青藏高原高寒草地物种多样性的海拔梯度格局及其对模拟增温的响应
2019-09-10温静张世雄杨晓艳秦瑞敏徐满厚
温静 张世雄 杨晓艳 秦瑞敏 徐满厚








摘要:为了研究青藏高原高寒植被生态系统的空间分布格局及其对全球气候变化的响应,以青藏高原高寒草地为研究对象,选取8个不同海拔梯度的典型试验区,每个试验区采取对照和增温2种处理。结果表明:(1)海拔梯度<4000 m的高寒草地禾本科为主要优势科,海拔梯度>4000 m的高寒草地莎草科为主要优势科;(2)中等海拔梯度的高寒草地物种多样性相对较高,低海拔地区的高寒草地物种多样性高于高海拔地区;(3)主动增温和被动增温对青藏高原高寒草地物种多样性产生不同影响,但整体上增温降低了物种多样性,且群落中优势科(禾本科和莎草科)对增温的响应更为敏感。因此,增温和海拔梯度对青藏高原高寒草地物种多样性产生重要影响,两者都降低了物种多样性。
关键词:高寒草地;物种多样性;增温;海拔;青藏高原
中图分类号:Q948.11 文献标志码:A 论文编号:cjas19030018
0引言
IPCC第五次评估报告指出,1980—2010年可能是有器测记录以来最热的30年,近10年(2000—2010年)温度达到最高点。气温升高会影响植物群落的结构和功能,甚至改变植物的空间分布格局,植物对全球变暖的响应研究正在成为新的热点领域和前沿课题。
温度控制着生态系统中许多生物化学反应过程的速率,几乎影响所有的生态学过程。全球变暖将影响到草地生态系统的结构和功能,一方面增温可以改变高寒植被的光合能力、生长速率及呼吸等生理过程,从而改变植物的物候[7]、生长期等;另一方面温度升高引起土壤含水率和营养物质利用效率的改变,会导致植物的生长、生物量生产分配及植物群落结构发生改变,甚至导致群落演替发生。
海拔梯度包含了温度、湿度和太阳辐射等环境因子,影响物种多样性的分布规律。高山地区植物群落物种多样性沿海拔梯度的变化表现为5种模式:负相关、中间高度膨胀、中等海拔高度较低、正相关和无关。关于物种多样性沿海拔的分布规律还未得出一致的结论,目前的研究中,中间高度膨胀和负相关2种模式比较普遍。
青藏高原是中国最大的高原,同时也是世界海拔最高的高原,作为全球气候变化的敏感区域,其草地生态系统面积巨大且极其脆弱。高寒草地是在高原特殊气候环境下发育的典型的植被类型,其生态系统十分脆弱,对气候变化和人类活动的响应强烈,破坏后不仅很难在短时间内恢复,甚至可能导致草场退化及沙漠化。因此,在全球变暖的大背景下,研究青藏高原高寒草地对气候变化的响应具有重要意义。
由于青藏高原多年冻土环境比较恶劣,在不同海拔设置试验点困难较大,目前相关的研究大部分是在1个试验点进行,对于高寒草地物种多样性与海拔梯度的相关关系研究较少。所以,本研究主要從不同海拔梯度的高寒草地对模拟增温的响应进行分析研究,以期探讨增温和海拔梯度对青藏高原高寒草地物种多样性的影响。
1研究方法
1.1研究区概况
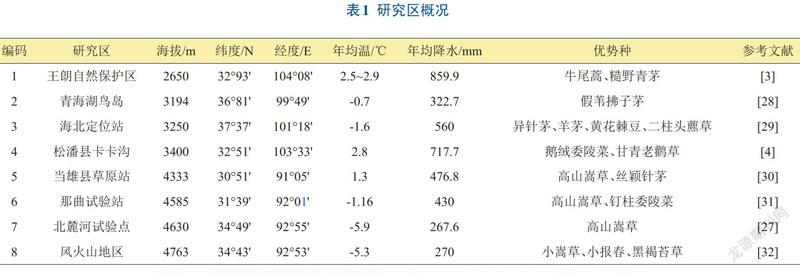
研究区位于青藏高原,属于高原大陆性气候,年平均气温在-5.9℃〜2.9℃之间,降水量在267.6〜859.9 mm左右,且大部分集中在生长季。分别从海拔2650〜4763 m之间选取8个试验点(见表1和图1),对高寒草地在常温和增温条件下的物种多样性变化特征进行分析。8个试验点位于30°51'N—37°37'N,91°05'E—104°08'E之间,高原腹地和边缘均有分布。
1.2数据来源
