芥菜TCP转录因子家族全基因组鉴定及表达分析
2019-09-17李坤杰谭杉杉闵爱玲李梦瑶
李坤杰,谭杉杉,孙 勃,饶 丹,何 琦,闵爱玲,李梦瑶
(四川农业大学园艺学院,成都 611130)
转录因子(transcription factor),是一类能够识别并结合特定序列DNA 结合域的启动子区域、并以此调节新陈代谢或发育过程的蛋白质[1]。TCP 是植物特有的转录因子家族之一,其基因的命名来自3 个不同物种的4 个基因的首字母缩写,分别是玉米 TB1、金鱼草 CYC、水稻 PCF1 和 PCF2[2-4]。这 4 个基因所编码的蛋白质共同点是在其N 末端都含有一段高度保守的60 个左右的氨基酸残基,又名TCP结构域,能够形成一个非典型的碱性-螺旋-环-螺旋的结构[5-6]。根据TCP 结构域的氨基酸序列差异,该家族可分为两个亚组:ClassⅠ和ClassⅡ,ClassⅠ又名PCF,ClassⅡ还可继续分为两个小组,分别为CIN 和 CYC/TB1[7]。
自1999年来,TCP 家族作为新的转录因子家族出现,对其功能的探索在不断加深。在TCP 命名的4个基因中,对基因的功能研究均显示它们参与了植物的生长发育。玉米TB1 抑制了侧生分生组织和雄花原基发育[2];金鱼草Cyc 基因调控花的对称发育[3];水稻PCFI 和PCF2 基因促进了分生组织中PCAN基因启动子的特异性表达[4]。最近研究表明,TCP 转录因子除了参与植物的生长发育,还与植物的免疫调控及响应逆境胁迫息息相关:CIN-like TCP 与植物抵御病原菌入侵的免疫反应有关[8];木薯MeTCP4基因在干旱及低温胁迫下表达受到抑制[9];水曲柳TCP4 响应寒冷、盐、干旱等非生物胁迫以及激素信号,说明其参与了植物逆境胁迫的调控[10]。
芥菜(Brassica juncea)是十字花科芸薹属重要的园艺作物。茎瘤芥是芥菜的一个茎用变种,其瘤茎在生长发育过程中,叶柄基部发生变态,形成1~5 个瘤状凸起,并随茎膨大而横向膨大[11]。茎瘤芥的瘤茎生长发育过程非常复杂,涉及到形态、生理、生化和遗传等多方面的变化。目前,对茎瘤芥的瘤茎膨大的机理研究尚不深入,尤其与茎膨大相关的功能基因的研究报道较少。TCP 家族基因广泛参与了植株的生长发育,目前的研究主要集中于拟南芥、水稻、玉米等植物中,对芥菜TCP 家族研究尚未见报道。
本研究,通过生物信息学方法,对芥菜基因组数据库进行检索,筛选并鉴定芥菜所有的TCP基因,并对这些基因的理化性质、结构特征以及基因表达等进行分析,以期为今后进一步研究芥菜TCP 基因的功能及参与茎膨大的调控机制提供理论基础。
1 材料和方法
1.1 芥菜TCP转录因子家族成员的检索与鉴定
从 Pfam 数据库(http://pfam.xfam.org/)下载 TCP家族模块序列(PF03634),从芥菜基因组数据库(http://brassicadb.org/)下载芥菜蛋白序列,利用HMMER 软件从芥菜基因组序列中检索TCP 蛋白序列[12]。将候选蛋白进一步提交至NCBI-CDD 和Pfam(http://pfam.xfam.org/)进行 TCP 结构域的确认。利用 ExPASy 网站(http://web.expasy.org/protparam/)对确认的TCP 蛋白序列进行蛋白质分子量、等电点和氨基酸大小预测,利用WOLF PSORT(http://www.genscript.com/wolf-psort.html)和 TargetP(http://www.cbs.dtu.dk/services/TargetP/)进行亚细胞定位分析。拟南芥和水稻的TCP 基因序列分别下载自TAIR(https://www.arabidopsis.org/)和 PlantTFDB(http://planttfdb.cbi.pku.edu.cn/)网站。
