布莱凯特黑牛A-FABP 基因表达规律及其肌内脂肪含量研究
2019-09-19刘瑞莉刘贤勋柏学进董雅娟
刘瑞莉,袁 玮,吴 磊,刘贤勋,柏学进,3,董雅娟,3*
(1.青岛农业大学动物胚胎工程中心,山东青岛 266109;2.山东省黑牛繁育工程技术研究中心,山东青岛 266109;3.山东布莱凯特黑牛科技股份有限公司,山东淄博 256306)
肌内脂肪含量与大理石花纹是肉牛非常重要的经济性状之一,提高这2 个指标可为国内牛产业带来巨大的经济效益[1-2]。脂肪型脂肪酸结合蛋白(Adipocyte Fatty Acid Binding-Protein,A-FABP),又称FABP4,是 脂肪酸结合蛋白(FABP)家族的重要成员之一[3]。早期研究发现,A-FABP 的主要功能为加强脂肪酸的转运扩散、促进细胞膜吸附脂肪酸、缓解不饱和脂肪酸对细胞的损伤作用、调节长链脂肪酸的氧化供能及磷脂和甘油三酯的代谢[4];调节细胞内脂肪酸浓度,调控多种细胞内的生化反应过程,参与细胞内长链脂肪酸隔室化分布[5]。Gerbens 等[6]首次发现猪A-FABP 基因内含子上的1 个微卫星,其多态性与肌内脂肪含量呈显著正相关;近年来研究发现牛A-FABP 基因在脂肪组织中表达量较高,其表达活性增强与脂肪细胞的分化有显著性关系[7],此外还与代谢综合征、糖尿病等疾病有关。由此推测,A-FABP 是影响肌内脂肪沉积重要的候选基因,在畜牧业和医学领域都有着广阔的应用前景。
布莱凯特黑牛是青岛农业大学科研成果转化基地及教学实习基地即山东布莱凯特科技股份有限公司2009年培育而成的优良肉牛种质资源[8],2015 年经国内肉牛权威专家鉴定其为新种群[9]。本实验采用荧光定量 PCR 技术探究黑牛不同组织、不同时期中的 A-FABP 基因 mRNA 的表达规律;采用 PCR-SSCP 方法分析其与肌内脂肪含量的相关性,为研究黑牛A-FABP 基因的生物学功能及其表达奠定理论基础,为布莱凯特黑牛肉质性状的分子选育提供参考。
1 材料与方法
1.1 实验材料 实验动物来自青岛农业大学科研成果转化基地及教学实习基地山东布莱凯特黑牛科技股份有限公司,选择饲养条件一致、体况相近、健康的黑牛12 头,分别采集2、6、10、12 月龄背最长肌,每时间点各3 头,立即放入液氮保存;黑牛宰杀后迅速采集甲状腺、胰腺、肾上腺等组织各2~5 g,立即放入液氮保存;实验动物为96 头去势后育肥的布莱凯特黑牛(25 月龄),批次与育肥环境均相同,采集背最长肌肉样,并进行肌内脂肪含量测定。
1.2 RNA 的提取及cDNA 的合成 用Trizol 法从背最长肌中提取RNA,用全式金反转录试剂盒将提取的RNA 反转录为cDNA,具体操作步骤见参考文献[10]。1.3 设计引物与合成 根据NCBI 数据库中牛A-FABP基因(GenBank 登录号:NM_174314.2 )和GAPDH基 因(GenBank 登 录 号:NM_001034034.2), 使 用Primer Premier 5.0 软件设计引物,引物由青岛擎科梓熙生物技术有限公司合成,引物信息见表1。
