亚热带地区5种土地利用模式土壤微生物生态特性的比较
2019-09-18杨满元杨宁吴磊姜琳万丽陈璟范适
杨满元,杨宁,吴磊,姜琳,万丽,陈璟,范适
(湖南环境生物职业技术学院园林学院,湖南 衡阳 421005)
在森林生态系统中,微生物是土壤活有机质,土壤养分重要来源,其多少反映了参与调控土壤中能量和养分循环以及有机质转化的微生物数量[1-2],其中土壤微生物生物量碳、氮和磷能够敏感且及时地反映土壤质量变化[3].土壤细菌、真菌和放线菌是土壤生态系统中最重要的3大微生物群,其组成和数量的变化是表征土壤环境质量最为重要的生物学指标[4].目前,土壤微生物生物量和土壤微生物数量作为土壤健康的生物指标来评价生态系统的恢复进程和指导生态系统管理等已逐渐成为研究热点[5].
杉木是中国南方广泛应用的造林树种,在中国乃至世界的森林资源中占十分重要的地位,但随其栽植面积扩大,生态问题严重突出,其生产力下降,林地土地肥力递减,严重阻碍了杉木人工林的持续生产与发展.国内外许多学者从生态系统养分循环、生物生产力、生态经济效益等角度对混交林进行了一系列的研究,取得了很多积极成果[6].不同树种的混交林可缓解单一树种长期种植对土壤的危害,但这种作用取决于树种的组成[7].樟树是我国亚热带代表性常绿阔叶树种,有报道指出樟树可在一定程度上提高土壤微生物的活性[8].本文从土地利用模式角度,对比研究撂荒地、纯杉木林、纯樟树林、杉木樟树混交林及次生阔叶林5种土地利用模式0~10、10~20、20~30 cm土层的土壤微生物特征以及与土壤理化性质的关系,揭示不同土地利用模式的物质代谢和肥力发展规律,为不同林分的可持续经营与管理及其生态恢复提供决策参考.
1 材料与方法
1.1 研究区概况
试验地位于湖南省长沙市南郊的天际岭国家森林公园N 28°06′~28°07′,E 113°02′~113°01′,其地层主要是第四纪更新世的冲积性网纹红土和砂砾,属典型的红壤丘陵区,地形以岗地为主;海拔46~116 m,坡度5°~25°,属典型的亚热带湿润季风气候,雨量充沛,年平均降雨量1 422 mm,主要集中于4~6月;年平均气温17.2 ℃,1月最冷(平均气温4.7 ℃,极端最低温度-11.3 ℃),7月最热(平均气温29.4 ℃,极端最高气温40.6 ℃);无霜期为270~300 d,日照时数平均1 677 h.园内小生境众多,植物种类达到2 200余种,植被以人工林为主.
1.2 样品采集
2014年5月上旬,选取纯杉木林(Cunninghamialanceolataforest,CLF)、纯樟树林(Cinnamomumcamphoraforest,CCF)、杉木樟树混交林(C.lanceolata-C.camphora,forest,CLCCF)、次生阔叶林(Secondary broad-leaved forest,SBF)以及附近撂荒地(Abandoned land,AL)5种土地利用模式作为样地,他们均位于海拔100 m高的丘陵坡地上,设置样地试验前均无农业措施等人为干扰,坡向/坡度(NW 20°~23°)相似,其中撂荒地(撂荒5 a)主要的草本植物为狗尾草(Setariaviridis)、淡竹叶(Lopnamerumgracile)以及少量的灌木盐肤木(Rhuschinensis)、六月雪(Serissaserissoides)等;纯杉木林、纯樟树林、杉木樟树混交林和次生阔叶林下层有少量白栎(Quercufabei)、美丽胡枝子(Lespedezaformosa)等灌木,草本层主要有刺芒野古草(Arundinellasetosa)、五节芒(Miscanthusfloridulus)与狗脊蕨(Woodwardiajaponica)等,5种土地种用模式样地的面积均大于1 hm2的样地(表1).在每个样地中取3个20 m×20 m的临时标准样方,采用“Z”5点取样法,采集0~10、10~20、20~30 cm土层土壤,重复3次,同层混匀带回实验室.一份自然风干供土壤理化性质测定;另一份保存于4 ℃冰箱中以备土壤微生物测定,并在10 d内完成土壤微生物生物量的分析.