1.2 不同物种TCP家族系统进化分析
使用Clustal X 2.1 软件对芥菜、拟南芥和水稻的所有TCP 蛋白序列进行多序列联配[13],将多序列联配的结果利用MEGA 6.06 软件采用邻接法(NJ)法进行系统进化分析[14],校验参数bootstrap 重复1 000次,构建了系统进化树。
1.3 芥菜TCP家族基因的染色体定位分析
从芥菜基因组数据库(http://brassicadb.org/)下载芥菜基因组信息,找出TCP 家族基因成员的染色体位置及结构信息,并利用MapInspect(http://mapinspect.software.informer.com)软件绘制TCP 家族基因的染色体位置分布图。
1.4 芥菜TCP家族基因成员的序列结构和保守基序分析
登录 GSDS 服务器(http://gsds.cbi.pku.edu.cn/),通过比对编码序列与基因序列信息,绘制TCP 家族基因的内含子和外显子的结构模式图。利用MEME(http://meme-suite.org/)对芥菜 TCP 家族基因成员的保守基序进行分析,参数设定保守基序(motif)大小为10~100 个氨基酸,输出的结构域数目最大为15,motif 结构绘图采用 TBtools 软件[15]。
1.5 芥菜TCP家族基因在茎膨大不同时期的表达分析
茎瘤芥品种“涪杂2 号”不同发育阶段(茎的直径分别为2、4、6、8 cm)的转录组测序由本课题组完成,测序所获得的数据已提交至NCBI 网站,登录号为SRP151320。使用FPKM 值来反映TCP 基因在芥菜不同发育阶段的表达丰度,基因表达热图使用Microsoft Excel 2010 作图。
2 结果与分析
2.1 芥菜TCP家族成员鉴定
通过对芥菜全基因组数据库进行检索,并利用Pfam 工具鉴定结构域,最终获得53 条具有典型TCP结构域的序列,分别命名为BjuTCP1~BjuTCP53。根据序列信息进行理化性质分析,基因名称、登录号、氨基酸大小、分子量、理论等电点、染色体定位和亚细胞定位等特征见表1。结果显示,这些TCP 基因的氨基酸大小、分子量及等电点呈现出较大差异:氨基酸长度最短的是BjuTCP25 基因编码的蛋白,只含有216 个氨基酸,其蛋白分子质量为23 116.85 Da,氨基酸最长的是BjuTCP31 蛋白,含有437 个氨基酸,其蛋白分子质量为50 079.68 Da,理论等电点预测为5.50~10.18。根据亚细胞定位分析,绝大部分基因主要定位于细胞核,和已报道的其他物种中TCP 转录因子基因类似。

(续表1)
2.2 芥菜TCP家族系统进化树及聚类分析
为了分析TCP 转录因子家族基因的进化关系,选用模式植物拟南芥、水稻与芥菜一起构建系统进化树。在TCP 家族进化树中,53 个芥菜TCP、24 个拟南芥TCP 和21 个水稻TCP 呈现明显的聚类现象,如图1 所示,TCP 家族可清楚地划分为2 个大的亚家族,其中Class Ⅱ还可以细分为2 个小组,根据前人的分类方法,分别命名为:Class Ⅰ(PCF)、ClassⅡ(CIN)和 Class Ⅱ(CYC/TB1)。Class Ⅰ(PCF)包含的成员数目最多,其中芥菜Class Ⅰ(PCF)包含30个基因,水稻包含10 个,拟南芥包含13 个;其次是Class Ⅱ(CIN)亚组,包含 12 个芥菜基因、8 个水稻、8 个拟南芥;Class Ⅱ(CYC/TB1)成员数目最少,芥菜有11 个,水稻有3 个,拟南芥有3 个。
2.3 芥菜TCP家族基因结构及保守基序分析