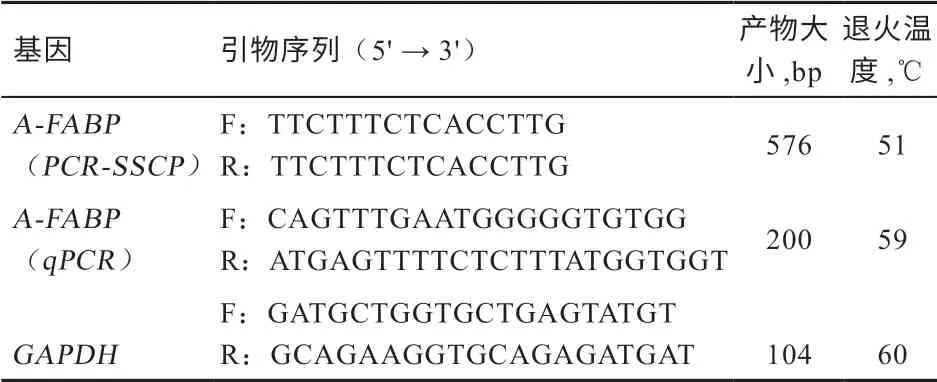
表1 引物信息
1.4 PCR 扩增目的片段 对目的片段进行PCR 扩增,扩增体系25 µL:上、下游引物(10 pmol/ µL)各0.5 µL,cDNA(50 ng/ µL)1 µL,MIX 12.5 µL,超纯水10.5 µL。反应程序:94 ℃预变性5 min,94 ℃变性30 s,特异的退火温度退火30 s,72℃延伸40 s,共45 个循环,72℃再延伸7 min,最后降至4℃。
1.5 A-FABP 基因的克隆与测序及生物信息学分析 对PCR 扩增产物进行胶回收,通过T 载体连接试剂盒将目的基因连接到p MD19-T 载体上,具体步骤见pGM-T克隆试剂盒[11];转化DH5ɑ 感受态细胞,摇菌,培养12 h 后,PCR 检测后获得的阳性克隆产物,送至上海生工生物有限公司进行测序。
利用DNAMAN 软件进行基因序列比对[11];采用MEGA 4.0 软件构建进化树;利用DNAstar 中的Protean程序对A-FABP 蛋白质的二级结构进行分析预测;高级结构预测采用(http://www.expasy.org/seissmod/swissmod-el.html);蛋白互作网络采用STRING 在线软件进行构建。
1.6 A-FABP 基因的表达规律与统计分析 反应体系(20 μL):iTaqUniwersal SYBR Green Supermix 10 μL,RNAfree Water 8 μL,上、下游引物各0.5 μL,模板cDNA 1 μL。反应程序:94℃预变性10 min;94℃变性30 s,60℃退火30 s,72℃延伸40 s,40 个循环[11]。每个样品设置3 次重复,采用2-ΔΔCT值法计算各样本中A-FABP相对于内参基因GAPDH 的表达量。利用SPSS 17.0 软件t 检验对数据进行差异显著性分析,结果以平均值±标准差的形式表示[11]。
1.7 A-FABP 基因PCR-SSCP 基因分型及其肌内脂肪含量统计分析 对扩增的目的片段进行硝酸银染色。操作流程:取2 µLPCR 产物、8 µL 配置好的变性缓冲液[98%去离子甲酰胺、10 mmol/L EDTA (pH 8.0)、0.025%二甲苯氰FF、0.025%嗅酚蓝]加到PCR 管内,98℃变性6 min,变性后冰浴10 min,采用14%丙烯酰胺凝胶(Acr: Scr=29 :1)进行检测,180 V 电泳10 h 结束后银染。
统计图谱中各基因型的数量,利用PopGen32 软件计算多态位点的等位基因频率、基因型频率、有效等位基因数(Ne);通过c2 适合性检验检测群体内基因型分布是否符合哈德-温伯格(Hardy-Weinberg)平衡定律;根据固定效应模型,应用SPSS 17.0 软件分析A-FABP基因不同基因型对肉质性状的影响效应。
