堇菜叶片草酸钙晶体与水分维持的关系
2019-09-10苏志孟张习敏马琳段盛光陈婷乙引
苏志孟 张习敏 马琳 段盛光 陈婷 乙引

摘 要:随着全球气候变化加重,干旱强度和持续时间逐渐增加,严重影响植物生长和作物产量。喀斯特为典型的干旱和高钙生境,植物叶片富集大量的草酸钙晶体,而该晶体与植物耐旱性之间的关系并不清楚。该研究以喀斯特适生植物堇菜(Viola verecumda)为材料,土壤进行自然干旱,分析堇菜叶片的草酸钙晶体变化特征与水分之间的关系。结果表明:在土壤自然干旱条件下,堇菜主要通过细胞内束缚水的释放,维持细胞内水分平衡;而在干旱后期,叶片通过关闭气孔,将部分自由水转变为束缚水,防止水分流失。此外,草酸钙晶体的密度与束缚水含量具有极其显著的强正相关线性回归关系(r=0.825 3,P<0.000 1),表明草酸钙晶体作为主要的束缚水物质。因此,堇菜植物在耐旱过程中可能协调草酸钙晶体和气孔的生理行为忍耐干旱胁迫。
关键词:喀斯特地区, 抗旱, 叶水势, 束缚水, 晶体特征
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2019)06-0720-09
Abstract:As the global climate changes dramatically, the increasing intensity and duration of drought seriously affects plant growth and crop yield. Karst is a typical arid and high-calcium habitat, and plant leaves in karst areas contain a large amount of calcium oxalate crystals, while the relationship between the crystal and the drought tolerance of plants is not clear.
In this study, we took Viola verecumda grown in karst area as material, and analyzed the relationship between the changing characteristics of calcium oxalate cyrstals and water in its leaves, by subjecting the leaves to natural dry soil. The results showed that in the early stage of drought stress, V. verecumda maintained the intracellular water balance mainly through the release of intracellular bound water. In the later stage of drought, the leaves mainly prevented water loss by closing