TCP 家族成员的基因结构分析显示,44 个成员具有外显子结构,占全部基因的83%,仅有9 个基因含有1~3 个内含子,其中内含子序列较长的是BjuTCP32,达 600 bp 左右(图 2)。芥菜与番茄[16]、黄瓜[17]和柑橘[18]TCP 基因所含内含子数量所占比例类似,表明TCP 转录因子家族基因的结构和进化在物种间相对保守。

图1 芥菜、拟南芥和水稻TCP 基因系统进化树图Figure 1 Phylogenetic tree of all TCP genes in B.juncea,Arabidopsis and Oryza sativa
利用MEME 网站对芥菜TCP 家族进行基因序列的保守基序分析,所得结果见图2 和表2。分析结果发现芥菜所有基因都含有TCP 家族中最基本的motif 1,表2 横线处为motif 1 中氨基酸同源性大于90%区域,说明motif 1 是TCP 家族最重要的保守结构。TCP 家族不同组内保守基序的分布有较大差异,ClassⅠ组的大部分基因的保守基序数量明显比Class Ⅱ组的高。ClassⅠ组全部含有motif 1 和motif 2,且排列顺序都一样,说明这2 个motif 在ClassⅠ组内非常的保守;此外,一部分TCP 蛋白含有特有的基序,如 BjuTCP01 ~06 特异性含有 motif 9,BjuTCP07~20 特异性含有 motif 14,BjuTCP21~20 特异性含有motif 5 和motif 8,这些特异性的基序结构,可能与不同成员参与一些特定的调控过程有关。对于Class Ⅱ组来说,CYC/TB1 组中均含有motif 13,而 CIN 亚组不含有 motif 13 但含有 motif 6,且CIN 含有的基序模块明显少于CYC/TB1,说明两个亚组存在显著的模块差异,可能影响两个亚组成员的功能。整体看来,大部分位于同一亚族的基因具有相似的保守基序,并表现出一定的排列规律,与前面的系统进化树的分类相吻合,不同亚族成员间的差异也为研究TCP 家族中不同亚族的进化提供了参考。
2.4 芥菜TCP家族基因染色体定位分析
在所筛选出的53 个TCP基因中,除了BjuTCP46尚未确定染色体位置,其他基因都有明确的染色体信息(表1)。本研究利用MapInspect 软件绘制出剩余52 个TCP 基因家族成员在染色体上的分布情况,如图3 所示。结果发现仅A01 和A04 染色体没有基因分布,其余16 条染色体上基因分布位置不均匀,且B 类亚基因组中染色体分布的基因比A 类亚基因组多。A 类亚基因组染色体共含有24 个基因,占46%;B 类亚基因组含有基因28 个,占54%。一些TCP 基因在染色体上聚集在相近或相同的区域,表现出成簇的现象,例如A03 染色体上的BjuTCP45 和 BjuTCP22、A07 染色体上的下端 4 个基因、B07 染色体上的 BjuTCP31 和 BjuTCP32、BjuTCP30 和 BjuTCP43 等。
对染色体上基因数量的分布情况进行统计(图4),发现只有A03、B03 和B07 染色体分布有不同亚组的TCP 基因;A02 和B03 染色体上基因数目最多,达到6 个,但B03 分布种类更加丰富;A08、A10、B01 和B06 等4 条染色体只分布有 1 个基因,大都位于染色体下端。
2.5 芥菜TCP家族基因表达分析
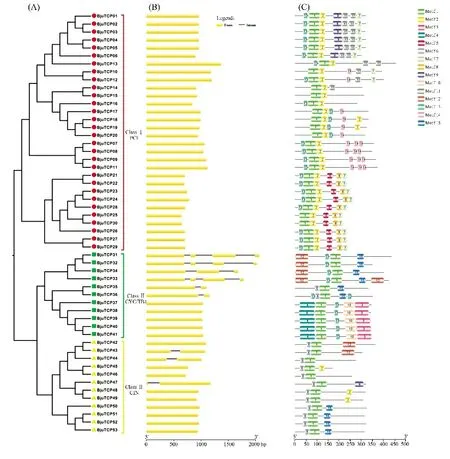
图2 芥菜TCP 家族成员基因结构分析图Figure 2 Gene structure analysis of TCP family members in B.juncea

表2 芥菜TCP 蛋白基序信息Table 2 Motif information of TCP family proteins in B.juncea