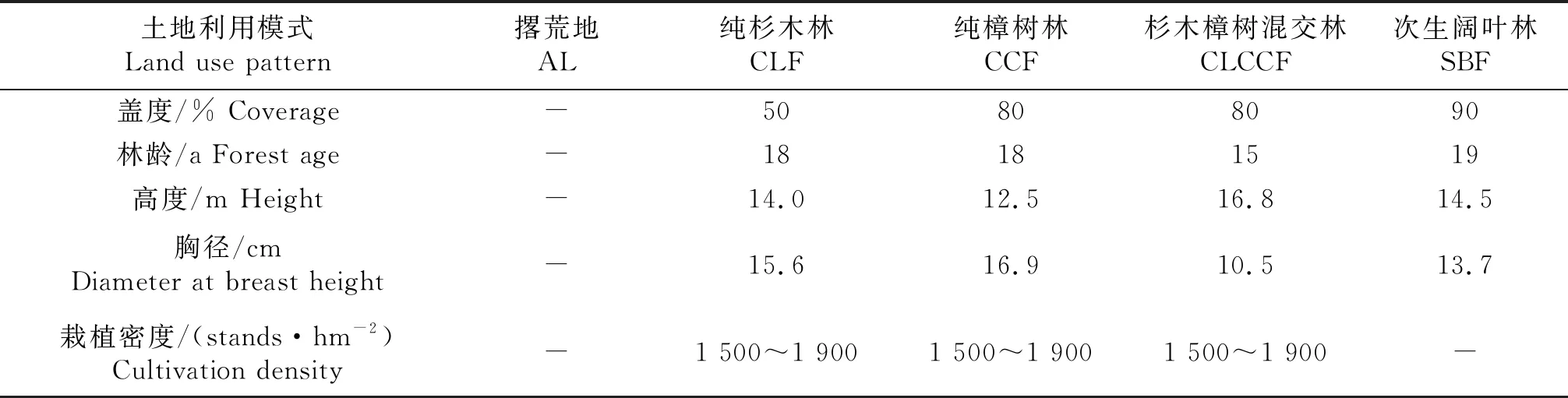
表1 土地利用模式概况
1.3 测定指标与方法
1.3.1 土壤理化性质 土壤理化因子包括土壤有机碳(soil organic carbon,SOC),全氮(Total nitrogen,TN),全磷(total phosphorus,TP),全钾(total potassium,TK),碱解氮(alkali-hydrolyzable nitrogen,AN),速效磷(available phosphorus,AP)以及速效钾(available potassium,AK)以及土壤pH值,测定方法为常规方法,参考文献[9].各土地利用模式理化性质特征见表2.
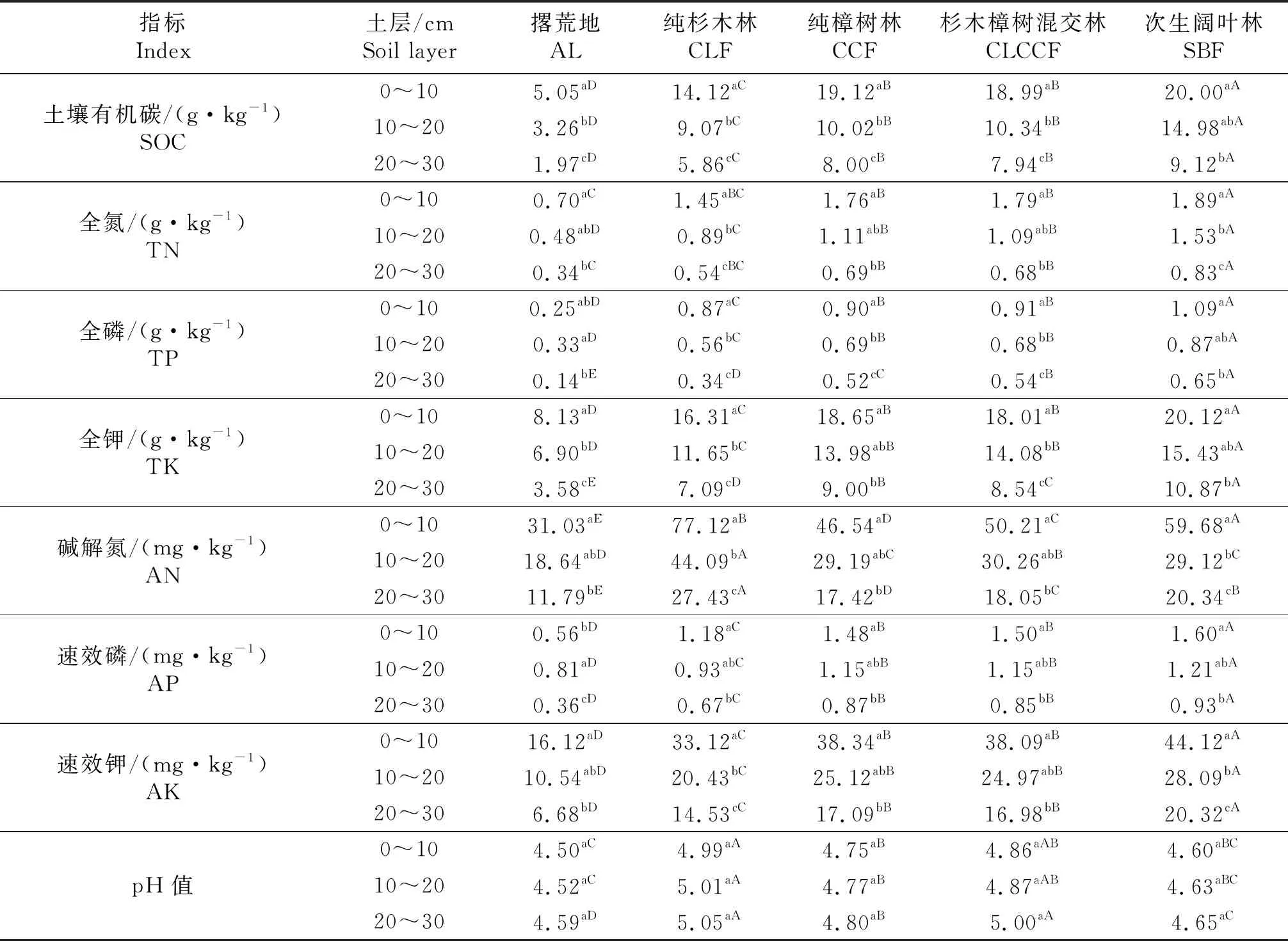
表2 土壤理化性质
不同大写字母表示相同土层不同土地利用模式差异显著,不同小写字母表示相同理化性质不同土层差异显著(P<0.05).
Different capital letters indicate significant differences in different use patterns at the same soil layer.Different lowercase letters indicate significant differences among different soil layers at the same physio-chemical property atP<0.05.
