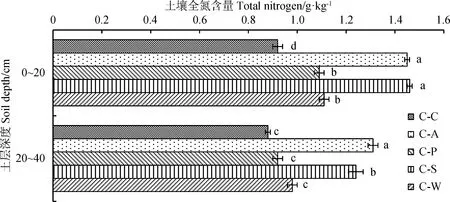
图3 不同轮作模式土层间土壤全氮含量Fig.3 Soil total nitrogen in different soil layers

图4 不同轮作模式土层间土壤微生物氮含量Fig.4 Soil microbial biomass nitrogen content in different soil layers
由图4可知,不同轮作模式下土壤微生物氮含量随土层深度的增加呈降低趋势,且不同土层间差异显著(P<0.05),变化范围为4.45%~13.74%。在0~20 cm土层中,土壤微生物氮含量的大小表现为C-S最高,C-C最低,其中C-A,C-P,C-S,C-W显著高于C-C处理(P<0.05),分别比C-C处理高出40.43%,20.68%,49.11%和18.31%;在20~40 cm土层中,土壤微生物氮含量表现与0~20 cm土层基本一致,C-A,C-P,C-S显著高于C-C处理(P<0.05),分别高出55.51%,22.08%,58.96%和30.28%。
2.3 不同轮作模式对土壤酶活性的影响
由表2可知,土壤蔗糖酶活性随着土层深度的增加呈降低趋势,且不同土层间差异显著(P<0.05),变化范围为3.53%~20.22%。在0~20 cm土层中,土壤蔗糖酶活性表现为C-S最高,C-C最低,C-A,C-S和C-W显著高于C-C处理(P<0.05),分别比C-C处理高出13.26%,17.11%和14.19%;在20~40 cm土层中,土壤蔗糖酶活性表现C-W最高,C-A,C-P,C-S和C-W显著高于C-C处理(P<0.05),分别高出30.81%,24.45%,31.54%和37.98%。说明轮作模式土壤熟化程度比连作高,有利于增加土壤中的营养物质。
土壤脲酶活性总体随土层深度的增加而降低(表2),且土层间差异达显著水平(P<0.05),变化范围为7.24%~19.78%。在0~20 cm土层中,土壤脲酶活性的表现为C-P最高,C-C最低,C-A,C-P,C-S土壤脲酶活性显著高于C-C处理(P<0.05),分别比C-C处理高出15.09%,26.42%和8.96%;20~40 cm土层脲酶活性表现为C-A最高,C-C处理其余处理差异不显著(P>0.05),说明不同轮作模式对下层土壤脲酶活性影响较小。
土壤过氧化氢酶活性随着土层深度的增加呈降低趋势(表2),土层间差异显著(P<0.05),变化范围为16.95%~29.14%。在0~20 cm土层,土壤过氧化氢酶活性大小表现为C-W最高,C-C最低,C-A,C-P和C-W显著高于C-C处理(P<0.05),分别提高10.89%,12.17%和13.46%;20~40 cm土层中,土壤过氧化氢酶活性表现与上层基本一致,与C-C处理相比,C-A和C-W过氧化氢酶活性显著高出17.09%和25.64%(P<0.05),说明轮作较连作更有利于增强土壤生物氧化过程。
