低温胁迫对结缕草抗氧化酶活性和脂肪酸含量的影响
2019-09-25李双铭徐庆国刘雪勇罗建章向佐湘胡龙兴
李双铭, 徐庆国, 杨 勇, 刘雪勇, 罗建章, 向佐湘, 胡龙兴*
(1. 湖南涉外经济学院, 湖南 长沙 410205; 2. 湖南农业大学农学院草业科学系, 湖南 长沙 410128; 3. 长沙学院, 湖南 长沙 410022)
结缕草(ZoysiaWilld) 属于禾本科结缕草属多年生草坪草种,广泛分布在热带、亚热带和暖温带地区。在我国,结缕草的分布北至东北的辽宁、吉林和华北的山东、河南,南至华南的广东、海南等地,具有抗旱、耐盐、耐践踏、低养护等诸多优良特性,广泛应用于城市园林绿化、运动场草坪、高尔夫球场、固土护坡和水土保持[1-2]。但结缕草为暖季型草坪草,其最适生长温为26~36℃,低于12℃时就会休眠,零下温度常造成冻害。因此,低温是影响结缕草地理分布、草坪质量和推广应用的主要限制因素[3]。
细胞膜是植物体细胞及细胞器与外界环境之间进行物质和能量交换的场所,同时也是细胞感应外界环境中各种逆境胁迫信号的细胞结构[4]。冷害和冻害常诱导植物体内活性氧(Reactive oxygen species,ROS)如O2-.、H2O2等显著积累而造成细胞膜系统受到破坏,而植物体内存在有效清除ROS的酶保护系统如超氧化物歧化酶(Super oxidase dismutase,SOD)、过氧化物酶(Peroxidase,POD)和过氧化氢酶(Catalase,CAT)等,可通过其含量或活性的增加来保护细胞膜免于ROS的伤害,使植物在一定程度上忍耐、减缓或抵御逆境伤害[5]。舒必超等研究表明,低温胁迫下狗牙根(Cynodondactylon)叶片抗氧化酶活性显著升高,其中耐寒性好的运动百幕大(Cynodondactylon×C.transvalensis‘Tifsport’)升高更显著[6]。已有的研究表明,细胞膜的结构及其稳定性与植物对低温胁迫的适应性密切相关,其耐寒性的大小主要体现在细胞膜的特性上,如膜脂的流动性和不饱和度,而脂肪酸是膜脂的主要成分[4]。脂肪酸含量的变化及其不饱和度与耐寒性的关系在多种植物中已有研究报道[7-10]。膜脂相变程度与不同脂肪酸的含量及比例关系密切,通常研究者都认为,植物的膜脂相变温度降低是由于不饱和脂肪酸含量的增高,从而增加了膜的流动性,使得植物的耐寒性增加;因此,膜不饱和脂肪酸指数(Index of unsaturated fatty acid,IUFA),即不饱和脂肪酸在总脂肪酸中的相对比值,可作为衡量植物抗冷性的重要生理指标[11]。
结缕草喜温暖湿润气候,具有较强的耐热性,抗寒性较差,但不同地理来源的结缕草由于其遗传背景的不同而耐寒性差异较大,研究不同地理来源结缕草的低温胁迫生理,对扩大其栽培区域具有重要的理论意义和实践应用价值。关于结缕草低温胁迫下的生理响应,前人大多只从冷害处理或冻害处理方面进行了研究探讨[12-18],但多个连续低温处理的研究国内外报道较少,而自然界草坪草在秋冬季常处于一个逐渐从最适生长温度、亚适温、冷害和冻害逐渐降温的过程。本试验以采自我国不同地理来源的结缕草为材料,研究在人工控制梯度降温条件下其叶片的抗性生理指标及脂肪酸含量的变化,以期阐明不同地理来源的结缕草在低温胁迫下抗寒性相关生理指标的动态变化及其差异,为结缕草抗寒性的鉴定提供参考依据以及为优异耐寒新品系的选育提供理论依据。
1 材料与方法
1.1 植物材料与培养
本研究根据前期研究结果[19],选择两个不同纬度来源的结缕草品种为研究对象,分别为Latitude-40(LT-40,来源于辽宁)和Latitude-22(LT-22,来源于广东)。
1.2 试验设计与处理