1.3.2 土壤微生物性质 土壤微生物数量采用稀释倾注平板涂布法测定.细菌、真菌和放线菌的培养基分别为牛肉膏蛋白胨、马丁-链霉素(链霉素30 mg/L)培养基和改良高氏1号(苯酚500 mg/L)培养基[10].土壤微生物生物量碳(soil microbial biomass carbon,SMBC)、土壤微生物生物量氮(soil microbial biomass nitrogen,SMBN)和土壤微生物生物量磷(soil microbial biomass phosphorus,SMBP)的测定方法分别参考文献[11]~[13].
1.4 数据分析
采用SPSS 13.0软件进行数据处理与统计分析.用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较不同数据间的差异,用Pearson分析法分析微生物生物量碳、氮和磷与理化因子的关系.表中数据为3次重复的均值.
2 结果与分析
2.1 土壤微生物数量特征
从表3可以看出,在相同土地利用模式中,土壤微生物总数随土层深的增加而显著减小(P<0.05),相同土层,次生阔叶林>纯樟树林(≈杉木樟树混交林)>纯杉木林>撂荒地(P<0.05);从不同类群微生物数量上看,细菌和真菌数量的大小顺序为次生阔叶林>纯樟树林(≈杉木樟树混交林)>纯杉木林>撂荒地(P<0.05),而放线菌数量的大小顺序为纯杉木林>次生阔叶林>纯樟树林(≈杉木樟树混交林)>撂荒地(P<0.05).在微生物区系中,每个土层均以细菌占绝对优势,占85%以上,其中在撂荒地细菌所占的比例最大,高达95.68%,最少为放线菌,仅占微生物总数的0.75%.