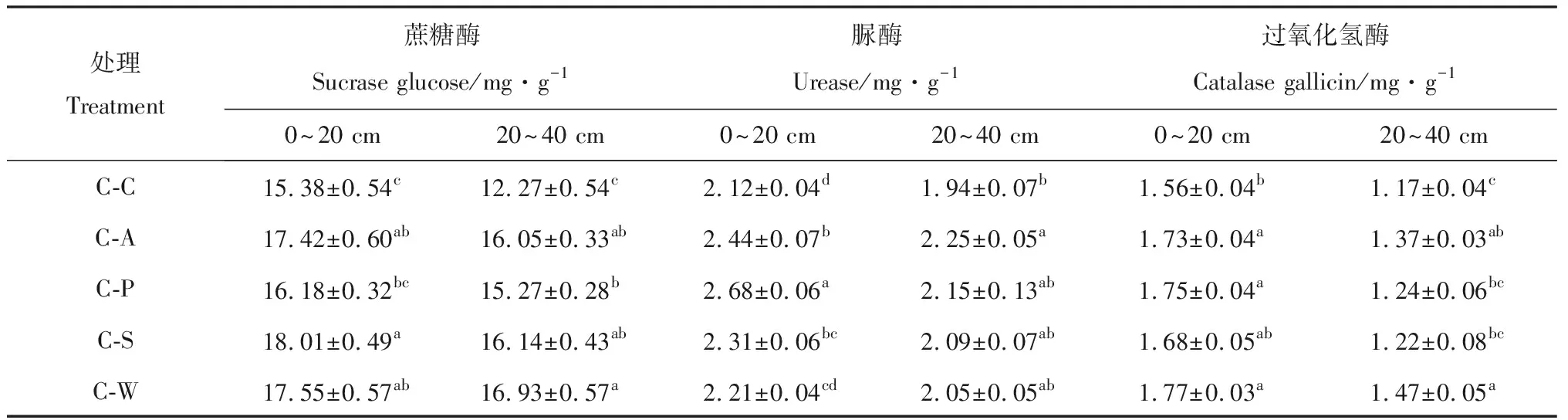
表2 不同轮作模式下不同土层土壤酶活性Table 2 Soil enzymes activities in different soil layers
注:同列不同小写字母表示处理间差异显著(P<0.05)
Note: Different lowercase letters within the same column for the same soil layer indicate significant difference among different treatments at the 0.05 level
2.4 土壤碳、氮含量与酶活性特征相关性分析
由图5可知,全氮含量与过氧化氢酶、脲酶、蔗糖酶活性呈显著正相关关系(P<0.05),其中全氮含量与蔗糖酶活性相关程度高,相关系数r=0.75;土壤有机碳含量与蔗糖酶活性呈显著正相关关系(P<0.05),相关程度较高,r=0.85;微生物碳含量与蔗糖酶活性呈显著正相关关系(P<0.05),且相关程度较高,r=0.81;微生物氮含量与蔗糖酶、脲酶、过氧化氢酶活性均呈显著正相关关系(P<0.05),其中微生物氮含量与蔗糖酶活性相关程度较高,r=0.71。
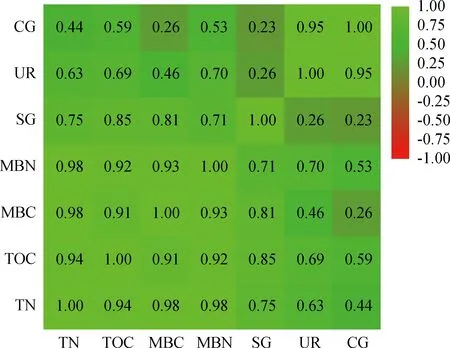
图5 土壤碳、氮与土壤微生物量碳、氮相关性分析热图Fig.5 Correlation analysis heatmap of soil carbon and nitrogen with soil microbial biomass carbon and nitrogen
2.5 不同轮作模式土壤碳、氮含量与酶活性特征聚类分析
由图6可知,根据不同指标间差异,利用origin 9.1软件对5种种植模式的土壤养分数据进行Z分数标准化,接近+1.5表示该轮作模式下养分含量较高,接近-1.5表示养分含量较低,后进行相似性聚类分析,将种植模式分为3组。C-A与C-S轮作模式的土壤养分状况聚类距离最近,距离值为0.96,聚集在组1中,C-W与C-C模式聚类距离值为3.08,聚集在组2;C-P模式接着与组1聚集在组3,聚类距离值为3.47。说明C-A与C-S轮作模式的土壤养分状况相似度最高,且土壤肥力表现较好。
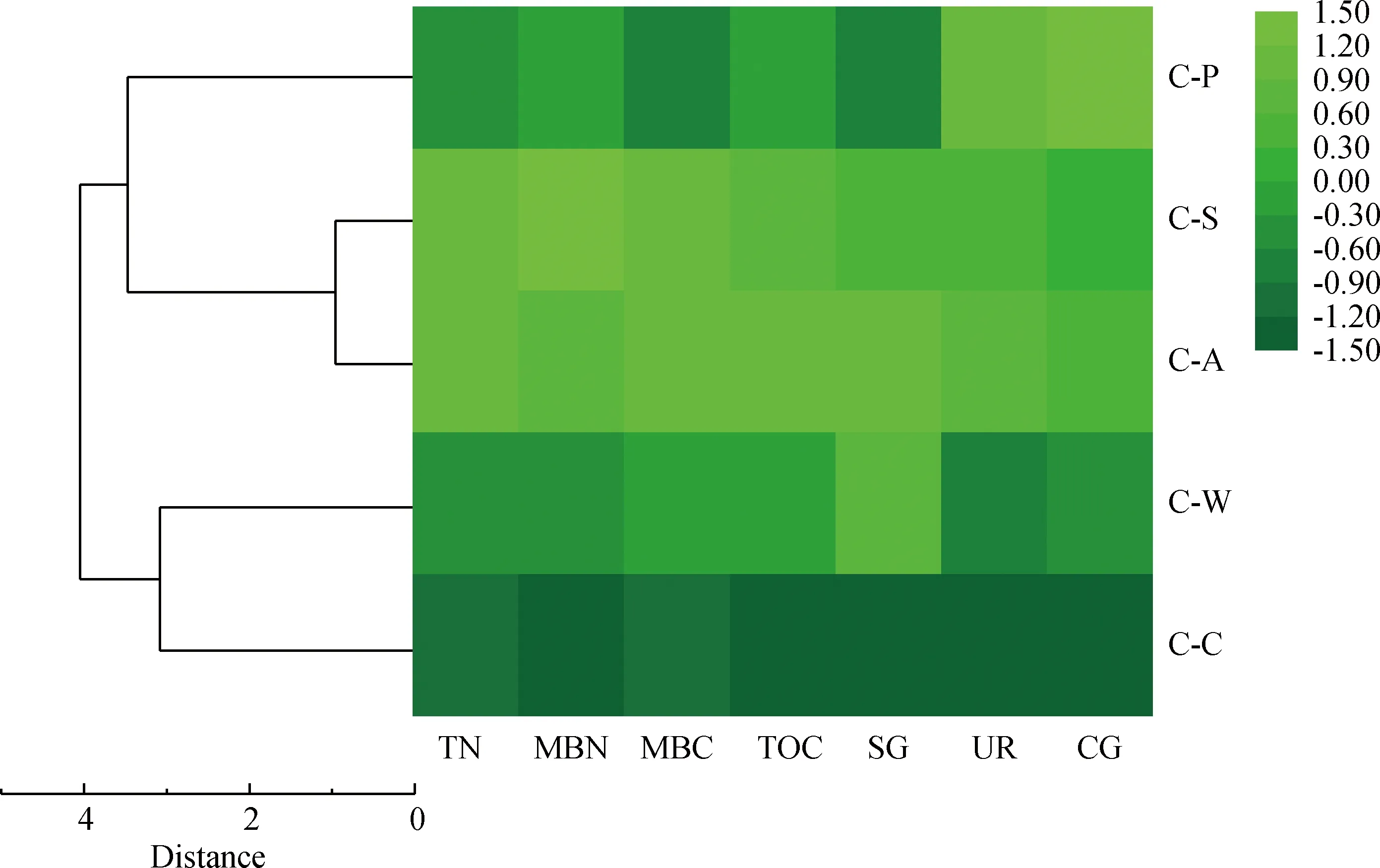