两个结缕草品种材料于2018年4月分别用草茎扩繁种植于湖南农业大学草业科学系教学科研试验基地。2018年7月,挖取致密的草皮块洗净其根部泥土,种植于装填了1.2 kg河沙和市售花土(体积比为1:1)混合基质的塑料盆中,盆的上口径为18 cm、下口径15 cm、高20 cm。每个基因型材料种植16盆(4个处理×4次重复),共计32盆,放置于湖南农业大学草业科学系教学科研试验基地玻璃温室中培养。根据土壤基质水分状况及时补充水分以保持水壤湿润,每3天修剪一次保持冠层高度约为5 cm,每周每盆浇1/2Hongland养液200 ml。
在温室中培养约45天冠层基本建成后,将各盆栽材料转入人工气候箱中按图1所示进行梯度降温处理:(1)适温处理。初始温度30℃/25℃(昼/夜)处理5 d;(2)亚适温处理。经初始处理材料再将气候箱温度降为18℃/12℃(昼/夜)处理5 d;(3)冷害处理。经初始和亚适温处理材料再在8℃/2℃(昼/夜)处理5 d;(4)冻害处理。经初始、亚适温和冷害处理材料再转入温度设为2℃/—4℃的生长箱中处理5 d。人工气候箱相对湿度设为65%~75%,12 h光照时间和12 000 lx的光照强度。各处理设4次重复(4盆),在各梯度降温处理结束时,每盆取植株功能叶片混匀,其中一部分叶样用于测定其电导率和叶绿素含量;另一部分鲜叶样在液氮速冻后保存于—80℃冰箱,用于抗氧化酶活性和脂肪酸含量的测定分析。

图1 温度处理及取样时间示意图Fig.1 The schematic diagram for the temperature treatment and sampling date
1.3 测定项目与方法
(1)细胞膜稳定性:用电导率(Electrolyte leakage,EL)表示,称取约0.1 g叶片,去离子水冲洗两次后剪成约1 cm的小段置于50 ml的离心管中,加去离子水10 ml后在摇床上以200 rpm振荡24 h后测电导率(E1),然后在高压灭菌锅中121℃灭菌15 min,待冷却至室温后测电导率(E2),相对电导率(%)=[(E2-E1)/E2]×100。
(2)叶绿素含量(Chl):用二甲基亚砜在室温避光条件下浸提48 h后,采用分光光度计法于波长663、645和470 nm下测定吸光值[20]。
(3)丙二醛(MDA)含量和抗氧化酶活性测定:称取约0.2 g新鲜叶片,用预冷的研钵研成粉后加3 ml预冷的酶提取液(50 mmol·L-1pH7.8的磷酸缓冲液)匀浆,匀浆经在4℃,12 000 rpm离心15 min,转移上清液至新的试管中用于MDA含量和抗氧化酶活性的测定。MDA采用硫代巴比妥酸法测定,SOD活性测定采用氮蓝四唑NBT光化还原法,POD活性采用愈创木酚法,CAT活性采用过氧化氢法测定,可溶性蛋白含量采用考玛斯亮蓝法测定,酶活性单位均以单位蛋白含量的酶活性表示[21]。
(4)脂肪酸提取与测定:参考Zhang[22]的方法进行。取0.2 g鲜样用液氮研磨后转至离心管中,加1 ml提取液(用甲醇配制的0.1 N盐酸,含5%的2,2-二甲氧基丙烷),同时添加十七碳酸(17:0)作为内标,在80℃水浴条件下提取90 min后冷确至室温,然后分别先后加入1 ml 0.5%的NaCl和1.5 ml己烷后涡旋30 s,在5 000 rpm离心5 min,将分层后的己烷层转移至新的试管中用于脂肪酸的测定。提取后的脂肪酸按Zhang[22]的方法,采用安捷伦5890A气相色谱仪进行测定分析,脂肪酸含量以单个脂肪酸占所测总脂肪酸含量的百分比表示,脂肪酸不饱和指数(IUFA)计算公式为:
IUFA=0×([16∶0]+[18∶0])+1×([16∶1]+[18∶1])+2×([16∶2]+[18∶2])+3×[18∶3]。
1.3 数据分析
采用SAS for Windows (9.0版本)软件进行试验数据统计,采用单因素方差分析,同温度处理下两个基因型间的差异采用t检测,同基因型不同温度间的差异采用Ducuan式多重比较检验(P<0.05).