2 结果与分析
2.1 A-FABP 基因和GAPDH 基因的PCR 扩增 由图1 可知,引物扩增的条带清晰且单一,A-FABP 基因产物大小分别为576 bp 和200 bp,GAPDH 基因产物为104 bp,与目的片段大小一致,表明引物特异性强,可以用于后续的克隆测序以及荧光定量PCR 实验。
2.2 A-FABP 生物信息学分析 生物信息学分析结果表明A-FABP 基因CDS 序列全长399 bp,编码132 个氨基酸,利用softberry 在线网站构建DNA 序列可视化3D结构,如图2 所示。通过DNAMAN 软件对获得的布莱凯特黑牛A-FABP 基因CDS 序列与牛(100.00%)、瘤牛(99.25%)、水牛(98.25%)、野猪(89.97%)、山羊(96.24%)、绵羊(95.99%)、人(89.47%)、大鼠(70.64%)和原鸡(72.68%)进行同源性比较,结果发现除大鼠(70.64%)和原鸡(72.68%)外,黑牛与其他脊椎动物的同源性均大于80%;采用 UPGMA 法对各物种进行聚类分析(图3),结果显示布莱凯特黑牛与牛、瘤牛和水牛相距最近,其次与绵羊、山羊相距较近;原鸡与上述动物相距最远。
2.3 A-FABP 基因编码蛋白的特征分析 布莱凯特黑牛A-FABP 蛋白的分子式为C646H1043N171O201S8,分子量为21 907.97,其等电点5.52,说明是偏酸性蛋白;不稳定系数为50.14;脂溶性指数为82.58,大于60,蛋白流动性大;总平均亲水性为-0.198,即水溶性蛋白;其摩尔消光系数为14 105。信号肽预测结果显示该蛋白质的 C-值、S-值、Y-值和D-值均小于0.5,期望跨膜螺旋数为0。A-FABP 蛋白的二级结构预测显示A-FABP蛋白以 α-螺旋和无规卷曲为主,含有少量的β-折叠和β-转角。利用SWISS-MODEL 对A-FABP 蛋白的三级结构进行在线预测,结果见图4。利用STRING 构建蛋白质互作网络(图5),圆圈代表蛋白质,该蛋白与同家族的FABP1、FABP6 蛋白互通性最好,其次是LIPE、PLIN1、PPARG 蛋白互通性较好。

图1 琼脂糖凝胶电泳检测图

图2 A-FABP DNA 序列可视化3D 结构
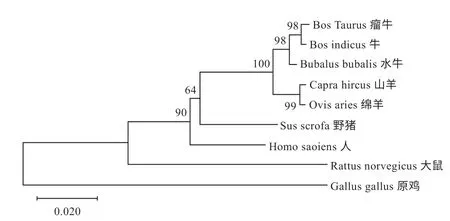
图3 A-FABP cDNA 序列构建的UPGMA 系统发生树

图4 A-FABP 蛋白的三级结构预测
2.4 实时荧光定量PCR 结果 由图6 可知,A-FABP 和GAPDH 基因熔解曲线峰值单一、无杂峰,未发现引物二聚体和非特异性扩增的情况,可知引物设计特异性良好,可用于下一步数据统计分析。
2.5 A-FABP 基因在不同组织中的表达分析 图7-A 显示, A-FABP 在背最长肌中表达量最高,其次是肾脏,均高于等其他组织(P<0.01);A-FABP 在胰腺、甲状腺、肺脏、脾脏、睾丸、肝脏、心脏中表达量逐渐降低且差异不显著。