图3 TCP 家族成员染色体定位信息Figure 3 Chromosomal localization of TCP family members

图4 芥菜各染色体TCP 基因分布数量Figure 4 Number of TCP genes in each chromosomes
根据芥菜瘤茎膨大不同时期的转录组数据,对TCP 家族基因的表达模式进行分析。通过比较不同发育阶段转录组数据中的表达丰度,除BjuTCP31、BjuTCP43、BjuTCP44、BjuTCP45 和 BjuTCP48 外,剩下的48 个基因都检测到了相应的表达量,基因的表达模式如图5 所示。芥菜TCP 基因的表达模式在不同亚组内具有较高的相似性,可以按亚组分成两类:ClassⅠ和ClassⅡ。ClassⅠ亚组基因整体呈现出较高的表达水平,推测Class Ⅰ亚组成员在茎发育中发挥重要的作用;其中ClassⅠ亚组中,BjuTCP02、BjuTCP03、BjuTCP04、BjuTCP05 和 BjuTCP06 在前 2个阶段表达量较高,后两个阶段明显下降,推测这几个基因参与了芥菜茎前期发育;ClassⅡ亚组成员整体表达量相对较低,除了BjuTCP39 外,其余基因表达量始终维持在较低的水平。在其他物种TCP 基因的功能研究中,发现Class Ⅰ亚组成员广泛参与了植物的生长发育,OsTCP7 基因在水稻分蘖期的表达量最高[19],TCP15 与TCP22 之间可能存在功能冗余并共同调节叶片的衰老死亡过程[20]。芥菜中大量的TCP 家族基因在茎发育阶段的高表达,对茎瘤芥产品器官的正常发育起了重要的作用。
2.6 不同物种TCP基因数目比较

图5 芥菜TCP 基因在茎不同发育时期的表达模式热图Figure 5 Expression pattern of TCP genes in B.juncea at different stages of stem development
随着分子生物学和生物信息技术的发展,TCP家族基因相继在番茄、黄瓜、柑橘、葡萄以及砀山酥梨[16-22]等多种物种中得到鉴定。为了比较不同物种间TCP 家族成员的构成,我们检索了14 种植物的TCP 家族基因成员,各物种TCP 基因数量的统计情况见表3。结果显示,大部分物种Class Ⅱ含有的基因数量整体大于Class Ⅰ;除了苹果、番茄和葡萄外,PCF 亚组含有基因数量最多,CIN 亚组次之,CYC/TB1 亚组含量最少,但玉米中不含有CIN 亚组成员。虽然在不同物种中各个亚组基因数目存在差异,但几乎每个亚族所占比例较为一致,这些结果均揭示了TCP 家族在物种间的进化具有较高的保守性。双子叶植物明显比单子叶植物含有的TCP 基因数量更多,可能与基因组的进化关系有关,推测TCP 家族在双子叶植物的演化过程中发生了大规模的扩增;在双子叶植物中,TCP 基因数量从15~78之间不等,各科各属之间的TCP 家族基因数量存在着较大差异,蔷薇科的梨和苹果基因数量明显大于葫芦科的黄瓜和西瓜;与芥菜同属于芸薹属的其他植物含有TCP 家族成员基因数量也较大,以异源四倍体的甘蓝型油菜含有基因数量最多,几乎是白菜和甘蓝的2 倍,大致符合禹氏三角理论。
3 讨论与结论
TCP 转录因子是植物特有的一类转录因子家族,参与植物生长发育中的多种生理过程[7]。随着分子生物学技术的发展普及,TCP 转录因子家族基因相继在拟南芥、水稻、番茄、黄瓜、苹果及葡萄等物种得到鉴定。在拟南芥和水稻基因组中分别发现24和 21 个 TCP 基因[23],黄瓜中鉴定 22 个[17],葡萄中鉴定 15 个[21],番茄中鉴定 36 个[24],苹果中鉴定 52 个[25]。但芥菜TCP 家族基因成员还未见报道。本研究采用生物信息学的方法对芥菜全基因组数据库进行检索,共鉴定出53 个芥菜TCP 转录因子家族基因成员,并利用生物信息学方法对其进行较为系统全面的分析。与不同物种的TCP 基因数量进行对比(表3),各物种间有较大差异,甘蓝型油菜TCP 基因数量含量多,含78 个,芥菜次之。