本研究通过对相关文献进行整理和数据筛选,并结合自己研究区(北麓河试验点)的试验数据进行综合分析。所采用的试验数据年份主要集中于2006—2014年之间,利用物种的重要值(IV)和丰富度指数(S)进行物种多样性的计算。
1.3增温样地设置
每个研究区均进行对照(CK)和增温(W)2种处理,增温方法除研究区7采用红外辐射增温的方法外,都采用国际冻原计划(ITEX)中的开顶箱(Open-top chambers,OTCs)模拟增温试验。红外线辐射器(Infrared radiation,IR)需要电力供应,为主动增温装置,其增温原理为模拟太阳辐射,通过红外线辐射对辐射区域进行加热(图2);OTCs无需电力供应,材料为透明的有机玻璃,上端开口,为被动增温装置,其增温原理为通过小室降低风速,由于空气湍流强度减弱使小室内的热量不易散失而达到增温的效果(图3)。各研究区的增温效果,见表2。
1.4数据处理
重要值(IV)计算,见公式(1)。
IV=(相对盖度+相对高度+相对频度)/3………………(1)
选用的多样性指标包括Margalef丰富度指数、Simpson指数、Shannon-Weiner指数和Pielou均匀度指数,计算分别见公式(2)〜(5)。其中:R为Margalef丰富度指数,H为Simpson指数,H'为Shannon-Weiner指数,E为Pielou均匀度指数,i是样方内的植物物种,S是样方内所有物种种类之和,p为是物种i的重要值。
1.5统计分析
数据处理与统计图绘制由Office 2016完成,多样性指数与温度、海拔的显著性检验由SPSS 19.0软件进行方差分析得到。
2结果与分析
2.1优势科随增温与海拔的变化情况
通过对8个研究区植物群落构成的整理,将其划分为禾本科、菊科、莎草科、豆科、蔷薇科和其他科6大功能群(见表3)。
在海拔梯度<4000 m的研究区,特别是在2650 m、3194 m和3250 m海拔处,禾本科植物重要值较其他功能群高出大约1倍以上,是海拔梯度<4000 m的植物群落中的优势科;蔷薇科类植物是海拔3400 m研究区的绝对优势科,其重要值远远高于本海拔的其他功能群;莎草科类植物在低海拔梯度上的分布较少,仅在海拔3250m的研究区出现。在增温作用下,海拔3250m的禾本科的重要值略微下降但不明显;2650 m、3194 m和3400 m海拔处研究区的禾本科类植物的重要值都有提高;蔷薇科作为海拔3400 m处的优势科在增温作用下重要值大幅度下降,但仍在群落中保持着绝对的优势。
在海拔梯度>4000 m的研究区,特别是在海拔4333 m、4585 m和4754 m的研究区,莎草科重要值最高,在海拔4630 m处,莎草科植物重要值略低于豆科植物,但在整个植物群落中,其重要值仍然较高,所以在海拔梯度>4000 m的研究区,莎草科类植物是其群落的优势科。在增温处理下,不同研究区植物群落中各功能群重要值变化情况各异。具体而言,海拔4333 m处蔷薇科和杂草的重要值增加,其他功能群重要值均下降;海拔4585 m处禾本科和莎草科重要值下降,其他功能群的重要值均增加;海拔4630 m处豆科和蔷薇科的重要值减少,其他功能群的重要值没有发生明显变化;海拔4754 m处杂草重要值下降,其他功能群重要值均增加。
综合8个海拔梯度的不同功能群所占的重要值,发现位于低海拔(2650 m、3194 m)和高海拔(4754 m)的研究区,其优势科在增温条件下重要值增加,中间海拔的研究区,在增温条件下其优势科的重要值均下降。对比各海拔增温前后优势科的重要值变化可以发现,大部分优势科受温度影响其重要值变化幅度相对较大,说明优势科对温度的响应相对敏感。