the stoma and converting part of the free water into bound water. In addition, the density of calcium oxalate crystals had an significant positive correlation with the bound water content (r=0.825 3, P<0.000 1), suggesting that calcium oxalate crystals act as the main matter binding water. Therefore, V. verecumda may coordinate the physiological behavior of calcium oxalate crystals and stomatal during drought tolerance.
Key words:karst area, drought resistance, leaf water potential, bound water, crystal characteristics
隨着全球气候变化和生态平衡失调,土地水资源利用短缺,干旱强度和持续时间逐渐增加,严重影响植物生长繁殖和作物产量。植物如何耐受干旱胁迫成为当今人们面临的重要问题。植物对干旱的适应可分为干旱避免、干旱耐受、干旱逃避和干旱恢复等方式(Fang & Xiong, 2015; Lawlor, 2013; Luo, 2010; Turner et al., 2001),涉及的主要机制分为植物保水、渗透调节和抗氧化胁迫(Fang & Xiong, 2015)。然而,这些机制主要是植物对短期水分胁迫的响应,在长期的干旱条件下,植物对干旱的适应性策略仍需进一步研究。
草酸钙是植物中最常见的生物矿物之一,广泛存在于藻类、低等维管束植物、裸子植物和被子植物中,被认为是植物生理代谢的惰性产物(Franceschi & Nakata, 2005; Macnish et al., 2003)。植物体内草酸钙晶体具有多种生理和生态学功能,如钙离子平衡(朱广龙等, 2014)、妨碍取食对象的取食(Ward et al., 1997; 乙引等, 2018)、解毒作用(Choi et al., 2001)以及光照的聚集和反射(Franceschi & Nakata, 2005)等。近年来,Tooulakou et al.(2016)研究发现,植物叶片被ABA诱导气孔闭合后,体内的草酸钙晶体进行分解形成二氧化碳参与光合作用。草酸钙晶体通常与水结合,它在植物体内的形成和降解可能影响植物体内的水分状态。然而,相关的研究报道却极少。
堇菜(Viola verecumda)是堇菜科堇菜属多年生草本植物,广泛分布在贵州喀斯特地区,全草可入药,具清热解毒、止咳、散瘀和抗菌的功效,多用于治疗肺热咳嗽、上呼吸道感染、毒蛇咬伤和刀伤出血等,被认为是一种天然抗生素(李彦连, 2008; 徐国兵等, 1995)。在中国西南贵州喀斯特地区(王德炉等, 2005),由于典型的喀斯特地质构造,土壤函水能力较差,植物经常遭受持续干旱胁迫(李渝等, 2009)。另外,堇菜植物叶片内富集大量草酸钙晶体,是研究喀斯特植物叶片内草酸钙与植物耐旱性关系的理想材料。因此,本研究以堇菜植物为材料,分析自然持续干旱胁迫下堇菜叶片水分参数、气孔导度和草酸钙晶体的变化特征以及干旱条件下堇菜叶片水分维持特征,为揭示喀斯特地区植物耐旱特征提供理論参考。
