亚金跳小蜂在不同生育期扶桑绵粉蚧寄主上的生物学特性(英文)
2019-09-10吕荣华付岗覃武
吕荣华 付岗 覃武

Abstract:【Objective】The present research was conducted to understand the influence of Aenasius arizonensis on Phenacoccus solenopsis at different development stages to provide reference for rearing and potential augmentation in biological control of P. solenopsis. 【Method】In the host choice based experiment and non-choice based experiment, 15 mealybugs of 4 different instar nymphs and 30 3rd instar larvae and adults of the mealybugs were used as test insects. A pair of A. arizonensis were placed in each cage and survival situation of host and parasitoids were recorded daily. 【Result】The results depicted that the highest host stage specific parasitism and mummy formation were recorded for 3rd instar nymph mealybugs under choice and non-choice based experiments, whereas the lowest parasitism was recorded on 2nd instar stage. Longevity of the female observed during the choice and non-choice based experiments were not significantly different from each other(P>0.05). Female A. arizonensis lived for 12.69±0.19 and 13.24±0.40 d when 2nd instar nymphs of the host provided under choice and non-choice based experiments. Similarly, female A. arizonensis lived for 15.51±0.28 and 15.12±0.14 d on 3rd instar nymphs and 15.88±0.25 and 15.75±0.24 d on adult host stage, respectively. The total development period of the parasitoid was greatly varied on different life-stage hosts. The total development period of A. ari-zonensis from egg insertion to adult emergence was the longest when 3rd instar nymphs of P. solenopsis were offered for patriotization, while the shortest time was recorded for 2nd instar nymph mealybugs under choice and non-choice based experiments. 【Conclusion】It is concluded that the maximum adult emergence of A. arizonensis was recorded on 3rd instar host stage and minimum on 2nd instar host stage.
Key words: Aenasius arizonensis; Phenacoccus solenopsis; development stage; host; biological characters
CLC number: S476.3 Document code: A Article:2095-1191(2019)09-1973-08
摘要:【目的】探讨亚金跳小蜂对不同生育期扶桑绵粉蚧的影响,为今后寄生蜂的饲养和推广及扶桑绵粉蚧的生物防治提供理论参考。【方法】分别以4个龄期的15头扶桑绵粉蚧、30头3龄幼虫和成虫为试虫进行选择性和非选择性试验,每虫笼放入1对亚金跳小蜂,每天记录寄主和寄生蜂的存活情况。【结果】亚金跳小蜂的最大寄生率和扶桑绵粉蚧的最大僵虫量均在扶桑绵粉蚧3龄若虫期出现,而最低值均在其2龄若虫期出现。在选择和非选择试验中,亚金跳小蜂雌虫的寿命差异不显著(P>0.05)。在以扶桑绵粉蚧2龄若虫为试验对象的选择性和非选择性试验中,雌性亚金跳小蜂的生命周期分别为12.69±0.19和13.24±0.40 d。在以扶桑绵粉蚧3龄若虫为试验对象的选择性和非选择性试验中,雌性亚金跳小蜂的生命周期分别为15.51±0.28和15.12±0.14 d。寄生蜂的发育周期因寄主龄期而有明显差异,在扶桑绵粉蚧3龄若虫期寄生的亚金跳小蜂生命周期最长,而在扶桑绵粉蚧3龄若虫期寄生的亚金跳小蜂生命周期最短。【结论】作为寄生蜂的亚金跳小蜂在寄主扶桑绵粉蚧3龄若虫期的成虫量最大,在2龄若虫期的成蟲量最小。
關键词: 亚金跳小蜂;扶桑绵粉蚧;生育期;寄主;生物学特性
0 Introduction
【Research significance】Aensius arizonensis is a solitary parasitoid of the invasive mealybug Phenacoccus solenopsis(Fallahzadeh et al.,2014). A. arizonensis has a wide range of host plants and most of them belong to Malvacae plant family. Mealybugs have piercing-sucking mouth parts,by which they suck cell sap and may also inject a toxin in plant ti-ssues that may cause yellowing of leaves,plant stun-ting and wilting. Several investigations had shown that this parasitoid has a high parasitism rate on P. solenopsis. In 2004,P. solenopsis was found on co-tton in Gujrat, India for the first time. P. solenopsis caused severe damage in cotton and other crops in 2005. It was found that time in Vehari district of Punjab and Sanghar district of Sindh Province. Nowa-days it has been found throughout the country infes-ting many fruits,vegetables,and field crops(Mahmood et al.,2011). 【Research progress】The cotton mealybug,P. solenopsis was originally described by Tinsley in 1898. Wang et al.