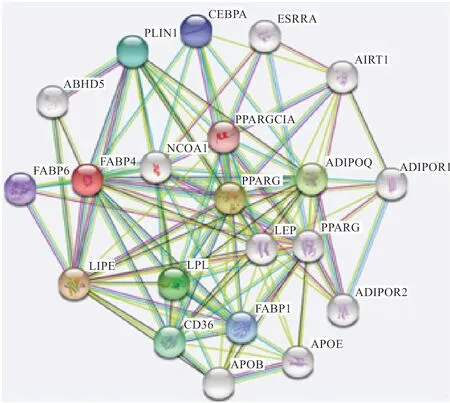
图5 差异表达基因蛋白质互作网络图
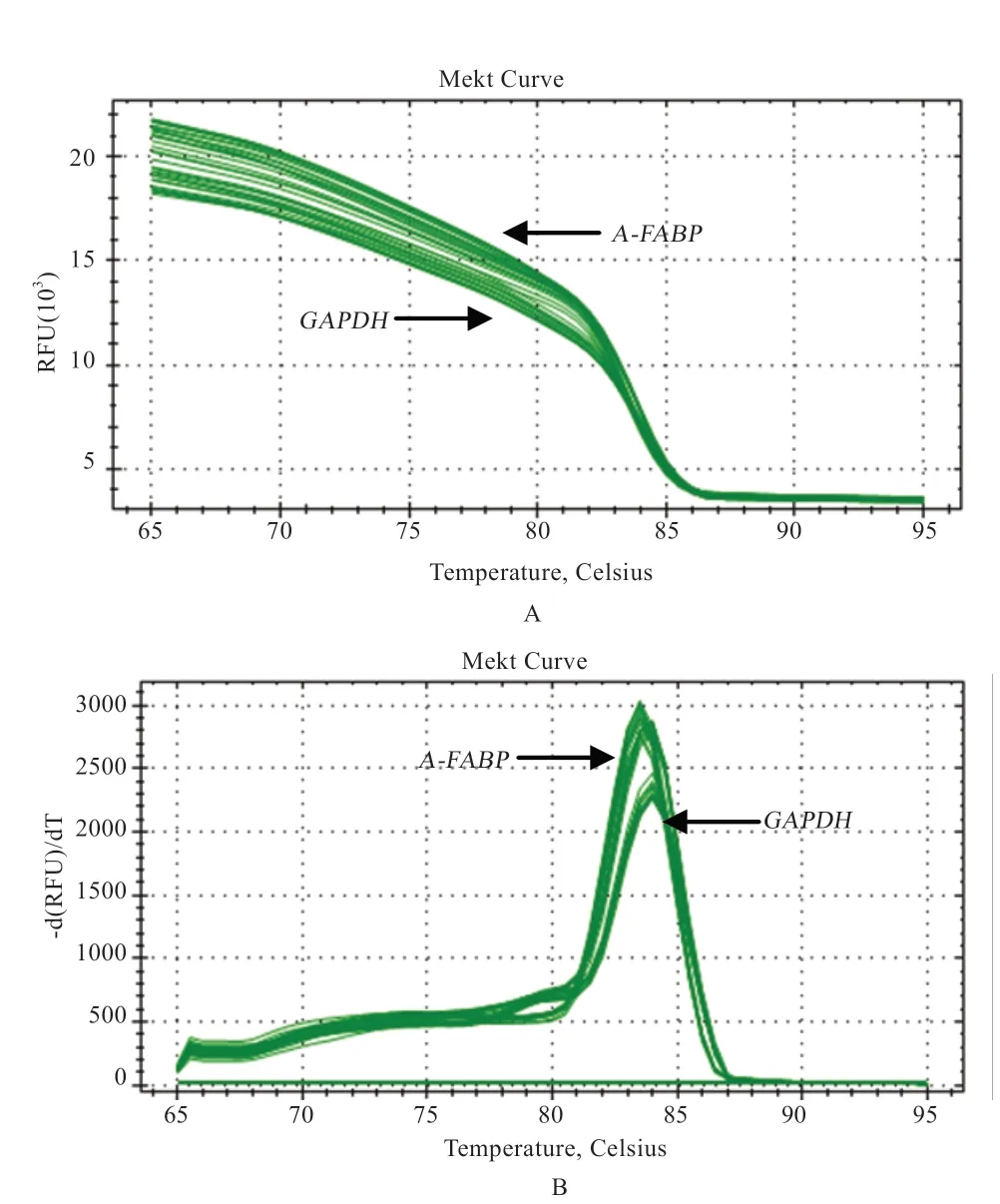
图6 A-FABP 基因和GAPDH 基因荧光定量扩增曲线(A)和熔解曲线(B)

图7 A-FABP 在不同组织(A)及不同时期背最长肌(B)中的表达水平
由图7-B 可知,A-FABP 在4 个不同时期背最长肌中均有表达,2 月龄的表达量最高且高于其他3 个时期(P<0.01); 6、10、12 月龄的表达量差异不显著,随着月龄的增加A-FABP 的表达量呈现逐渐降低的趋势。2.6 A-FABP 基因基因型及基因频率的分布 PCR 扩增产物经PCR-SSCP 后带型分布见图8,扩增区发现3 种带型,即AA、BB、AB 基因型,2 种等位基因A、B。在黑牛第1 内含子区存在多态性(表2),优势等位基因为A,其等位基因频率为0.651。根据c2 独立性检验矫正公式,检验表明黑牛基因频率和基因型频率符合哈德-温伯格平衡(P>0.05)。
2.7 A-FABP 基因不同基因型与肌内脂肪含量的相关性分析 由表3 可知,AA、AB、BB 基因型肌内脂肪含量均值较高,肌内脂肪含量关系为AA>BB>AB,AB 基因型低于其他基因型(P<0.05);在大理石花纹评分方面,不同基因型差异不显著。

图8 布莱凯特黑牛A-FABP 基因PCR-SSCP 检测图

表2 布莱凯特黑牛A-FABP 基因PCR-SSCP 基因分型及基因频率

表3 不同基因型肌内脂肪含量和大理石花纹评分均值
3 讨 论
3.1 A-FABP 基因克隆与生物信息学分析 本研究结果显示,A-FABP 基因CDS 序列全长为399 bp,具有完整的阅读框,编码132 个氨基酸。同源性分析发现,不同来源的A-FABP 在CDS 序列上具有较高的同源性,其中与牛、瘤牛、山羊、绵羊、人均大于80%,说明该基因CDS 序列在脊椎动物中相对保守,在进化过程中具有高度保守性,与“重要功能蛋白氨基酸置换率较低”[12]的理论一致。生物信息学分析显示,A-FABP 蛋白偏酸性,表现亲水性,说明该蛋白为水溶性蛋白,其二级结构很容易接近水分子[13]。二级结构发现,α- 螺旋和无规卷曲为主要结构元件,含有少量 β-折叠,易于形成球状蛋白,属于all-alpha 型[14],蛋白质的水溶性较好,这与上述预测的蛋白质基本性质中的总平均亲水性相一致。A-FABP 蛋白三级结构均以一条多肽链构成,呈桶状,说明该基因编码蛋白的功能区域在进化过程中保守性较强。该结果符合“多基因家族的协同进化”[15]理论,与上述同源比对中揭示的物种进化关系相一致。