2 结果与分析
2.1 低温对结缕草叶片细胞膜稳定性的影响
随着温度的降低,两种结缕草叶片电导率均呈上升趋势,但在亚适温下两个基因型叶片的电导率与适温对照相比均无显著性差异(图2);与适温对照相比,冷害和冻害条件下两个基因型叶片的电导率均显著升高,但LT-40显著低于LT-22,其中高纬度基因型LT-40的叶片电导率升高至适温对照的4.5和8.5倍,而低纬度基因型LT-22则分别升高至适温对照的6.1和9.2倍(图2)。
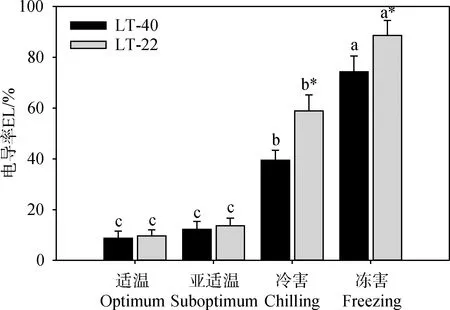
图2 低温胁迫对不同纬度来源结缕草叶片电导率的影响Fig.2 Effect of low temperature stress on electrolyte leakage in the leaf of zoysiagrass collected from different latitudes注:图柱上标注不相同小写字母表示同基因型不同温度处理间具有显著性差异,*表示同一温度理处下两个基因型材料间具有显著性差异(P<0.05),下同Note:Vertical bars marked with different lower-case letters indicate significant difference among temperature treatment in a given genotype, bars marked with * indicate significant difference between two genotypes in a given temperature treatment at the 0.05 level,the same as below
2.2 低温对结缕草叶绿素含量的影响
两种结缕草叶绿素含量均随着处理温度的降低而呈下降趋势(图3)。高纬度基因型LT-40在亚适温、冷害和冻害温度处理下的叶绿素含量分别下降至适温对照的89%,71%和46%,而低纬度基因型LT-22的叶绿素含量则分别下降至适温对照的75%,41%和27%;在冷害和冻害条件下,高纬度基因型LT-40的叶绿素含量显著高于低纬度基因型LT-22(图3)。

图3 低温胁迫对不同纬度来源结缕草叶绿素含量的影响Fig.3 Effect of low temperature stress on leaf chlorophyll content in zoysiagrass collected from different latitude