(2010) reviewed its distribution and suggested that it was native to the Uni-ted States. The first record was made outside continental USA from Hawaii in 1966(Kumashiro et al.,2001); Mexico in 1978 and South America from 1985 onwards(Williams and Granara de Willink,1992) and in Asia from 2000s(Hodgson et al.,2008). Various studies focused on observing the parasitization efficiency of A. arizonensis under the laboratory conditions,and natural parasitism of P. solenopsis by the parasitoid in the field in India(Kumar et al.,2009). Subsequently,A. arizonensis was also discovered in Pakistan(Ashfaq et al.,2010). Previous laboratory experiments have demonstrated that A. arizonensis could parasitize the P. solenopsis instars(Fand and Suroshe,2015). 【Research breakthrough point】A set of experiments on parasitism rate and longevity of A. arizonensis on developmental stages of mealybug P. solenopsis were conducted through choice and non-choice based method. 【Solving pro-blems】This paper have tested hypothesis that host stages affect the biology of parasitoid,A. arizonensis in choice and non-choice based method. In the present study,biology of parasitoid was greatly affected in relation to choice and non-choice host stage. The results will be helpful in future for rearing and potential augmentation in biological control programmes of mealybugs.
1 Materials and methods
1. 1 Development of culture mealybug
The study on host-stage specific biology of A. arizonensis on P. solenopsis was conducted in the laboratory of Department of Entomology,Sindh Agriculture University,Tandojam.
The culture of P. solenopsis was developed in the laboratory,for this purpose mealybug was collec-ted from the cotton growing field and other crops at Tandojam in the morning,kept in ice chest box and brought to laboratory. In the laboratory,field collec-ted mealybugs were kept in glass cages(30.48 cm×30.48 cm×30.48 cm),pumpkin unpeeled and partially chapped were left in the cages to feed the mealybug individuals,by this way a sizeable culture of mealybug was developed.
1. 2 Parasitoids
The parasitized mealybugs called mummies,were collected from field of cotton,tomato,and brinjal,these mummies were kept in Petri dishes(2.54 cm×15.24 cm) up to the emergence,after adult emergence the parasitoids were collected in glass vials separately. After identifying them as male and female by morphology(Hayat,2009), a pair of them was shifted in one glass jar for mating purpose then released to host cage. The solution of honey and water was given to the parasitoids as food through soaked cotton swab.
1. 3 Host stage preference choice based experiment
Nymphal instars i.e.,2nd,3rd and adult females of mealybugs were kept in the same cages with piece of pumpkin as food,each cage was contained 15 mealybugs of each stage and then a pair of A. ari-zonensis was released in each cage for the parasiti-zation of mealybugs. After 24 h, the parasitoids were removed from the cages and released to other cages containing the same number and same host stages,in this way four batches of mealybugs were exposed to the parasitoids to obtain the results of host-stage preference of A. arizonensis. Mealybugs were observed every day and finally the mummies of mealybugs were collected from cages and kept in Petri dishes for emergence.
1. 4 Host stage preference non-choice based expe-riment
The 2nd and 3rd instar nymphs and adult female mealybugs were kept in separate cages respectively with a piece of pumpkin as food. Each cage was containing 30 individuals of only a single host stage,then a pair of A. arizonensis were released in each cage for the parasitization of mealybugs,after 24 h,the parasitoids were removed from the cages and released to other cages containing the same number and same host stages,in this way four batches of mealybugs were exposed to the parasitoids,observation was taken on parasitism/parasitism rate and host stage preference. Daily examination of the cages was conducted for the data collection,finally the data was statistically analyzed.