2.2自然条件下海拔梯度对物种多样性的影响
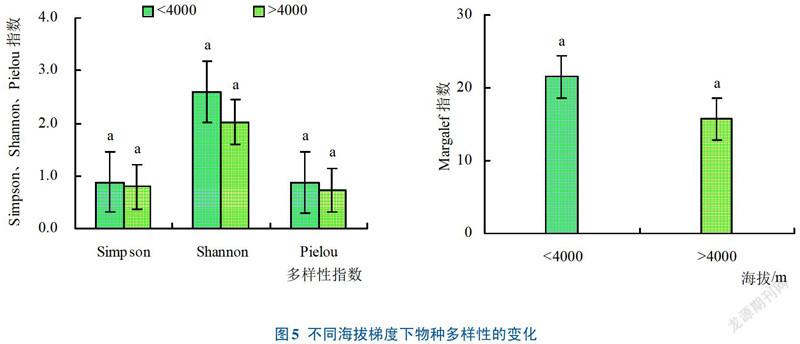
在自然条件下,海拔4585 m、3250 m、3400 m和2650 m的Simpson指数相对较高;海拔3250 m、4333 m、3400 m和4630 m的Shannon指数较高;除海拔4585 m的研究区外,其他研究区的Pielou均匀度都比较高;Margalef指数较高的研究区主要集中于海拔为3250 m、3400 m和4630 m的中等海拔地区。整体而言,海拔3250〜4333 m研究区的Simpson指数、Shannon指数、Pielou指数、Margalef指数都相对较高(图4),说明中等海拔梯度的物种多样性相对较丰富,而低海拔地区(2650 m、3194 m)和高海拔地区(4754 m)物种多样性的4个指标相对较低,且低海拔地区的物种多样性指标高于高海拔地区。将8个研究区划分为低海拔<4000 m和高海拔>4000 m个梯度(见图5),选择各研究区的对照样地作为研究海拔与物种多样性的基础数据,发现海拔<4000 m的研究区4个物种多样性指数值均高于海拔>4000m的研究区(P>0.05),说明低海拔地区的物种多样性大于高海拔地区。
2.3不同海拔梯度下增温对物种多样性的影响
从物种多样性指标随海拔的变化可以看出(见图4),增温处理下,低海拔地区(2650 m、3194 m)和高海拔地区(4754 m)物种多样性指标均不同程度的下降;海拔4585 m的研究区物种多样性指数都较其他研究区低,但在增温处理下,其各项指标均增加;中等海拔的其他研究区各物种多样性指数变化没有明显的规律。具体而言,Simpson指数在海拔3400〜4630 m之间的研究区增温后都有所增加,特别是海拔4333 m的研究区增加情况最为明显,其余4个研究区的Simpson指数在增温后都下降,下降最明显的研究区为海拔4754 m的地区。Shannon指数和Pielou指数在增温处理下,除海拔3250 m和4585 m的研究区增加外其余均减少,其中海拔4333 m研究区下降最为明显。Margalef指数在增温处理下,2650 m、4333 m和4763 m海拔处下降,在3194 m、3250 m、3400 m和4630 m海拔处没有发生变化,在海拔4585 m处略有增大。对比8个研究区增温样地与对照样地的物种多样性发现,Simpson指数在增温处理下整体略微上升(P>0.05),Shannon指数、Pielou指数和Margalef指数在增温情况下整体下降(P>0.05)(见图6)。因此,增温整体上减少了物种多样性。
3讨论
生物多样性沿环境梯度的变化规律是生物多样性研究的一个重要议题,而物种多样性是最简单有效的描述群落和区域多样性的方法,是生物多样性的本质内容。草地生态系统作为陆地生态系统的重要组成部分,其群落结构相对单一,干扰因素相对较少,为物种多样性研究提供了理想对象。温度作为影响植物生长的重要因素,特别是在青藏高原这个特殊的地理单元上,从影响温度的人为(模拟增温)和自然(海拔梯度)2个方面探讨其对物种多样性的影响对于研究全球气候变化的响应有一定意义。
3.1物种多样性对增温的响应
增温时间长短影响高寒草地的物种多样性,本研究中的8个研究区增温时长不一,可大体分为1年短期模拟增温和3、4年长期模拟增温。其中短期模拟增温影响下,除个别研究区物种数量下降,其余物种多样性指标整体变化都不明显,说明短期增温对物种多样性的影响较小。在长期增温条件下,物种多样性指标变化相对短期增温明显,且除Simpson指数外其余指数都下降,总的来说长期增温会导致物种多样性的下降。牛书丽等认為生态系统不同组分和过程对温度的敏感性不同,短期的增温试验往往不能得出作为整体的生态系统响应与适应全球变暖的正确结论,而长期增温植物受水分、养分等环境因子的改变的影响生长受到限制。