2.3 低温对结缕草叶片丙二醛含量的影响
两种结缕草叶片MDA含量均随着处理温度的下降而显著升高(图4)。高纬度基因型LT-40叶片的MDA含量在亚适温、冷害和冻害温度处理条件下分别升高至适温对照的1.3,1.8和2.5倍,而低纬度基因型LT-22则分别升高至适温对照的1.6,2.4和3.3倍;在冷害和冻害条件下,高纬度基因型结缕草LT-40叶片的MDA含量均显著低于低纬度基因型结缕草LT-22(图4)。
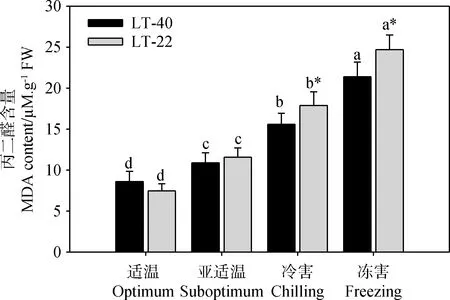
图4 低温胁迫对不同纬度来源结缕草叶片丙二醛含量的影响Fig.4 Effect of low temperature stress on MDA content in zoysiagrass collected from different latitude
2.4 低温对结缕草叶片抗氧化酶活性的影响
由表1可知随着处理温度的下降,两个不同纬度来源的结缕草叶片SOD活性均显著升高。在亚适温、冷害和冻害条件下,低纬度基因型LT-22的SOD活性分别升高至适温对照的1.28,1.8和2.3倍,而高纬度基因型LT-40的SOD活性则分别升高至适温对照的1.3,2.2和3.2倍。
两个不同纬度来源结缕草叶片POD活性均随着处理温度的降低而显著升高,在亚适温、冷害和冻害条件下,低纬度基因型LT-22的POD活性分别升高至适温对照的1.1,2.1和2.5倍,而高纬度基因型LT-40的POD活性则分别升高至适温对照的1.7,2.9和4.0倍。
随着处理温度的下降,两个不同纬度来源的结缕草叶片CAT活性均显著升高。在亚适温、冷害和冻害条件下,低纬度基因型LT-22的CAT活性分别升高至适温对照的1.28,1.8和2.3倍,而高纬度基因型LT-40的CAT活性则分别升高至适温对照的1.3,2.2和3.2倍。
在亚适温、冷害和冻害条件下,高纬度基因型LT-40的CAT活性显著高于低纬度基因型LT-22,而SOD和POD活性则仅在冷害和冻害下高于LT-22。
2.5 低温对结缕草叶片脂肪酸含量的影响
结缕草叶片的膜脂脂肪酸主要由棕榈酸(16∶0)、棕榈油酸(16∶1)、硬脂酸(18∶0)、油酸(18∶1)、亚油酸(18∶2)和亚麻酸(18∶3)6种脂肪酸组成。其中,饱和脂肪酸以棕榈酸为主,占总脂肪酸的13.5%~28.5%;而不饱和脂肪酸中以亚麻酸的相对含量最高,占总脂肪酸的43.6%~62.6%。