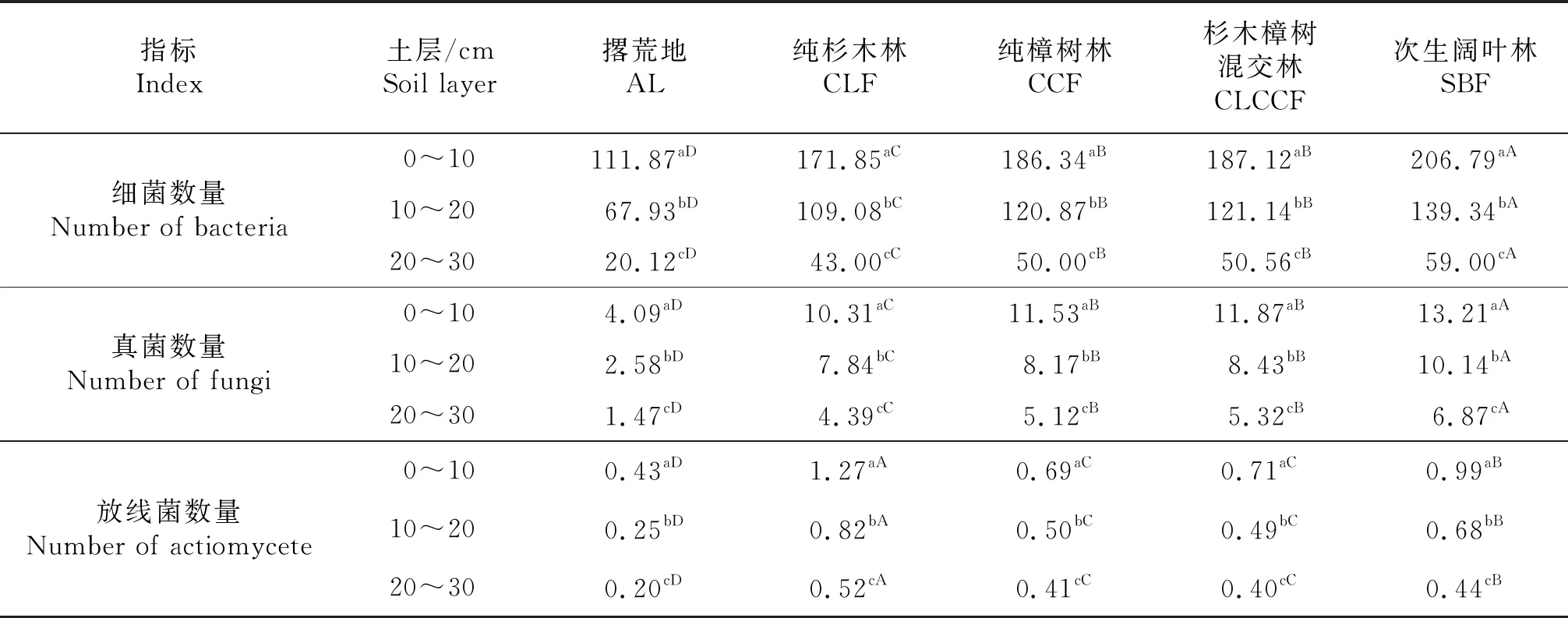
表3 土壤微生物数量
2.2 土壤微生物生物量
由表4可知,5种土地利用模式土壤微生物生物量碳普遍较低,均在50.00 mg/kg以下.5种土地利用模式的土壤微生物生物量碳均随土层深度的增加而显著减小(P<0.05).相同土层不同土地利用模式土壤微生物生物量碳的大小顺序为次生阔叶林>纯樟树林(≈杉木樟树混交林)>纯杉木林>撂荒地(P<0.05),与土壤养分的变化趋势基本一致(表2).同样,不同土地利用模式土壤微生物生物量氮和土壤微生物生物量磷也呈现不同的变化规律.随着土层深度的增加,5种土地利用模式的土壤微生物生物量氮以及纯杉木林、纯樟树林、杉木樟树混交林和次生阔叶林的土壤微生物生物量磷均表现出随着土层深度的增加而显著减小(P<0.05),而撂荒地的土壤微生物生物量磷随着土层深度的增加呈现先增后减的规律.在5种土地利用模式中,0~10 cm土层的土壤微生物生物量氮、土壤微生物生物量磷的变化规律与土壤微生物生物量碳的变化规律相似,次生阔叶林土壤微生物生物量氮(4.43 mg/kg)、土壤微生物生物量磷(18.16 mg/kg)最高,撂荒地最低,分别仅为次生阔叶林的16.70%、28.19%.