表3 不同物种TCP 基因数目比较Table 3 The number of TCP genes in different species
根据序列基本特征分析显示,这些基因序列长度、氨基酸大小、分子量和等电点差异较大,且多编码碱性氨基酸,位于细胞核上;系统进化树分析把芥菜的53 个家族基因分为两大类:ClassⅠ(PCF)和Class Ⅱ(CIN),又称(CYC/TB1);基因结构分析表明芥菜TCP 基因结构较为简单,大部分基因没有内含子,暗示了家族序列具有较高的保守性;保守基序分析结果与进化分析相匹配,各亚组内部保守基序有很强的特异含有性;染色体定位分析表明TCP 基因并不完全分配在芥菜的全部18 条染色体上,且呈现分布不均的现象,B 亚基因组含有的TCP 基因多于A 亚基因组,基因在染色体上呈现成簇或分散的现象,这可能与基因复制相关。
本研究进一步利用芥菜转录组数据,分析了TCP 基因在芥菜茎膨大时期的表达模式。基因表达分析结果也表现出聚类现象,基于进化分析,可将结果按表达量的相似性分为Class Ⅰ和Class Ⅱ两类。Class Ⅰ(PCF)大部分成员显示出较高的表达量,Class Ⅱ成员表达水平相对较低。这说明同一亚组的基因,功能上可能也存在较大的相似性。研究发现,TCP 转录因子家族在植物整个生长周期中参与多种生物学过程的调控,尤其与生长发育密切相关。Class Ⅰ(PCF)基因表达范围广泛,尤其在含有大量分生细胞的组织器官中特异性表达[5]。拟南芥AtTCP14 和AtTCP15 在幼穗茎、幼花花梗、子叶和叶原基节间表达,参与调节叶片叶形发育,通过促进细胞增殖来调节节间发育[26]。J.M.Davière 等[27]发现GA 激素参与TCP 基因对株高的调控过程,双突变体 AtTCP14 和 AtTCP15 会导致植物矮化;西瓜ClTCP14a 和ClTCP15 也有相似功能,通过与激素的相互作用参与植株的生长发育[28]。茎瘤芥的主要产品器官是其肉质瘤状茎,茎的膨大涉及大量细胞的增殖分化,有关基因参与茎膨大的转录调控研究目前还较少报道。芥菜Class Ⅰ(PCF)基因在茎发育时期显著表达,尤其是一些基因(如BjuTCP02、BjuTCP 03、BjuTCP04、BjuTCP05 和 BjuTCP06)显示出明显的阶段表达特点,推测该组成员在茎发育中起重要作用。Class Ⅱ由CYC/TB1 和 CIN 两个亚组组成,对该类成员的功能研究较多。金鱼草CYC 是第1 个被鉴定出调控不对称花发育的TCP 基因[3];TB1 基因是侧枝发育的负调控元件,在拟南芥、玉米、水稻和矮牵牛中,该基因通过控制腋芽的生长来影响分蘖数,其缺失突变体均表现为腋芽增长侧枝增多[29-32]。CIN 亚组基因受到 miRNA319 调控,与叶片发育、叶片形态建成有关[33-36]。AtTCP2 和AtTCP17与植物光合作用有关,参与光信号传导途径[37-38]。在芥菜茎发育中,Class Ⅱ组成员表达量水平整体较低,可能未直接参与茎的膨大过程,但猜测可能在植物其他生长发育如叶的发育、花的发育中起作用。综合以上结果,我们发现芥菜的TCP 家族不同亚组成员对植物的调控存在一定的差异,各基因的具体功能还需要进一步的功能验证。本文基于全基因测序结果和转录组数据结果,对芥菜TCP 家族的成员进行了较为系统的鉴定和分析,为今后研究TCP 家族基因功能提供了理论依据,并为研究茎瘤芥生长发育奠定理论基础。