由表2可知,随着温度的降低,饱和脂肪酸中的棕榈酸在不同纬度来源的结缕草中均呈下降趋势,其中在高纬度基因型LT-40中下降更为显著,在亚适温、冷害和冻害条件下LT22的棕榈酸相对含量显著高于LT-40;随着温度的下降,饱和脂肪酸中的硬脂酸在低纬度基因型LT-22中无显著变化,但在高纬度基因型LT-40中则显著下降。
随着处理温度的下降,不饱和脂肪酸中的棕榈油酸、油酸和亚麻酸在低纬度基因型LT-22中均无显著变化;在高纬度基因型LT-40中,棕榈油酸、油酸的相对含量在冻害条件下显著低于适温,在亚适温和冷害条件下则无显著变化,而亚麻酸的相对含量在冷害和冻害条件下显著高于适温和亚适温处理,亚适温条件下的亚麻酸含量与适温相比无显著差异。
在适温、亚适温、冷害和冻害四个温度处理条件下,脂肪酸的不饱和指数在低纬度基因型LT-22中均无显著变,而在高纬度基因型LT-40中则随着处理温度的降低而呈上升趋势,`其中在冷害和冻害温度处理下的不饱和指数显著高于适温对照。
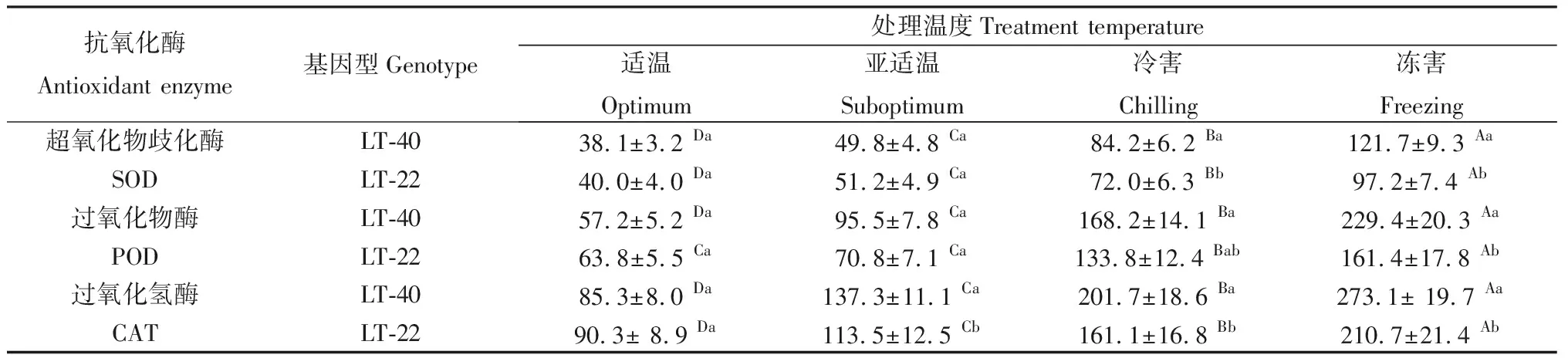
表1 低温胁迫对不同纬度来源结缕草叶片抗氧化酶活性的影响Table 1 Effect of low temperature stress on leaf antioxidant enzyme activity in zoysiagrass collected from different latitude / U.min-1.mg-1 protein
注:同行中不同大写字母表示同一基因型在不同温度处理下差异显著(Duncan’s多重比较,P<0.05),同列中不同小写字母表示同一温度处理下不同基因型之间差异显著(t检验,P<0.05)。下同
Note:Means in a row followed by different upper-case letters indicate significant difference among temperature treatments in a genotype,values followed by the different lower-case letters in a column indicate significant difference between two genotypes within a temperature treatment with t-test at the 0.05 level. The same as below
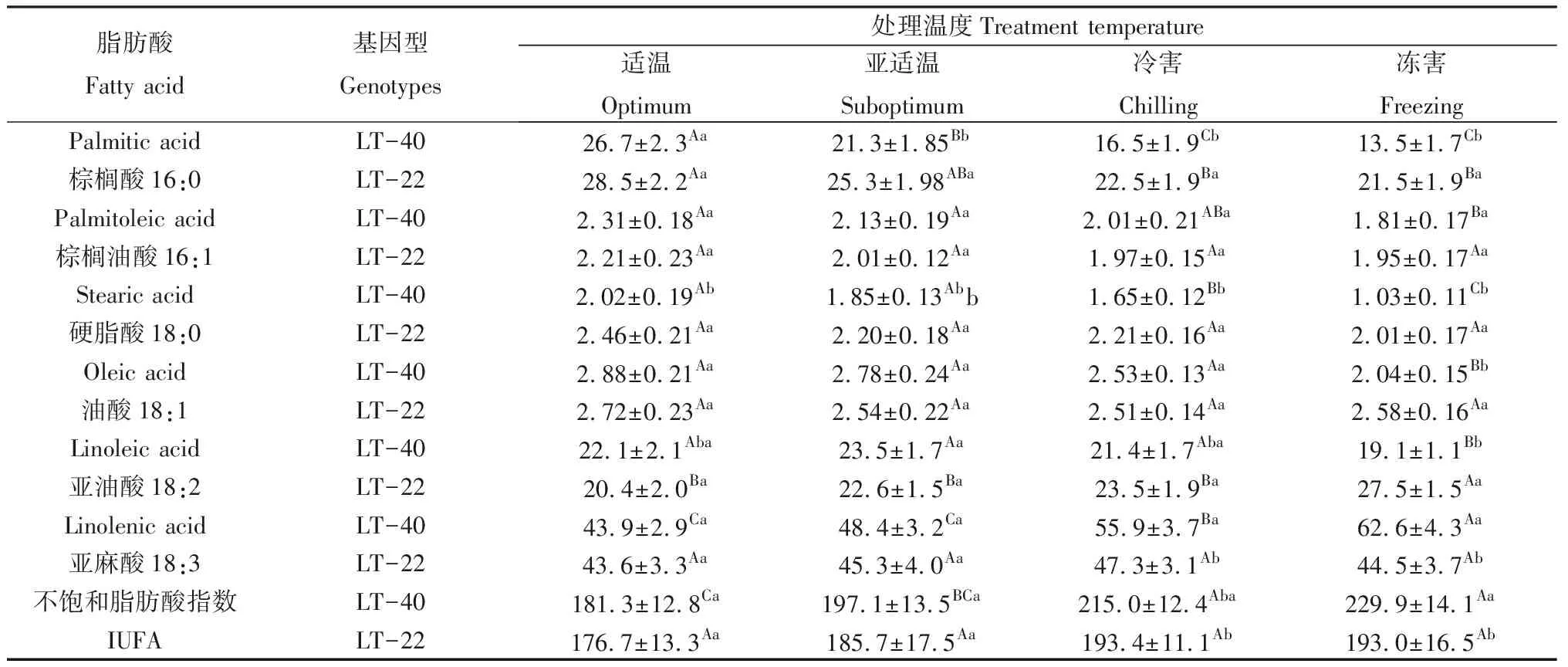