2.3 土壤微生物生物量与理化性质的相关性
在纯杉木林、纯樟树林、杉木樟树混交林与次生阔叶林4种土地利用模式中土壤微生物生物量均与理化性质正相关(P<0.01),撂荒地,土壤微生物生物量只与土壤有机碳、全钾、碱解氮、速效磷、速效钾呈正相关(P<0.05或P<0.01),土壤微生物生物量碳、土壤微生物生物量磷与全氮之间以及土壤微生物生物量磷与全磷之间相关性达极显著水平(P<0.01),而土壤微生物生物量碳、土壤微生物生物量磷与全磷之间的相关性不明显(P>0.05,表5).由此可见,在纯杉木林、纯樟树林、杉木樟树混交林与次生阔叶林4种土地利用模式中土壤微生物生物量之间关系极其密切,且与理化因子的相关性极强.
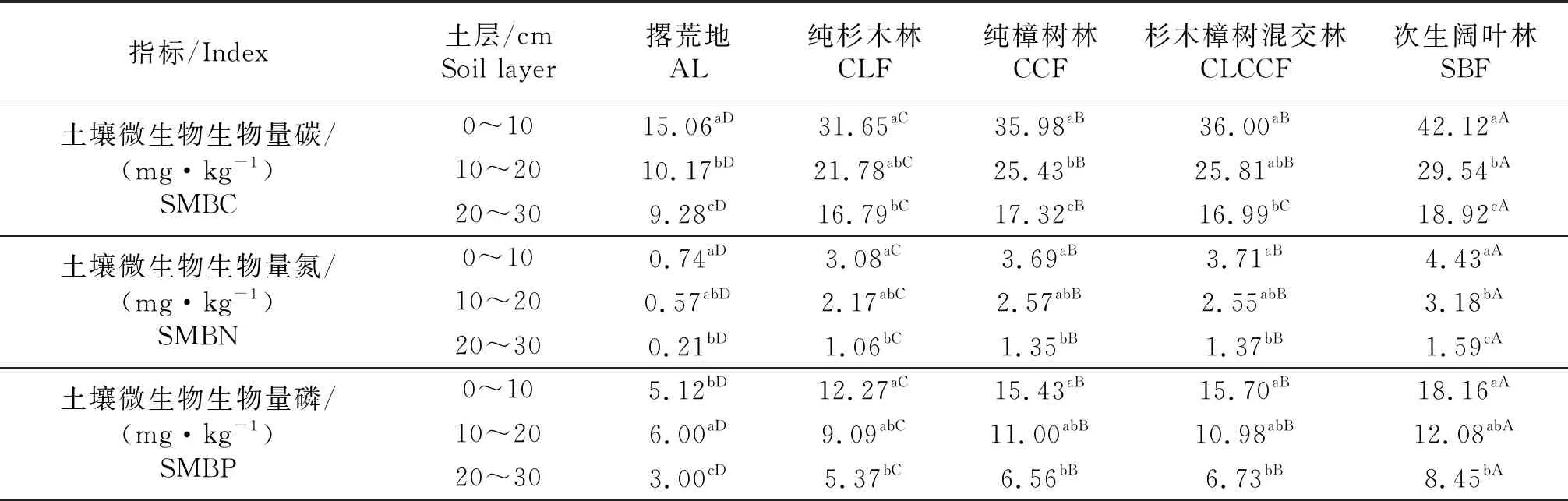
表4 土壤微生物生物量
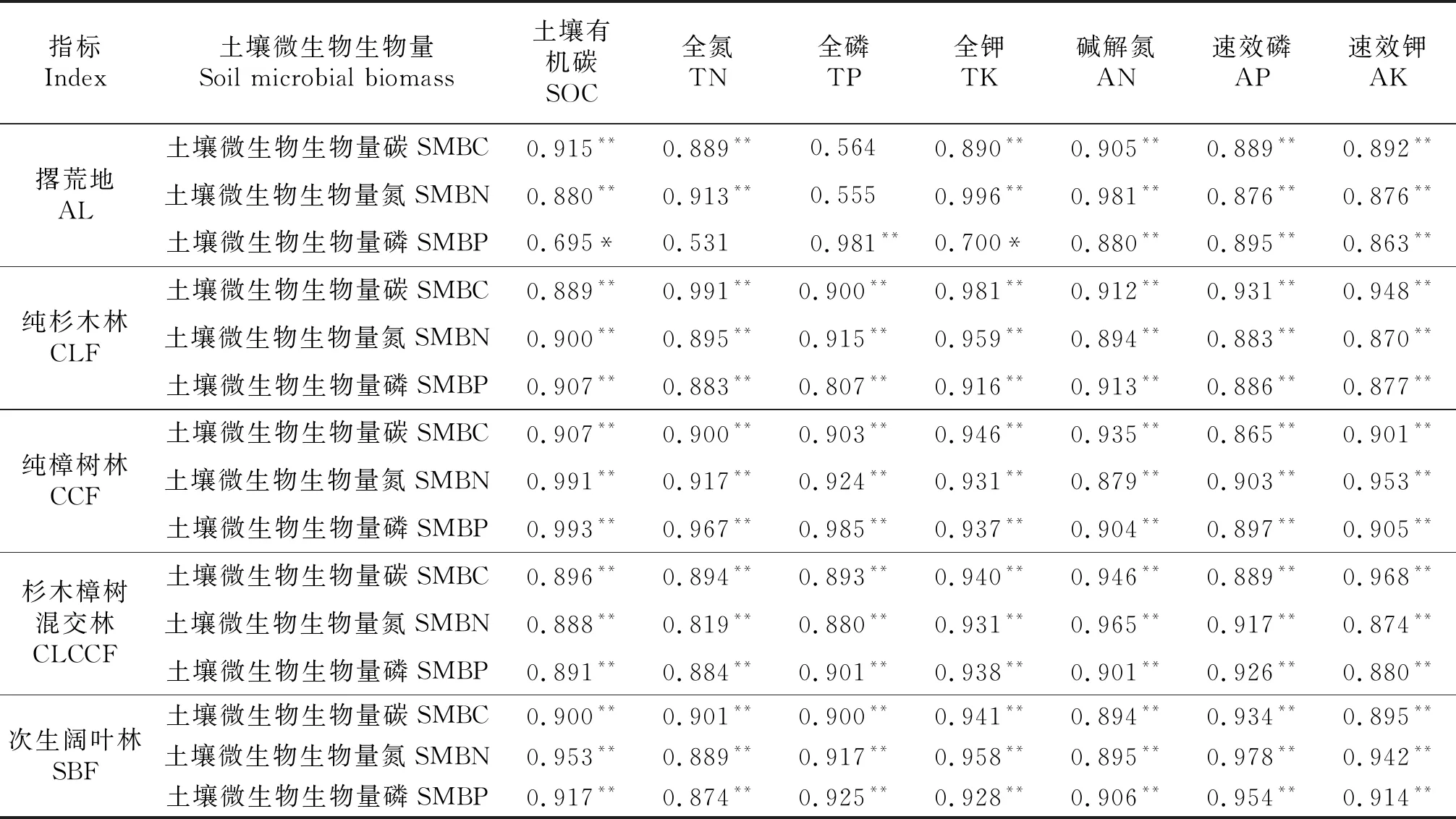
表5 土壤微生物生物量与理化性质间的相关性
**P<0.01,*P<0.05.
3 讨论
研究发现,土壤微生物数量与微生物生物量在不同的土地利用模式以及不同的土层呈现出显著的数量差异(表3~4).撂荒地、纯杉木林、纯樟树林、杉木樟树混交林以及次生阔叶林5种土地利用模式,植被凋落物以及根系主要集中于土壤表层,凋落物和根系分解所产生的有机碳相对集中于土壤表层,表层营养源充足(表2),加之表层的水热与通气状况良好,有利于土壤微生物的生长与繁殖,随着外界环境的变化,表层经历了更为剧烈的温湿度变化,更容易受到分解物和根系分泌物的影响,根系分泌物种类和数量的增加有利于土壤微生物多样性的增加[14-15],因此,随着土层深度的增加,5种土地利用模式的微生物数量与微生物生物量随土层增加而显著减小(P<0.05).