2 Results and analysis
2. 1 Oviposition rate and longevity of A. arizonensis
The oviposition rate of A. arizonensis was highly influenced by host’s developmental stages,under choice based experiment,the maximum mean parasi-tism rate(103.33 mummies) was recorded for 3rd instar host stage followed by adult host stage(37.66 mummies),whereas the minimum parasitism(11.66 mummies) was noticed when the 2nd instar mealybugs were offered as host for parsitization. The longevity of female parasitoid was recorded the maximum(24.00 d) on the 3rd host stage,while the minimum(16.00 d) reported on adult host stage,followed by 2nd instar nymph mealybugs(18.00 d)(Table 1). Under non-choice based experiment,the highest parasitism rate(155.00 mummies) was recorded for the 3rd instar host stage followed by 2nd instar host stage(68.00 mummies),whereas the minimum parasitism(57.66 mummies) was noticed when adult stage mealybugs were offered as host for parsitization. The longevity of female parasitoid with an average was 21.00,23.00 and 17.00 d for the 2nd,3rd nymphs and adult stage mealybug,respectively(Table 2).
2. 2 Host stage specific parasitisation by A. arizonensis
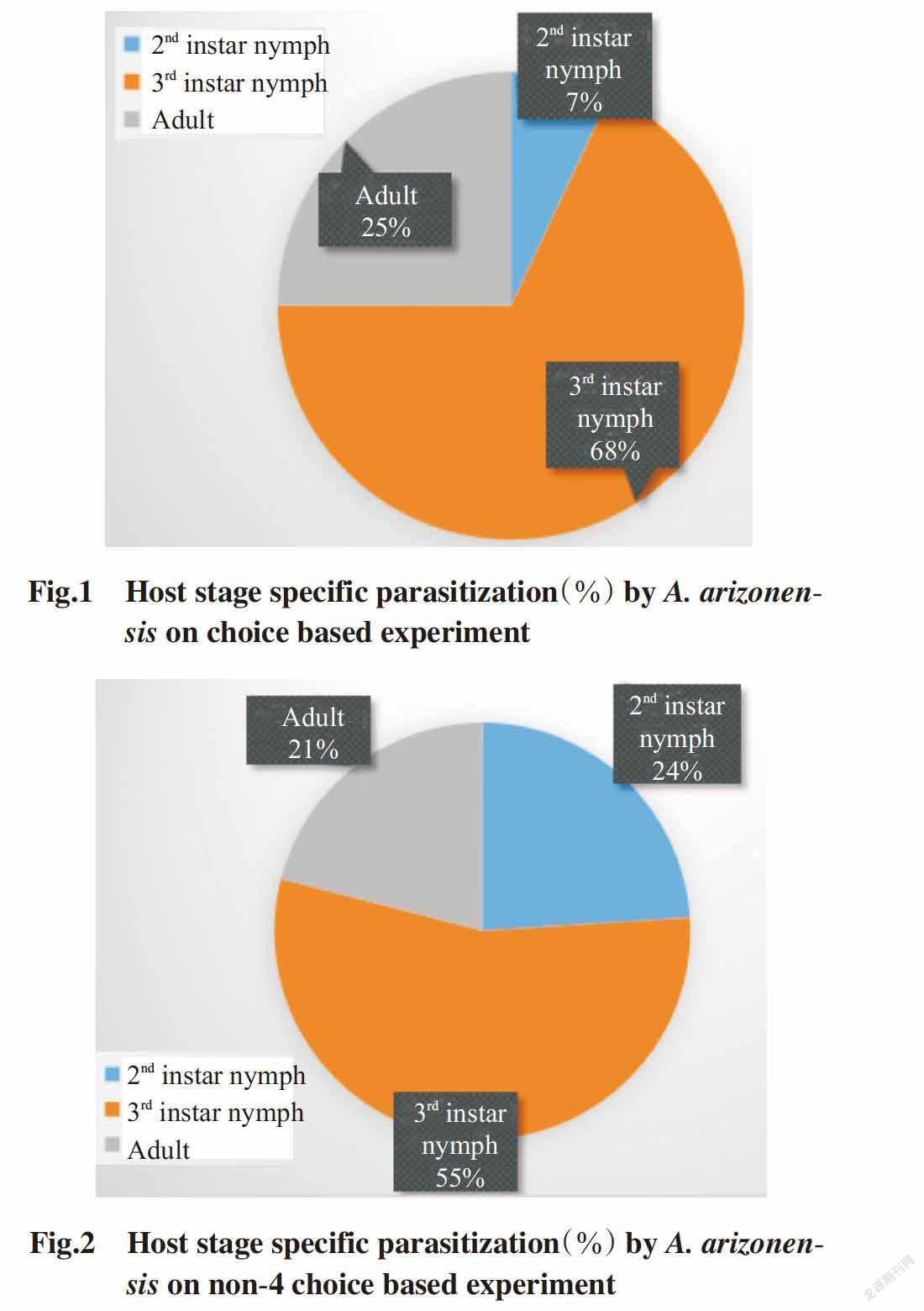
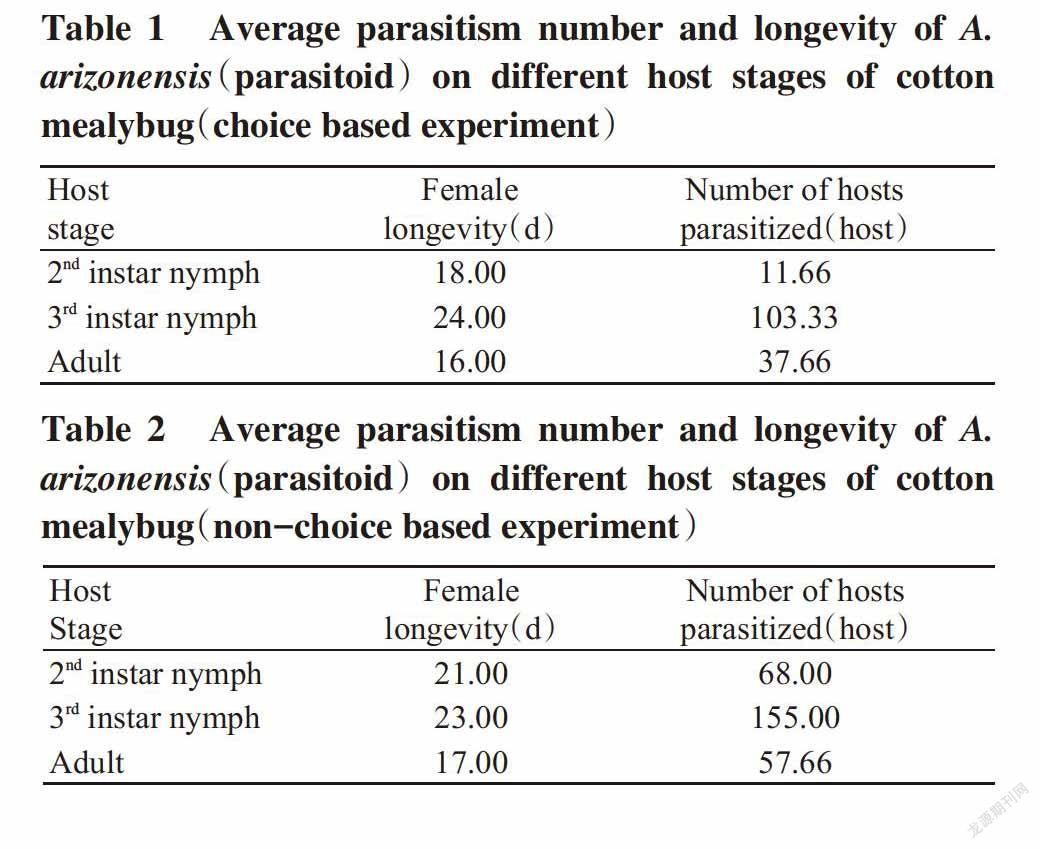
On choice based experiment,the maximum host stage specific parasitisation(68%) on P. solenopsis was observed for the 3rd instar nymph fo-llowed by 25% for adult host stage,whereas the mi-nimum host stage specific parasitisation(7%) was observed for the 2nd instar nymph. On non-choice based experiment,the maximum host stage specific parasitisation(55%) was observed for the 3rd instar nymph followed by 24% for the 2nd instar nymph stage,whereas the minimum host stage specific parasitisation(21%) was observed for adult host stage(Fig.1 and Fig.2).