增溫装置分为开顶箱和红外线辐射器2种。开顶箱在青藏高原高寒草地的研究中应用较为普遍,是最经济、最简单的增温装置,但对于气候变化的模拟并不充分,本研究选取的研究区中有7个采用此增温装置。红外线辐射器能够模拟全球变暖的增温机制和日变化,且对土壤及植被无物理干扰,但其耗费电力较多,本研究中有1个研究区采用此增温装置。通过对比2种增温装置对物种多样性指数的影响,认为2种增温装置对物种多样性产生不同的影响。其中被动增温增温幅度较小,物种多样性增加,而主动增温的增温幅度较大,物种多样性下降,说明在一定增温范围内增温有助于物种多样性的增加,超过一定范围物种多样性会下降,这与李英年等的结论一致。Rustad等通过Meta-analysis的方法将不同地点、不同植被类型的试验结果平均起来进行比较,认为增温装置对土壤呼吸及植物生长对增温的反应等的影响不显著。而牛书丽等认为,不同增温装置的加热机制不同,可能会引起陆地植物和生态系统特异性的反应。目前中国的增温试验大都使用不同规格的增温装置试验于不同的生态系统中,加大了试验结果整合分析的难度和模型预测的不确定性,这也是本研究遇到的最棘手的问题。
综合增温时间和增温装置两者对高寒草地物种多样性的影响,得出增温使青藏高原高寒草地的物种多样性降低的结论。该结论与李英年等的结论一致。李英年等的研究表明:经过5年的增温试验,矮嵩草草地物种多样性比对照样地的物种多样性有所减少。徐满厚等的研究表明:增温下物种多样性略有增加,但是响应不敏感。这与本研究的结论不一致,这可能与研究区域有关,徐满厚等的研究区年平均气温为-5.9℃,受低温胁迫严重,模拟增温刚好缓解了这种胁迫,使得生境得到改善,物种多样性增加。本研究选取8个研究区研究物种多样性对增温的响应,并综合对研究区的物种多样性指数进行分析,一定程度上减少了个别因素对于整个研究结果的影响。
3.2物种多样性对海拔的响应
本研究表明,中间海拔梯度的物种多样性相对较高,低海拔梯度的物种多样性高于高海拔地区的。刘哲等的研究表明:2条样带不同海拔梯度高寒草地植物群落的Shannon指数、Simpson指数、Margalef的最大值均出现于中间海拔梯度,说明在同一座山体上中间海拔梯度有较高的物种多样性。这与本研究结论大体上相一致,也符合贺金生总结的陆地物种多样性的海拔梯度变化模式中的中间高度膨胀模式。关于物种多样性随海拔单峰分布格局的原因,可能是人为过度干扰、水热环境与物种竞争能力三重因素共同作用形成的:低海拔地区,环境相对优越,竞争力强的物种会在群落中占据优势,但人类活动的过度干扰导致某些物种的丧失,所以物种多样性低;高海拔地区,水热条件比低海拔地区恶劣,物种的适应能力和竞争能力弱会导致物种的消失,所以其物种多样性低;而中海拔梯度,人为干扰较低海拔地区少,水热条件较高海拔地区好,所以其物种多样性最高。
4结论
本研究主要从增温和海拔梯度2个方面对青藏高原高寒草地的物种多样性进行分析研究,发现禾本科为海拔梯度<4000 m的主要优势科,莎草科为海拔>4000 m的主要优势科。青藏高原高寒草地的物种多样性受到增温和海拔梯度变化的影响,增温降低高寒草地群落物种多样性,随着海拔升高,物种多样性减小。主动增温和被动增温2种增温方式都增温降低了物种多样性,且群落中优势科(禾本科和莎草科)对增温的响应更为敏感。