1 材料与方法
1.1 植物生长与自然干旱
堇菜(Viola verecumda)为喀斯特地区常见的多年生草本植物,能够在岩石缝隙中健康生长(图1)。本研究从贵州省贵阳市花溪区党武镇喀斯特山地选取长势相同、大小基本一致的一年生堇菜幼苗,移栽花盆中,每个花盆中种植4~5株植株。置于 25 ℃,16 h/8 h(昼/夜)光周期,相对湿度60%的培养室内培养。在生长期内,定期为植物浇水。待植株生长3个月后,选取长势相同、大小基本一致的10盆植株进行自然干旱处理。
1.2 自然干旱胁迫
在最后一次给土壤浇水饱和后,停止浇水。此后,不断监测土壤含水量,待土壤相对湿度在40%左右时,开始记为干旱胁迫(0 d),以后每隔2 d测定一次相关参数。
1.3 测量方法
1.3.1 土壤重量含水量和土壤相对湿度测量 在根周围取5 cm左右的土壤,测定其鲜重,并置于65 ℃烘箱烘干至恒重,称量土壤干重。土壤重量含水量计算公式:土壤重量含水量=(土壤鲜重-土壤干重)/土壤干重×100%,土壤相对湿度(RSWC)=土壤重量含水量/土壤持水量×100%。
1.3.2 叶片含水量和叶片相对含水量测量 叶片含水量用称重法,叶片相对含水量用饱和称重法。取堇菜新鲜离体叶片,称其鲜重,称重后立即浸入去离子水,4 ℃放置8 h后,称饱和质量,并置于105 ℃烘干箱中杀青1 h,80 ℃烘干至恒重,称其干重。叶片含水量=(叶片鲜重-叶片干重)/叶片鲜重×100%;叶片相对含水量=(叶片鲜重-叶片干重)/(叶片饱和鲜重-叶片干重)×100%。
1.3.3 叶水势测量 植物叶片水势采用WP4-T露点水势仪(WP4-T,Decagon,美国)测定。
1.3.4 水分饱和亏缺测量 水分饱和亏缺(WSD)采用饱和称重法,WSD=(叶片饱和鲜重-叶片鲜重)/(叶片饱和鲜重-叶片干重)×100%。
1.3.5 叶片自由水和束缚水测量 取适量待测植物叶片样品(1 g左右),剪成1~2 mm的小块后称重(Wy),并放入称量瓶中,加入6 mL 60%的蔗糖溶液:先测定蔗糖溶液旋光度,通过标准曲线计算蔗糖溶液质量百分比浓度(C1),称蔗糖溶液质量(W),密闭称量瓶,在黑暗环境中放置6 h,期间不停摇动,使叶片和蔗糖溶液充分接触。再测定蔗糖溶液的旋光度,通过标准曲线计算蔗糖溶液质量百分比浓度(C2)。蔗糖溶液旋光度使用阿贝氏折射仪测定。计算公式:叶片束缚水含量= 叶片含水量-叶片自由水含量。
叶片自由水含量 = W(C1-C2)C2Wy×100%。
1.3.6 叶片蒸腾速率和气孔导度测量 用Li-6400光合测定系统(LI-6400XT,LI-COR,美国)和红蓝光源叶室(6400-02B)测定蒸腾速率(Tr)和气孔导度(Gs)。测定时,光合有效辐射设置为400 μmol·m-2·s-1,流速为500 μmol·s-1,于上午9:00—11:00进行,每次随机选择5盆幼苗叶片测定。