图6 土壤养分状况与5种种植模式聚类分析热图Fig.6 Cluster analysis heat map of 5 planting patterns and soil nutrient status
3 讨论
有研究[21-25]发现,包括苜蓿的轮作模式,苜蓿根部会形成大量根瘤菌,自身的固氮作用使土壤氮含量提高,且根系分泌的有机物和腐烂根系会增加土壤有机质含量;也有研究[13]表明,连作方式使土壤处于免耕状态,土壤的免扰动导致土壤有机碳含量保持在相对稳定的水平。本研究结果表明,与玉米-作物轮作模式相比,玉米-苜蓿轮作更有利于土壤有机碳的积累,改善土壤结构。其中玉米-马铃薯轮作模式中土壤有机碳含量最低,这可能是由于马铃薯产量较高,对土壤养分消耗较大,导致土壤有机碳含量下降。同时有研究[26]认为,轮作根系生物量较高的作物可以有效改良土壤氮素水平。本研究发现,土壤全氮含量随着种植年限的增长而降低,但玉米-苜蓿、玉米-大豆2种轮作处理的全氮和碱解氮含量呈现一定程度的提高,这可能是由于豆科作物能将空气中的氮固定在土壤中,提高了土壤的氮含量[27],因此玉米-苜蓿、玉米-大豆轮作模式下的土壤全氮含量处于较高水平。剡斌[28]通过对不同轮作模式下土壤氮素与酶活性进行比较试验,研究结果显示土壤脲酶活性较高时会加速土壤全氮的分解,导致土壤全氮含量降低。本研究结果与其一致,在玉米-马铃薯轮作模式下的土壤全氮含量较低,土壤脲酶活性较高。
不同的轮作处理显著影响土壤微生物量碳、氮含量。本研究中,玉米-作物轮作处理的微生物碳、氮含量高于玉米连作模式。有研究[29-31]表明,全氮与总有机碳的输入会提高土壤微生物碳、氮含量,这是由于碳源和氮源的供应改良了土壤理化环境,为微生物活动提供了丰富的营养物质,促进了微生物繁殖,使得土壤微生物碳、氮含量处于较高水平。但如果土壤碳、氮含量过高,土壤保持较高的速效养分浓度并不利于微生物的活动和繁殖。本研究结果显示,玉米-作物轮作处理的微生物碳、氮含量均高于玉米连作模式,而且玉米-苜蓿、玉米-大豆轮作模式的土壤全氮含量与有机碳含量较高,其微生物碳、氮含量也处于较高水平。但玉米连作模式的土壤有机碳含量相对较低,微生物碳、氮含量处于较低水平。与轮作相比,这可能是由于轮作倒茬的方式丰富了土壤的生物多样性,提高了土壤有机物质含量,且有机碳含量相对稳定,进而改善了土壤微生态环境,促进了微生物活动及腐殖质的形成,而且由于玉米根系较深,耗氮能力强等特点,使玉米连作的土壤微生物碳、氮含量降低。臧逸飞[32]研究发现,轮作模式相比于连作可以明显提高土壤微生物碳、氮含量,轮作倒茬的方式在土壤中存留了作物残渣及腐烂根系,为土壤微生物活动及繁殖提供了重要的营养物质,且不同作物自身对养分利用能力的差异性导致土壤结构变化,增强了土壤微生物活性,本研究结果与其一致。因此,通过合理轮作可以有效提高土壤微生物碳、氮含量。