然而,由于文章数据大部分来源于整理他人文献所得,每个试验样地选址、增温装置、试验时间和试验目的等都有所相同,这些因素都或多或少对试验结果产生一定影响。故此,今后需探索一种更为合理的方法对增温处理下物种多样性沿海拔梯度的响应变化进行深入研究。
参考文献
[1]IPCC.IPCC fifth Assessment Report(AR5)[M].Cambridge:Cambridge University Press,2013.
[2]贠汉伯,吴青柏,芮鹏飞,等.适用于多年冻土区具有碳通量自动观测性能的OTC系统开发设计[J].冰川冻土,2015,37(2):454-460.
[3]徐振锋,胡庭兴,李小艳,等.川西亚高山采伐迹地草坡群落对模拟增温的短期响应[J].生态学报,2009,29(6):2899-2905.
[4]石福孙,吴宁,罗鹏.川西北亚高山草甸植物群落结构及生物量对温度升高的响应[J].生态学报,2008(11):5286-5293.
[5]Klanderud K,Totland O.Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hot spot[J].Ecology,2005,86(8):2047-2054.
[6]珊丹,韩国栋,赵萌莉,等.控制性增温和施氮对荒漠草原土壤呼吸的影响[J].干旱区资源与环境,2009,23(9):106-112.
[7]徐满厚,薛娴.气候变暖对高寒地区植物生长与物候影响分析[J].干旱区资源与环境,2013,27(3):137-141.
[8]Walther G R,Beissner S,Burga C A.Trends in the upward shift of alpine plants[J].Journal of Vegetation Science,2005,16(5):541-548.
[9]Carlyle CN,Fraser LH,Turkington R.Response of grass-land biomass production to simulated climate change and clipping along an elevation gradient[J].0ecologia,2014,174:1065-1073.
[10]余欣超,姚步青,周华坤,等.青藏高原两种高寒草甸地下生量及其碳分配对长期增温的响应差异[J].科学通报,2015,60(4):379-388.
[11]Xu MH,Peng F,You QG,et al.Year-round warming and autumnal clipping lead to downward transport of root biomass,carbon and total nitrogen in soil of an alpine meadow[J].Environmental and Experimental Botany,2015,109:54-62.
[12]林丽,张德罡,曹广民,等.高寒嵩草草甸植物群落数量特征对不同利用强度的短期响应[J].生态学报,2016.36(24):1-10.
[13]Gaston KJ.Global patterns in biodiversity[J],Nature,2000,405:220-227.
[14]贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997(01):93-101.