1.3.7 草酸钙晶体特征 先随机取5株新鲜植物叶片,沿主脉剪为0.5 cm长,0.5 cm宽的小片,用3%的次氯酸钠溶液进行脱色,直至叶片变为白色,置于饱和水合氯醛溶液中放置24 h。然后于光学显微镜(Nikon 80i, Nikon, Japan)观察,拍照。采用Digimizer计算图片中草酸钙晶体的密度,随机测定50个独立晶体的直径。
1.3.8 草酸钙含量 先取适量的待测植物的叶片(约200 mg),放入研钵中,加入1 mL的2 mol·L-1 HCl,充分研磨,转入2 mL离心管中;再用1 mL 的2 mol·L-1 HCl清洗研钵和研棒,将清洗液全部转移至离心管中。于4 ℃ 10 000 r·min-1离心10 min,收集上清液。上清液经0.45 μm孔径滤膜过滤,采用安捷伦1200液相色谱系统(Agilent Technologies Inc. Santa Clara,USA)在215 nm测定其含量。测定条件:色谱柱[Carbomix H-NP 10:8%, 7.8 mm × 50 mm, 10 μm(Phenomenex, Torrance,CA, USA)];流动相为0.1% H3PO4;流速为0.6 mL·min-1;柱温为55 ℃;进样量为10 μL。采用上述相同方法,用草酸(购于sigma公司)作为标准,制作标准曲线,并根据标准曲线计算草酸含量。
1.4 数据处理
采用SPSS 16.0统计软件进行数据处理和统计分析,采用SigmaPlot 10.0软件作图。所有数据均为至少3次重复,值为平均值±标准差。
2 结果与分析
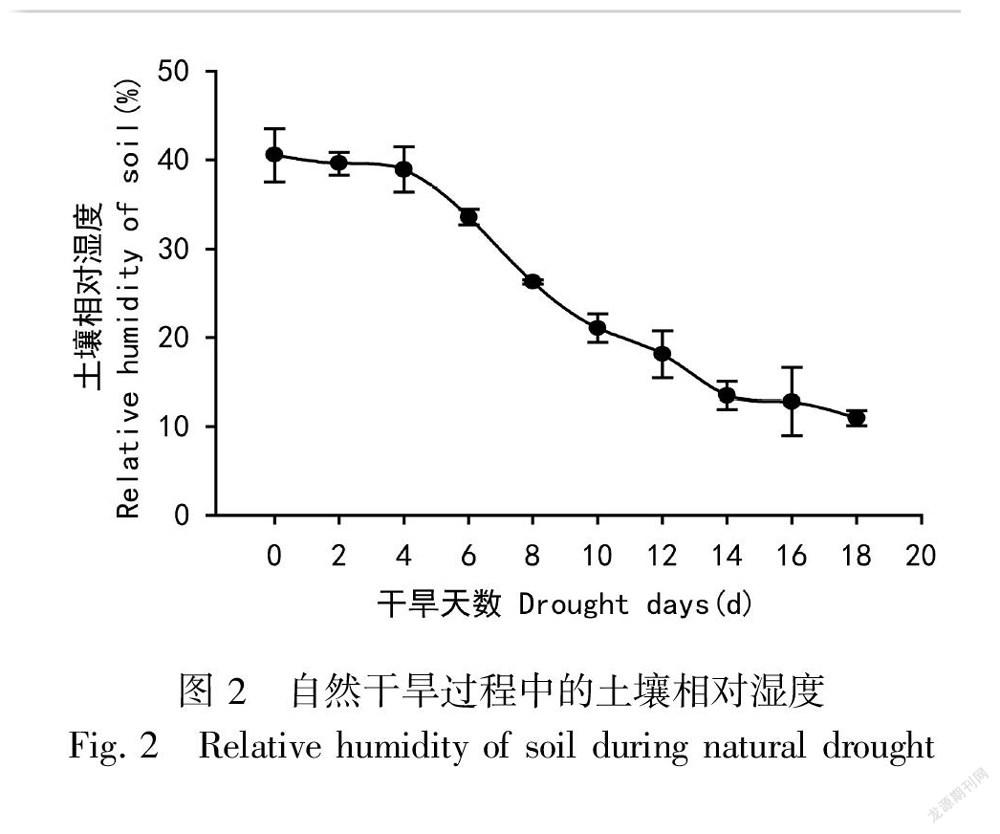
2.1 自然干旱下的土壤相对湿度
由图2可知,在土壤自然干旱过程中,干旱时间18 d,土壤相对湿度从40%左右降低到约10%。
从第0天到第4天,土壤的相对湿度下降较为缓慢,下降了1.60%;从第4天开始到第14天,土壤的相对湿度急剧下降,下降了25.46%;从第14天开始,土壤的相对湿度下降又趋于平缓,到第18天,共下降了2.49%。
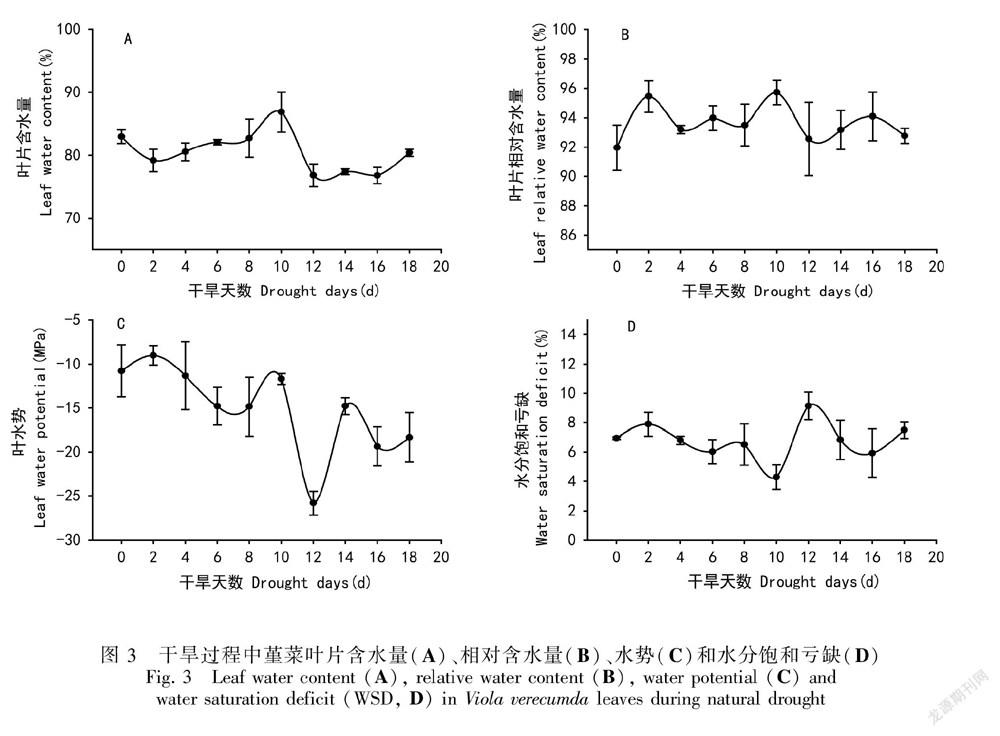
2.2 自然干旱下堇菜叶片含水量、相对含水量、水势和水分饱和亏损的变化
自然干旱过程中,堇菜叶片含水量基本维持在80%左右(图3:A)。同样,在自然干旱过程中,堇菜叶片相对含水量并没有出现显著的降低,一直维持在94%左右 (图3:B)。WSD不高于10%(图3:D)。
表明在干旱过程中,叶片水分流失较少,保水能力较强。另外,在干旱过程中,堇菜叶水势表现为缓慢的下降过程,在第0天到第10天的干旱期内,其叶片水势基本维持在-10 ~-15 MPa范围内;在第12天,水势出现急剧下降(-25.82 MPa);而后又升高到-20 MPa左右(图3:C)。结果表明,在干旱过程中,叶水势维持在较低水平,这有助于植物叶片水分的保持。
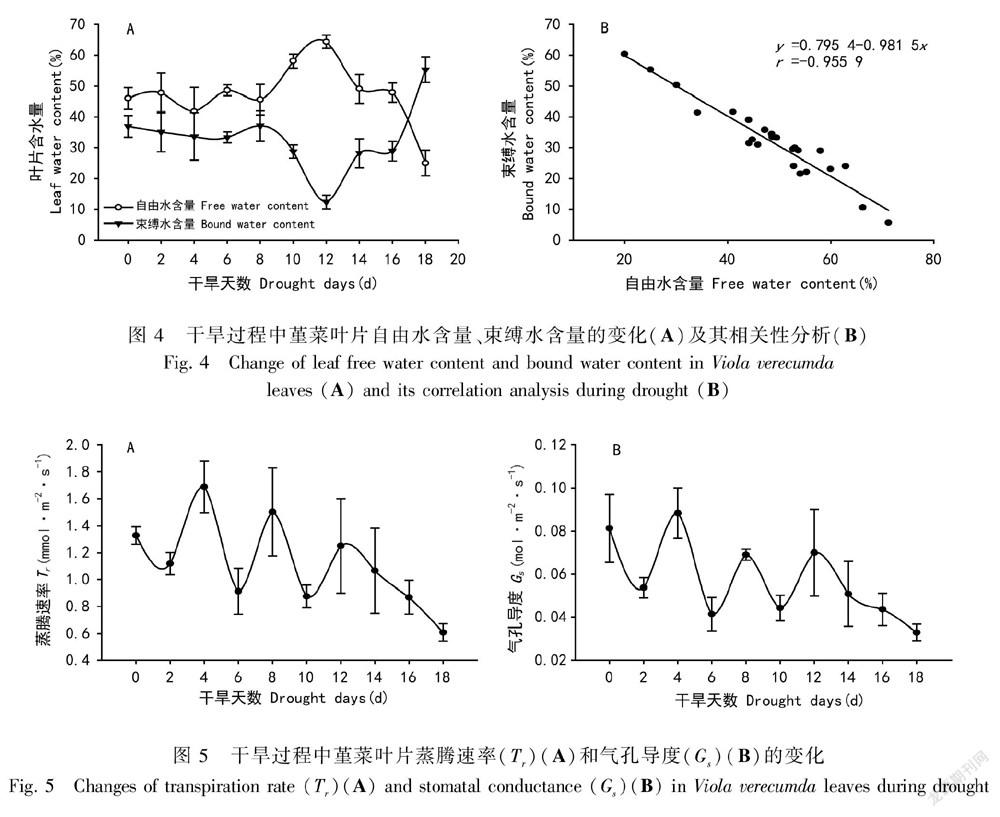
2.3 自然干旱下叶片自由水和束缚水含量的变化