表2 低温对结缕草叶片脂肪酸含量的影响Table 2 Effect of low temperature on fatty acid content in leaves of zoysiagrass/%
3 讨论
在南北气候过渡带区域,冷害和冻害是暖季型草坪草常面临的环境逆境。当自然环境温度下降到最适生长温度以下时草坪草生长减缓,草坪质量下降,当温度降低到10℃以下时常发生冷害和冻害,但不同地理纬度来源的草坪草在自然选择和进化过程中逐渐形成了相应的适应机制而耐寒性差异较大[13-15]。娄燕宏等[23]通过在大田自然降温条件下对不同地理来源的狗牙根研究表明,高纬度来源的狗牙根具有较强的耐寒性。孙建明等[24]通过对在自然环境零下温度条件下不同地理来源日本结缕草的低温响应研究表明,来源于高纬度的日本结缕草具有较高的脯氨酸和可溶性糖含量而具有较好的耐低温能力。本试验通过人工控制模拟自然降温过程,研究了不同纬度来源的结缕草基因型对低温胁迫的响应,结果显示在低温处理下高纬度来源的基因型LT-40具有较高的细胞膜稳定性和较低的膜脂过氧化水平,叶绿素含量较高,表明纬度来源与结缕草的耐寒性密切相关,高纬度来源的结缕草具有较好的耐寒能力,这与娄燕宏等[23]和孙建明等[24]在大田自然环境条件下对草坪草的研究结果一致。
在正常生长条件下,植物体内ROS的产生与清除处于动态平衡而不会导致植物细胞膜受到伤害。但在低温逆境条件下,这种动态平衡常被打破而产生大量的ROS,造成细胞膜脂过氧化反应,导致质膜系统的损伤[25]。植物体为保护自身免受ROS的伤害,细胞内存在SOD,POD和CAT等抗氧化系统,使体内活性氧保持动态平衡,从而减轻或缓解逆境伤害[2]。在本研究中,两个不同纬度来源的结缕草在低温条件下,叶片的SOD,POD和CAT三种酶活性均显著上升,表明低温胁迫诱导了结缕草植株体内抗氧化防御系统的增强而提高了自身清除氧自由基的能力以减轻低温对细胞膜的伤害。其中,来源于高纬度的结缕草LT-40在低温条件下叶片的SOD,POD和CAT表现出更强的活性,表明高纬度来源的结缕草具有更好的耐低温能力,这与舒必超等[6]和孙宗玖等[26]在狗牙根以及付娟娟等[27]在披碱草中的研究结果相一致,在低温胁迫下抗寒性强的品种或材料具有较高的抗氧化酶活性。高纬度来源的结缕草具有更好的耐寒性,这可能是由于高纬度地区温度较低,长期的低温锻炼和适应使其具有相对发达的抗氧化防御系统及其对低温胁迫具有较高的响应水平有关[23-24]。但是,目前对植物在低温胁迫下抗氧化酶活性的研究结论仍存有差异。