次生阔叶林物种丰富、凋落物量多质好,土壤微生物总量与微生物生物量最高[16];纯樟树林以及杉木樟树混交林植被覆盖度较高,凋落物的输入量与多样性比纯杉木林和撂荒地要大,基质质量较优的凋落物分解速度较快(樟树凋落物和细根的平均分解系数分别为0.98和0.65[17]),其土壤具有较高的氮素有效性,相对肥沃的土壤形成有机矿质复合体能借助物理机制保护来源于分解过程中进入土壤的有机质,从而储存更多的土壤有机碳和氮,为土壤微生物提供更多的碳源和氮源,土壤微生物总数与微生物生物量较高[18-19];虽然纯杉木林的凋落物数量较高,但凋落物纤维素和半纤维素含量较高,矿物质和其他元素含量较低,腐质化系数较低,凋落物难以分解,而树木生长又需要养分,因此,纯杉木林容易造成土地退化,土壤微生物总数与微生物生物量较低[20-21];撂荒地草本稀疏,地表裸露大,土壤侵蚀严重,土壤水分蒸发强烈,抑制了土壤酶活性,加之生物量低,凋落物少,有机质矿化加剧,导致土壤微生物数量最少,微生物生物量最低,这一变化趋势与一般演替规律相符合,与土壤有机碳等养分含量规律一致(表2),同时也说明,在亚热带红壤丘陵区不同的土地利用模式中,可将土壤微生物数量与微生物生物量作为评价不同土地利用模式的土壤肥力生物学指标,通过加强森林抚育管理,减少人为干扰,以及不同改变土地利用模式的手段为土壤微生物的生长繁殖创造了优越环境,促进土壤微生物群落发育,同时也提示在森林的林分管理过程中,可施菌肥来提高土壤生化强度,促进亚热带红壤区土地质量的改善[22-23].
由于细菌,真菌和放线菌的生物学特征的差异,细菌呼吸机制复杂,营养类型多,适应能力强,代谢旺盛,繁殖快速,成为土壤中的优势群,占土壤微生物的85%以上,真菌次之,放线菌最少(表3),与徐惠风等[24]对长白山沟谷乌拉苔草(Carexmeyeriana)湿地土壤微生物群落研究结果基本一致,而与南丽丽等[25]对甘肃荒漠灌区的研究结果不一致,主要原因为该亚热带地区5种土地利用模式的土壤pH值处于4.50~5.05之间,偏酸性(表2),而甘肃荒漠灌区土壤整体偏盐碱性,pH在8.42~8.70之间,真菌具有对酸性土壤的偏好,而放线菌具有喜热耐碱的特征,喜欢在偏碱性环境中生长,因此,在本研究中,真菌和数量相对于放线菌较多[26];同样,在本研究中,细菌和真菌以次生阔叶林最高,纯樟树林和杉木樟树混交林次之,撂荒地最低,而放线菌则以纯杉木林最高,显著高于其它土地利用模式(P<0.05,表3),这是因为纯杉木林土壤pH值显著高于其他土地利用模式土壤pH值(P<0.05,表2);另外,由于不同土地利用模式凋落物质与量的差异,根际微环境对三大微生物类群的生长与繁殖的影响不同,因此,其数量与比例也各异.
4 结论
本研究发现,5种土地利用模式的土壤微生物生物量均与土壤有机碳正相关(P<0.05或P<0.01),说明土壤有机碳是影响土壤微生物生物量重要因素;纯杉木林、纯樟树林、杉木樟树混交林以及次生阔叶林土壤微生物生物量与土壤有机碳、全氮、全磷、全钾、碱解氮、速效磷和速效钾均呈正相关(P<0.01),说明土壤养分是影响土壤微生物生物量主要因素.在撂荒地中,土壤微生物生物量碳、土壤微生物生物量磷与土壤有机碳、氮和钾含量呈正相关(P<0.01),但与全磷含量的相关性不明显(P>0.05,表5),这与彭佩钦等[27]的研究结果基本一致,但与蔡晓布等[28]的研究结果相异,原因可能与土壤对氮、磷、钾等物理及化学固定作用,氮素的挥发、淋溶损失等各种因素的影响有关.由此可见,亚热带地区不同土地利用模式土壤微生物与养分之间具有紧密的联系,土壤微生物生长有赖于土壤肥力水平和环境状况,土壤微生物与土壤肥力之间相互促进、协同发展(表5)[29-30].因此,在亚热带地区地区的林分管理及生态恢复中,可采用改变土地利用模式的手段,改善土壤微生物特性,促使土壤以及整个生态系统朝更高级的、健康的方向发展.