植物细胞内存在自由水和束缚水,在植物干旱胁迫下,自由水易于丢失。图4结果表明,0~8 d叶片自由水和束缚水基本保持稳定,第8天叶片束缚水含量显著下降,第12天到达最低(12.35%),之后开始增加,第18天叶片束缚水含量急剧增加,值为55.39%,高于干旱前期的含量(图4:A);而自由水的变化趋势与束缚水却相反(图4:A)。整个干旱过程中,堇菜叶片自由水与束缚水具有极显著的负相关线性回归关系(r=0.955 9,P<0.000 1)(图4:B)。
2.4 自然干旱下叶片蒸腾速率和气孔导度的响应
为了分析干旱过程中,叶片的气孔生理行为,测定了堇菜叶片气孔导度和蒸腾速率的变化。图5结果显示,在干旱过程中,蒸腾速率(图5:A)与气孔导度(图5:B)在0~12 d呈现出动态波动的变化趋势,而在12~18 d两者总体呈现下降趋势。
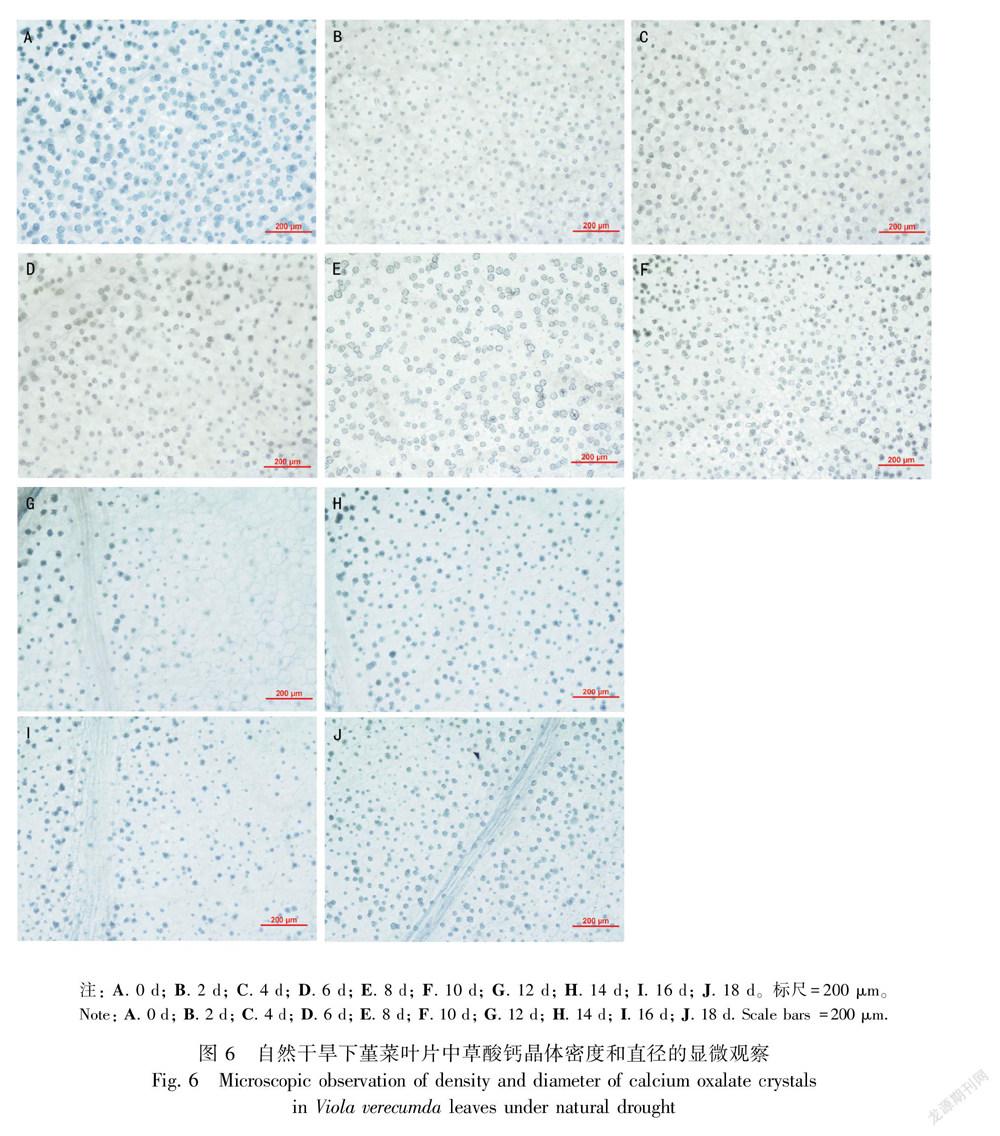
2.5 自然干旱过程中堇菜叶片中草酸钙晶体的变化特征
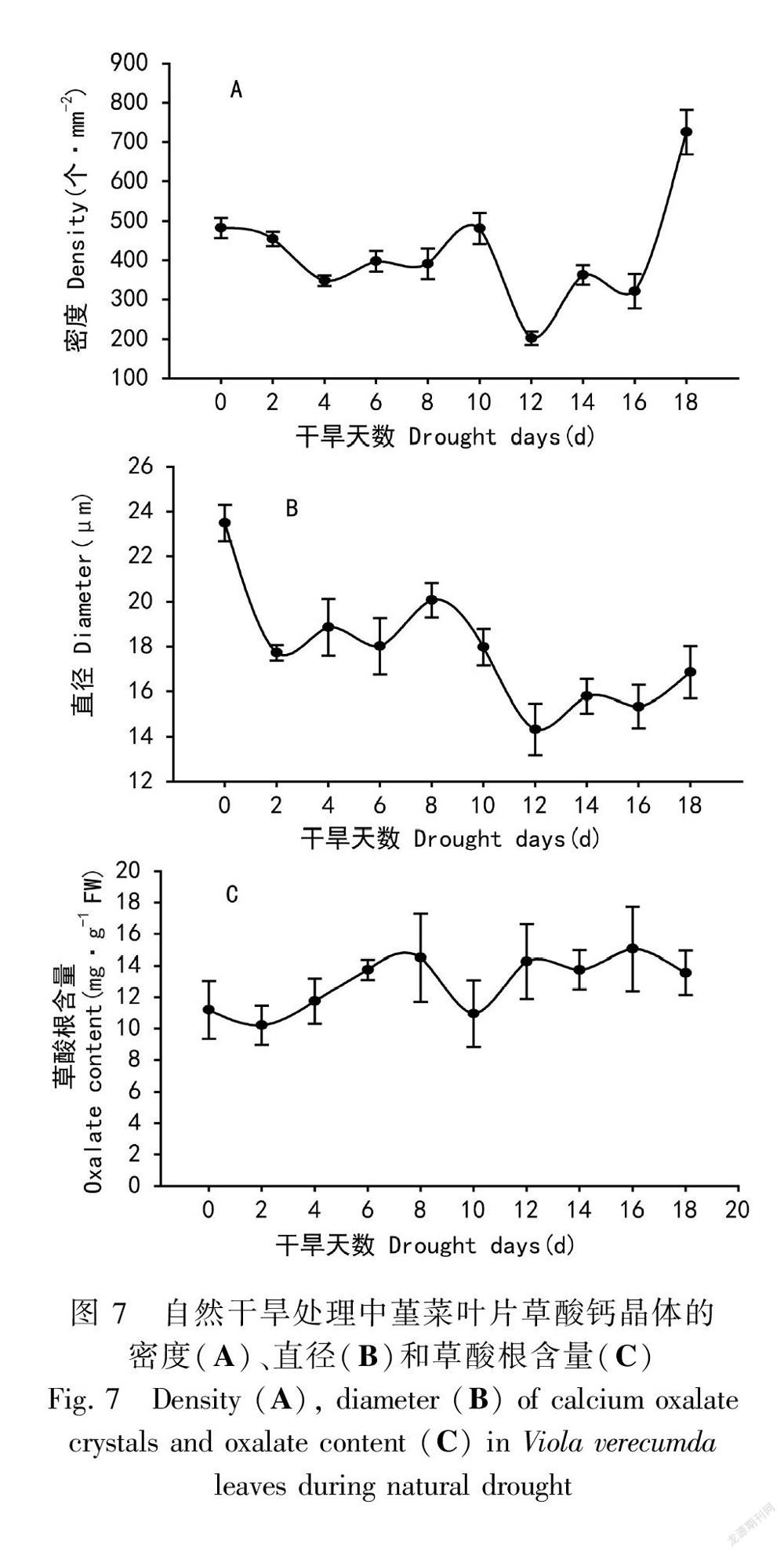
在干旱过程中,草酸钙晶体的密度和直径会发生明显变化。晶体密度上的变化主要体现在第12天时大量减少(图6:G,图7:A)和第18天时大量增加(图6:J,图7:A)。在第0天至第4 天,草酸钙晶体的密度显著减少;在第4天至第10 天,晶体的密度缓慢增加;第12天发生剧减至最低;第12天至第18 天,草酸钙晶体密度又呈现缓慢升高的趋势,并在第18 天达到最大值725 个· mm-2(图6,图7:A)。在第0 天,草酸钙晶体直径最大,为23.5 μm;干旱第2天,直径迅速下降为17.72 μm;在第12 天时,直径最低为14.33 μm,之后升高到16 μm左右(图6,图7:B)。由图7:C可知,整个干旱过程中,堇菜叶片中的总草酸根(包括可溶性草酸和草酸钙晶体中的草酸根)未见显著变化。
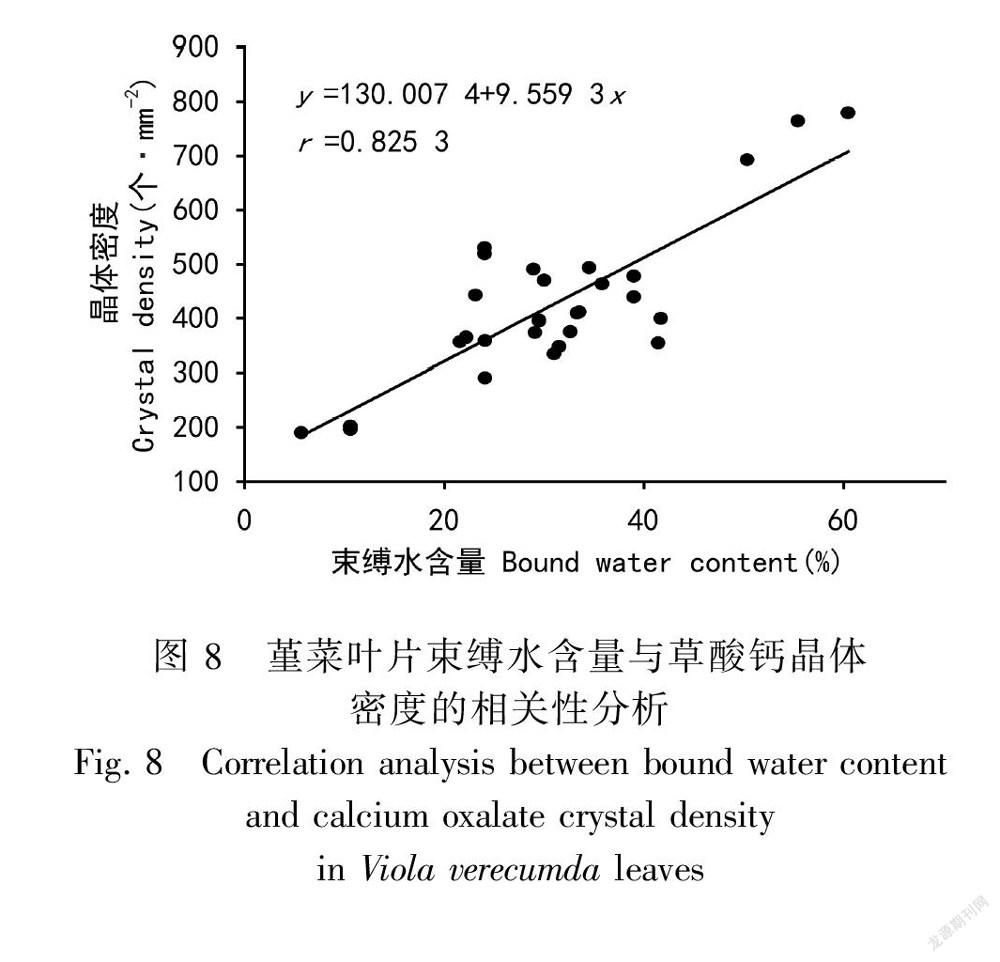
2.6 自然干旱过程中堇菜叶片草酸钙晶体密度与叶片束缚水相关性分析
为了分析束缚水和草酸钙晶体密度之间的关系,对二者进行相关性分析,分析结果表明叶片束缚水含量与草酸钙晶体的密度具有极显著的正相关关系(r= 0.825 3,P<0.000 1)。
3 討论
耐旱植物有许多的生理机制参与调节植物的耐旱特征,如渗透调节、清除活性氧对植物的伤害、增加蛋白质的稳定性等(Fang & Xiong, 2015; 邵艳军和山仑, 2006)。此外,植物叶片水分的保持也是植物耐旱的重要方式之一(Fang & Xiong, 2015),植物可通过诱导叶片气孔关闭,减少水分蒸腾从而维持叶片水分(郭自春等, 2015; 张利刚等, 2012)。另外,瓦西里也娃和董任瑞(1956)发现植物叶片内的自由水和束缚水的比值也是评价植物抗旱的一个重要指标。为此,本研究通过研究堇菜在干旱过程中叶片水分调节动态过程、气孔导度和草酸钙晶体的响应特征,揭示喀斯特适生植物的耐旱机制。
喀斯特生境的最显著特征之一是土壤含水量低,堇菜能在该生境中长期生存和繁育,这表明该植物具有一定的耐旱能力。植物叶片含水量表示叶片的水分状况,叶片相对含水量和水分饱和亏缺分别是反映植物保水能力和缺水状况的重要指标(Fang & Xiong, 2015; 吴丽丽等, 2018)。与耐旱性弱的物种相比,耐旱性强的物种能够维持较高的含水量和相对含水量(刘贤德等, 2004; 彭小玲, 2011) ,另外,彭小玲(2011)发现岩壁上生长的三种苦苣苔科植物也具有类似的保水抗旱机制。在当前研究中,堇菜叶片水分含量、相对水含量和水分饱和亏缺在干旱过程中,基本保持不变,表明了植物在干旱过程中,叶片具有较强的保水能力,从而避免干旱胁迫。