周艳虹等[28]在黄瓜幼苗中研究表明,在低温处理后SOD、POD活性上升,而彭金光等[29]在西瓜中研究认为,低温提高了耐性品种SOD,POD活性,而在敏感品种中则呈现先增后减的趋势。Back等[30]研究表明,短期的低温锻炼诱导了抗氧化酶活性的升高,有利于提高植株的抗寒性。玉米幼苗在短期低温胁迫(2~3 d)下SOD和POD活性上升,而CAT活性则下降,较长时间胁迫(7 d)下SOD,POD和CAT活性均显著下降[31]。这些在低温胁迫下抗氧化酶活性的变化差异可能与植物种类、耐性、胁迫温度和时间长短等不同有关。
植物体与外界环境相互作用时,生物膜是植物接受外界环境信息的受体,因而质膜的生理生化特性的变化是植物抵抗环境胁迫能力的直接反映[32]。本研究结果显示,随着温度的下降,结缕草叶片脂肪酸组分中饱和脂肪酸棕榈酸在两个基因型中呈下降趋势,不饱和脂肪酸中的亚麻酸在高纬度基因型LT-40中显著提高;低温胁迫下不饱和脂肪酸尤其是亚麻酸含量增加,这与前人在海滨雀稗[33]、冬青[34]的研究结果一致。不饱和脂肪酸含量与脂肪酸的不饱和指数直接相关,而不饱和指数直接影响质膜的流动性[32]。低温胁迫下结缕草叶片不饱和脂肪酸含量的升高有利于增加脂肪酸不饱和度和不饱和指数,从而维持膜的稳定性和流动性。脂肪酸不饱和指数在低纬度基因型LT-22中均无显著变化,而在高纬度基因型LT-40中则随着温度的降低而显著升高,表明不饱和脂肪酸的含量和不饱和指数与结缕草的耐寒性密切相关。张玮等[35]研究表明,耐寒性弱的竹子在低温下的不饱和脂肪酸含量及变化相对较低,且低温驯化对竹种耐寒性的累积提高是一个关键过程。低温胁迫可改变植物质膜脂肪酸的含量和组成比例,已有的研究大多数认为,低温驯化(锻炼)可以提高不饱和脂肪酸含量和不饱和脂肪酸指数,且与植物的耐寒性密切相关[7-10,36]。在本研究中,我们采用从适温到亚适温,再到冷害和冻害逐渐降温的过程,对结缕草的耐寒性起到了驯化的作用,尤其是对来源于高纬度的结缕草,在降温的过程逐渐通过提高不饱和脂肪酸含量和不饱和指数来提高细胞内束缚水含量而稳定细胞内水分,维持原生质结构,防止结冰脱水而造成机械损伤,从而提高了耐低温能力[32]。
4 结论
低温胁迫可造成结缕草叶片细胞膜稳定性丧失,电导率和MDA含量升高,叶片衰老而叶绿含量下降,进而影响结缕草的正常生长发育和草坪质量,同时造成细胞内自由基的平衡被破坏而诱导抗氧化酶活性的升高。来源于高纬度的结缕草在低温下具有较好的耐寒性,主要与其具有较高的不饱和脂肪酸亚麻酸含量和较高的脂肪酸不饱和指数有关,表现出更强的膜流动性和稳定性,以及抗氧化防御保护系统,因而其耐寒能力要显著高于低纬度来源的结缕草。