3.2 A-FABP 基因相对定量表达分析 脂肪型脂肪酸结合蛋白属于脂肪酸结合蛋白家族,其家族包括至少9 种蛋白,早期研究发现该家族蛋白在细胞脂肪酸代谢及转运中发挥重要作用[15]。进一步研究发现,在哺乳动物中A-FABP 蛋白主要在脂肪细胞中表达,参与甘油三酯形成,调控脂肪沉积[16],推测参与肌内脂肪沉积的相关调控过程。本研究发现,A-FABP 在背最长肌中表达量最高,其次为肾脏,在胰腺、甲状腺、肺脏、脾脏、睾丸、肝脏、心脏中表达量低且差异不显著,这与研究猪A-FABP 基因的表达差异性比较时其主要沉积于肌肉组织中的结果相一致[17]。邝良德等[18]指出,除脂肪组织外,A-FABP 可在九龙牦牛的骨骼肌、肾、脾中检测到表达。荣光辉等[19]研究发现,北京鸭 A-FABP 基因主要在脂肪组织中表达。本研究选取组织有限,9 种组织A-FABP 在背最长肌表达量最高,其是肌内脂肪含量的相关基因,A-FABP 基因的脂肪沉积形式及部位存在差异性,还需要进一步研究。本实验发现A-FABP 在2月龄的表达量最高,12 月龄的表达量最低,随着月龄增加A-FABP 表达量呈逐渐降低的趋势。周志楠等[20]研究发现,巨型玫瑰冠鸡不同时期A-FABP 基因的 mRNA 表达量也随着周龄增长呈下降趋势;晁珊珊等[21]研究发现,幼鱼A-FABP 基因表达水平显著高于较大的鱼和成鱼,A-FABP 基因表达随着鱼类的发育呈下降趋势。综上推测,A-FABP 基因在布莱凯特黑牛肌内脂质代谢过程中发挥重要作用。
3.3 A-FABP 基因不同基因型与肌内脂肪含量的相关性分析 Gerbens 等[6]研究发现,梅山猪A-FABP 基因的多态性与背最长肌上的肌内脂肪含量显著相关。进一步研究发现,秦川牛A-FABP 的突变与肉质性状指标相关[22]。由上述研究成果推测,该基因可以作为肉牛肉质性状标记辅助选择的候选基因。本研究发现,在黑牛第1内含子区存在多态位点,对肌内脂肪含量影响显著,AA>AB>BB,表明AA 基因型对肌内脂肪含量影响显著。近期研究发现,长白猪A-FABP 与肌内脂肪含量存在显著相关[23];苏姜猪、姜曲海猪、长白猪群体A-FABP的遗传多态性对肉质性状的影响显著[24],与本实验结果一致。综上表明A-FABP 与肌内脂肪含量显著相关,对调控肌内脂肪含量发挥重要作用,可以作为肉质性状的候选基因。
本实验中,不同基因型大理石花纹评分差异不显著。周国利等[25]利用PCR-RFLP 技术对鲁西黄牛研究发现,A-FABP 对大理石花纹的影响不显著。高妍等[26]研究黑猪和白猪A-FABP 基因的遗传多态性,结果证实A-FABP 基因与大理石纹显著相关,与本研究结果不一致。分析原因可能是:一是实验用牛的品种不同,与遗传背景有很强的相关性;二是大理石花纹的等级评分具有个人的主观判断性,不同的评定者对同一块肉的评分会有一定的差异;三是采用的评分标准可能不同,具体差异分析有待进一步研究。
4 结 论
综上推测,A-FABP 基因参与布莱凯特黑牛肌内脂质代谢相关过程,对调控脂肪的沉积发挥重要作用,可作为肉质性状的辅助标记基因,为布莱凯特黑牛的选育提供分子遗传学依据。