叶片束缚水和自由水的比值是耐旱性的重要指标之一(瓦西里也娃和董任瑞, 1956; 陈莉等, 2008)。在两种硬粒小麦品种Capeiti8(耐旱性)和Creso(干旱敏感)的研究中发现,耐旱性较强的Capeiti8比Creso富集了更多的束缚水(Rascio et al., 1992)。这表明叶片束缚水的富集有助于增强植物的耐旱能力。本研究中,堇菜干旱8~12 d,叶片束缚水转变为自由水,弥补干旱前期自由水的亏缺,干旱12~18 d,自由水转变成束缚水,有效防止水分流失,增强植物的耐旱性能。
草酸钙晶体定位于植物叶片细胞的液泡中,它对液泡内的水分具有较大影响。有研究发现在荒漠植物沙冬青(Ammopiptanthus mongolicus)、合头草(Sympegma regelii)和梭梭柴(Haloxylon ammodendron)的叶片含晶细胞内具有大量的草酸钙晶体,这些含晶细胞具有较高的渗透势和很强的贮水能力,因此认为含晶细胞是荒漠植物的抗旱结构特征之一(刘家琼, 1982; 朱广龙和魏学智, 2016)。本研究中,干旱0~10 d,叶片细胞内草酸钙晶体的密度变化不明显,晶体的直径降低,表明草酸钙晶体可能发生少量降解。当干旱至第12天时,草酸钙晶体直径和密度降为最低,此时叶片内束缚水含量也急剧下降,表明草酸钙晶体的密度和直径与叶片束缚水含量存在一定关系,叶水势出现显著下降,可能是草酸钙晶体分解为一些离子,如草酸根和钙离子,造成细胞渗透势的降低,导致水势降低。另外,Tooulakou(2016)研究发现草酸钙晶体降解形成二氧化碳和水。本研究通过测定草酸根含量表明干旱过程中草酸根含量未发生显著变化,叶片中大量草酸钙晶体降解,使得晶体内的束缚水被释放,增加了自由水的含量。
气孔在调节叶片水分平衡中具有重要作用,本研究中,干旱胁迫前期(0~12 d),叶片气孔导度和蒸腾速率存在波动变化,表明该阶段气孔不是限制水分丢失的主要因子。相反,干旱后期(12~18 d),气孔导度和蒸腾速率急剧下降,防止水分流失,同时晶体的密度和直径增加,表明该阶段叶片细胞内的自由水可能与草酸钙晶体形成束缚水。为更进一步分析干旱过程中束缚水和自由水的相互关系,对束缚水和草酸钙晶体密度进行相关性分析,表明草酸钙晶体的密度和束缚水之间存在显著正相关性(r = 0.825 3,P<0.000 1)。
综上所述,在干旱胁迫前期,堇菜主要通过细胞内草酸钙晶体的降解维持细胞内水分平衡;而在干旱后期,叶片通过关闭气孔,将部分自由水转变为束缚水,为下一阶段的干旱胁迫维持水分平衡。因此,堇菜植物在耐旱过程中可能协调草酸钙晶体和气孔的生理行为忍耐干旱胁迫。
参考文献:
CHOI YE, HARADA E, WADA M, et al., 2001. Detoxification of cadmium in tobacco plants:Formation and active excretion of crystals containing cadmium and calcium through trichomes [J]. Planta, 213(1):45-50.