土壤酶在参与土壤碳、氮循环过程中起着重要作用,其中包括土壤蔗糖酶、脲酶和过氧化氢酶等。不同种植模式下土壤酶活性呈不同程度变化,研究发现轮作的种植模式土壤酶活性高于连作模式[33-34]。本研究结果表明,玉米连作较玉米-作物轮作模式相比,土壤酶活性较弱,这可能是由于长年连作同种作物会抑制土壤过氧化氢酶活性,土壤中的过氧化氢得不到酶的分解产生堆积,从而引起连作障碍[35-36]。轮作模式可以丰富植物多样性,为土壤输入新的有机物。改良土壤理化性状不仅可以促进微生物活动,同时增强土壤酶活性,抑制土壤连作障碍的产生[37]。由相关性分析表明,不同种植模式的土壤碳和氮、土壤微生物碳和氮与土壤酶活性大多存在正相关关系,说明土壤酶活性对土壤环境变化较为敏感[12]。土壤微生物碳、氮含量与土壤蔗糖酶、脲酶、过氧化氢酶活性相关性较强,推测土壤酶与土壤微生物代谢密切相关,说明在土壤养分循环过程中,土壤碳、氮的转化会影响土壤微生物复杂的化学反应,从而引起土壤微生物碳、氮的改变,土壤酶参与了土壤碳和氮循环,三者协同在土壤质量改良过程中发挥着重要作用。但有研究得出土壤微生物碳、氮含量对土壤蔗糖酶、过氧化氢酶活性无影响,而本研究结果与其不一致,可能与作物间不同的生理特性、气候、土壤质量、种植年限等因素有关,需进一步探究其原因。根据指标间差异性,对5种种植模式的土壤养分状况进一步聚类分析,结果表明玉米-苜蓿、玉米-大豆轮作模式土壤养分状况相关性最强,可能由于轮作倒茬为土壤输入更多有机物质,缓解了连作作物对土壤肥力资源的过度利用,且有豆科作物参与的轮作模式更有利于改良土壤肥力。其具体原因还需进一步探究。
4 结论
在陇中旱作区采用玉米-玉米连作与玉米-作物轮作方式下,全氮含量以玉米-苜蓿、玉米-大豆轮作模式最高;总有机碳含量以玉米-苜蓿,玉米连作模式较高,玉米-作物轮作较低;微生物量碳、氮含量以玉米-苜蓿、玉米-马铃薯轮作模式最高;蔗糖酶、脲酶和过氧化氢酶活性以玉米-大豆、玉米-马铃薯和玉米-小麦轮作模式最高,玉米-玉米连作模式最低,轮作的种植方式较连作相比可以有效培肥地力。
相关性分析表明,土壤碳和氮、土壤微生物碳和氮含量与土壤酶活性之间存在正相关关系,聚类分析表明,玉米-苜蓿和玉米-大豆轮作模式土壤养分状况相似度最高,土壤肥力状况较好。本研究从土壤碳、氮含量和土壤酶活性的角度,阐明了有豆科作物参与的轮作模式改良土壤碳、氮含量及土壤酶活性效果最好,其中苜蓿为多年生作物,而大豆为一年生作物,轮作周期较短,因此,为发挥更优经济效益,针对陇中旱作区作物轮作制度和土壤生态环境状况,推荐农户采用玉米-大豆轮作模式。