[15]Gentry A H.Changes in plant community diversity and floristic composition on environmental and geographical gradients[J].Annals of the Missouri Botanical Garden,1988,75:1-34.
[16]Whittaker R H.Vegetation of the Siskiyou Mountains,Oregon and Califomia[J],Ecological Monographs,1960,30:279-338.
[17]Peet R K.Forest vegetation of the Colorado,Front Range;Pattern of species diversity[J].Vegetatio,1978,37:65-78.
[18]Itow S.Species turnover and diversity patterns along an elevation broad-leaved forest coenocline[J].Journal of Vegetation Science,1991,2:477-484.
[19]卢训令,胡楠,丁圣彦,等.伏牛山自然保护区物种多样性分布格局[J].生态学报,2010,30(21):5790-5798.
[20]Daubenmire R,Daubenmire J B.Forest vegetation of eastern Washington and northern Idaho[J].Washington Agric.Expt.Sta.Tech.Bull.,1968,60:1-104.
[21]段敏杰,高清竹,郭亚奇,等.藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局[J].草业科学,2011,28(10):1845-1850.
[22]孔祥海,李振基.福建梅花山常绿阔叶林植物物种多样性及其海拔梯度格局[J].植物分类与资源学报,2012,34(02):179-186.
[23]牛常青,曲波,牛霞霞.乌金山植物群落物种多样性的垂直分布格局[J].晋中学院学报,2014,31(03):56-63.
[24]Baruch Z.Ordination and classification of vegetation along an altitudinal gradient in the Venezuelan paramos[J].Vegetation,1984,55:115-126.
[25]Wilson J B,Sydes M T.Some tests for niche limitation by examination of species diversity in the Dunedin area,New Zealand[J].N.Z.J.Bot.,1988,26:237-244.
[26]刘哲,李奇,陈懂懂,等.青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响[J].生物多样性,2015,23(04):451-462.
[27]徐满厚,刘敏,薛娴,等.增温、刈割对高寒草甸植被物种多样性和地下生物量的影响[J].生态学杂志,2015,34(09):2432-2439.
[28]陈骥,曹军骥,金钊,等.模拟增温对青海湖鸟岛高寒草原群落结构影响初步研究[J].干旱区资源与环境,2014,28(05):127-133.
[29]周华坤,周兴民,赵新全.模拟增温效应对矮嵩草草甸影响的初步研究[J].植物生态学报,2000(05):547-553.
[30]宗宁,柴曦,石培礼,等.藏北高寒草甸群落结构与物种组成对增温与施氮的响应[J].应用生态学报,2016,27(12):3739-3748.
[31]姜炎彬,范苗,张扬建.短期增温对藏北高寒草甸植物群落特征的影响[J].生态学杂志,2017,36(03):616-622.
[32]李娜,王根绪,杨燕,等.短期增温对青藏高原高寒草甸植物群落结构和生物量的影响[J].生态学报,2011,31(04):895-905.
[33]Kratochwil A.Biodiversity in Ecosystems:Principles and Case Studies of Different Complexity Levels[M].Springer,Netherlands,1999.
[34]Magurran AE.Ecological Diversity and Its Measurement[M].Princeton:Princeton University Press,1988.
[35]趙同谦,欧阳志云,贾良清,等.中国草地生态系统服务功能间接价值评价[J].生态学报,2004(06):1101-1110.
[36]牛书丽,韩兴国,马克平,等.全球变暖与陆地生态系统研究中的野外增温装置[J].植物生态学报,2007,31(02):262-271.
[37]李英年,赵亮,赵新全,等.5年模拟增温后矮嵩草草甸群落结构及生产量的变化[J].草地学报,2004,12(3):236-239.
[38]Rustad LE,Campbell JL,Marion GM,et al.A meta-analysis of the response of soil respiration,net nitrogen mineralization,and aboveground plant growth to experimental ecosystem warming[J].0ecologia,2001,126:543-562.