CHEN L, YUAN HJ, WEI SZ, 2008. Analysis of the bound water / free water and transpiration intensity of Sedum plants [J]. Agric Technol, 28(6):80-81. [陈莉, 原红娟, 尉淑珍, 2008. 景天植物的束缚水/自由水与蒸腾强度的分析 [J]. 农业与技术, 28(6):80-81.]
FANG Y, XIONG L, 2015. General mechanisms of drought response and their application in drought resistance improvement in plants [J]. Cell Mol Life Sci, 72(4):673-689.
FRANCESCHI VR, NAKATA PA, 2005. Calcium oxalate in plants:Formation and function [J]. Ann Rev Plant Biol, 56(1):41-71.
GUO ZC, ZENG FJ, LIU B, et al., 2015. Photosynthetic cha-racteristics of Alhagi sparsifolia and Tamarix ramosissima and the relevant environment factors in different periods [J]. Acta Bot Boreal-Occident Sin, 35(1):189-198. [郭自春, 曾凡江, 刘波, 等, 2015. 疏叶骆驼刺和多枝柽柳不同时期光合特性日变化及其与环境因子的关系 [J]. 西北植物学报, 35(1):189-198.]
LAWLOR DW, 2013. Genetic engineering to improve plant performance under drought:Physiological evaluation of achievements, limitations, and possibilities [J]. J Exp Bot, 64(1):83-108.
LUO LJ, 2010. Breeding for water-saving and drought-resistance rice (WDR) in China [J]. J Exp Bot, 61(13):3509-3517.
LI YL, 2008. Medicinal plant resources of viola and its utiliazation in Shandong Province [J]. N Hortic, 32(1):237-238. [李彦连, 2008. 山东省堇菜属药用植物资源及利用 [J]. 北方园艺, 32(1):237-238.]
LI Y, JIANG TM, WANG J, 2009. Seasonal drought characte-ristic of guizhou karst mountainous area and its counterneasures:Taking Tongzi County as the example [J]. Guizhou Agric Sci, 37(5):43-46. [李渝, 蒋太明, 王静, 2009. 贵州喀斯特山区季节性干旱特征及对策:以桐梓县为例 [J]. 贵州农业科学, 37(5):43-46.]
LIU JQ, 1982. The xeromorphic structure of different typical plants in deserts of China [J]. Chin J Plant Ecol, 6(4):314-319. [劉家琼, 1982. 我国荒漠不同生态类型植物的旱生结构 [J]. 植物生态学报, 6(4):314-319.]
LIU XD, LI XH, LI WH, et al., 2004. Analysis on difference for drought responses of maize inbred lines at seedling stage [J]. J Maize Sci, 12(3):63-65. [刘贤德, 李晓辉, 李文华, 等, 2004. 玉米自交系苗期耐旱性差异分析 [J]. 玉米科学, 12(3):63-65.]
MACNISH AJ, IRVING DE, JOYCE DC, et al., 2003. Identification of intracellular calcium oxalate crystals in Chamelaucium uncinatum (Myrtaceae) [J]. Aust J Bot, 51(5):565-572.
PENG XL, 2011. The droughe tolerance of three gesneriaceae species in danxia landscape [D]. Xiamen:Xiamen Univer-sity:1-53. [彭小玲, 2011. 丹霞地貌3种苦苣苔科植物的耐旱性研究 [D]. 厦门:厦门大学:1-53.]
RASCIO A, PLATANI C, FONZO N, et al., 1992. Bound water in durum wheat under drought stress [J]. Plant Physiol, 98(3):908-912.
SHAO YJ, SHAN L, 2006. Advances in the research on the mechanism of plants [J]. Chin J Eco-Agric, 14(4):16-20. [邵艳军, 山仑, 2006. 植物耐旱机制研究进展 [J]. 中国生态农业学报, 14(4):16-20.]
TOOULAKOU G, GIANNOPOULOS A, NIKOLOPOULOS D,et al., 2016. Alarm photosynthesis:Calcium oxalate crystals as an internal CO2 source in plants [J]. Plant Physiol, 171(4):2577-2585.
TURNER NC, WRIGHT GC, SIDDIQUE KHM, 2001. Adaptation of grain legumes (pulses) to water-limited environments [J]. Advan Agron, 71(1):193-231.
VASILYEVA HT, DONG RR, 1956. The relationship between the ratio of free water and bound water in plant leaves and drought resistance of plants [J]. Plant Physiol J, 2(3):50-53. [瓦西里也娃, 董任瑞, 1956. 论植物叶片中自由水和束缚水的比例与植物抗旱性的关系 [J]. 植物生理学报, 2(3):50-53.]
WARD D, SPIEGEL M, SALTZ D, 1997. Gazelle herbivory and interpopulation differences in calcium oxalate content of leaves of a desert lily [J]. J Chem Ecol, 23(2):333-346.
WANG DL, ZHU SQ, HUANG BL, 2005. Preliminary study on types and quantitative assessment of karst rocky desertification in Guizhou Province, China [J]. Acta Ecol Sin, 25(5):115-121. [王德炉, 朱守谦, 黄宝龙, 2005. 贵州喀斯特石漠化类型及程度评价 [J]. 生态学报, 25(5):115-121.]
WU LL, WANG DL, LI ZY, 2018. Physiological response of Ligustrum robustum to continuous drought and rewatering [J]. Plant Physiol J, 54(5):837-844. [吴丽丽, 王德炉, 李自玉, 2018. 粗壮女贞对持续性干旱及复水的生理响应 [J]. 植物生理学报, 54(5):837-844.]
XU GB, ZENG WZ, WAN DG, 1995. Plant resources of ciola and prospects of exploitation in Sichuan Province [J]. Chin Med Mat, 18(5):227-231. [徐国兵, 曾万章, 万德光, 1995. 四川省堇菜属药用植物资源与开发前景 [J]. 中药材, 18(5):227-231.]
YI Y, SU ZM, ZHANG XM, et al., 2018. Calcium oxalate biosynthesis in plant and its defense functions [J]. J Guizhou Norm Univ (Nat Sci Ed), 36(4):1-7. [乙引, 蘇志孟, 张习敏, 等, 2018. 草酸钙在植物中的生物合成及其防御功能 [J]. 贵州师范大学学报(自然科学版), 36(4):1-7.]
ZHANG LG, ZENG FJ, LIU B, et al., 2012. Study of the photosynthesis characteristics and physical signs of four plants at the desert-oasis ecotone [J]. Acta Pratac Sin, 21(1):103-111. [张利刚, 曾凡江, 刘波, 等, 2012. 绿洲-荒漠过渡带四种植物光合及生理特征的研究 [J]. 草业学报, 21(1):103-111.]
ZHU GL, MA Y, HAN L, et al., 2014. Current status of research on morphological structure, biological function and formation mechanism of plant crystals [J]. Acta Ecol Sin, 34(22):6429-6439. [朱广龙, 马茵, 韩蕾, 等, 2014. 植物晶体的形态结构、生物功能及形成机制研究进展 [J]. 生态学报, 34(22):6429-6439.]
ZHU GL, WEI XZ, 2016. Leaf morphological plasticity of Ziziphus jujuba var. spinosa in response to natural drought gradient ecotopes [J]. Acta Ecol Sin, 36(19):6178-6187. [朱广龙, 魏学智, 2016. 酸枣叶片结构可塑性对自然梯度干旱生境的适应特征 [J]. 生态学报, 36(19):6178-6187.]