2. 3 Development time from nymphs to mummy formation
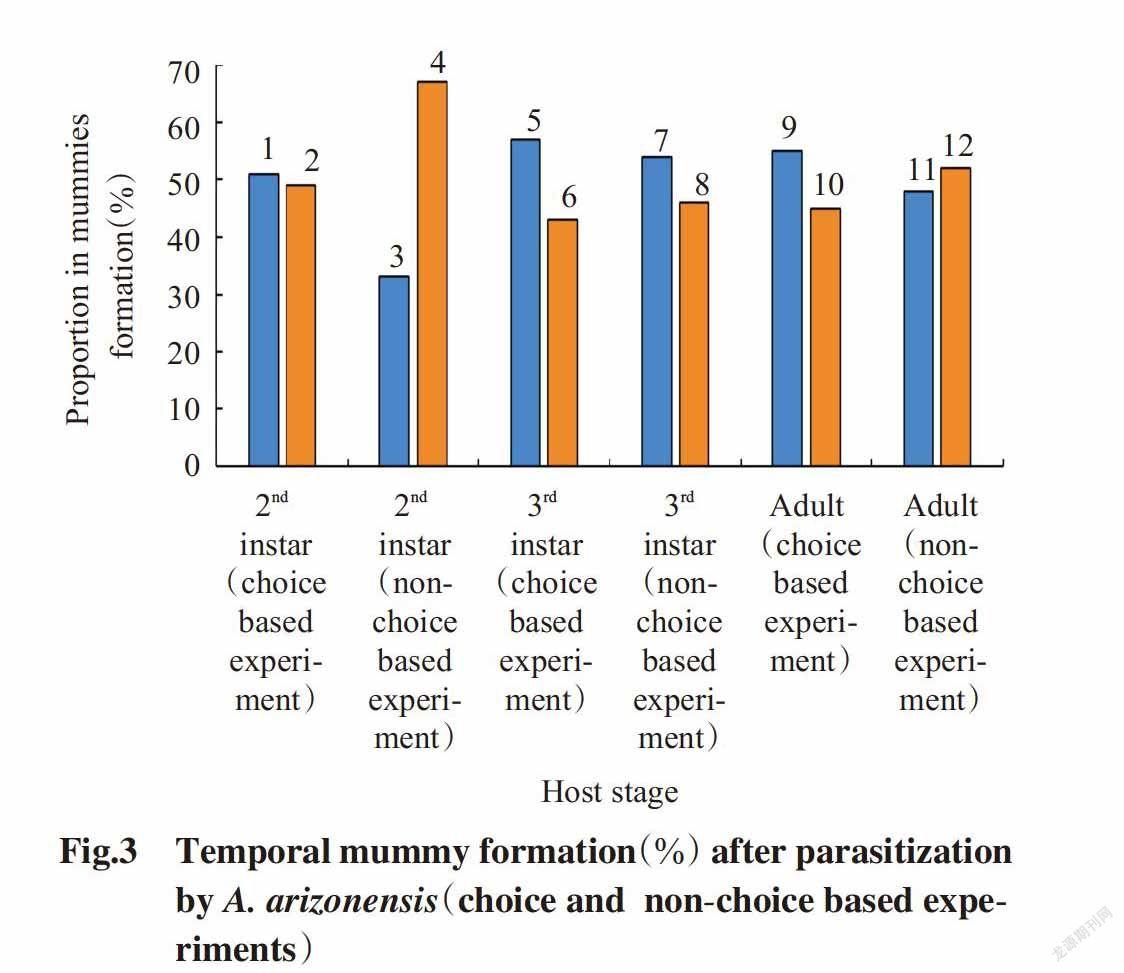
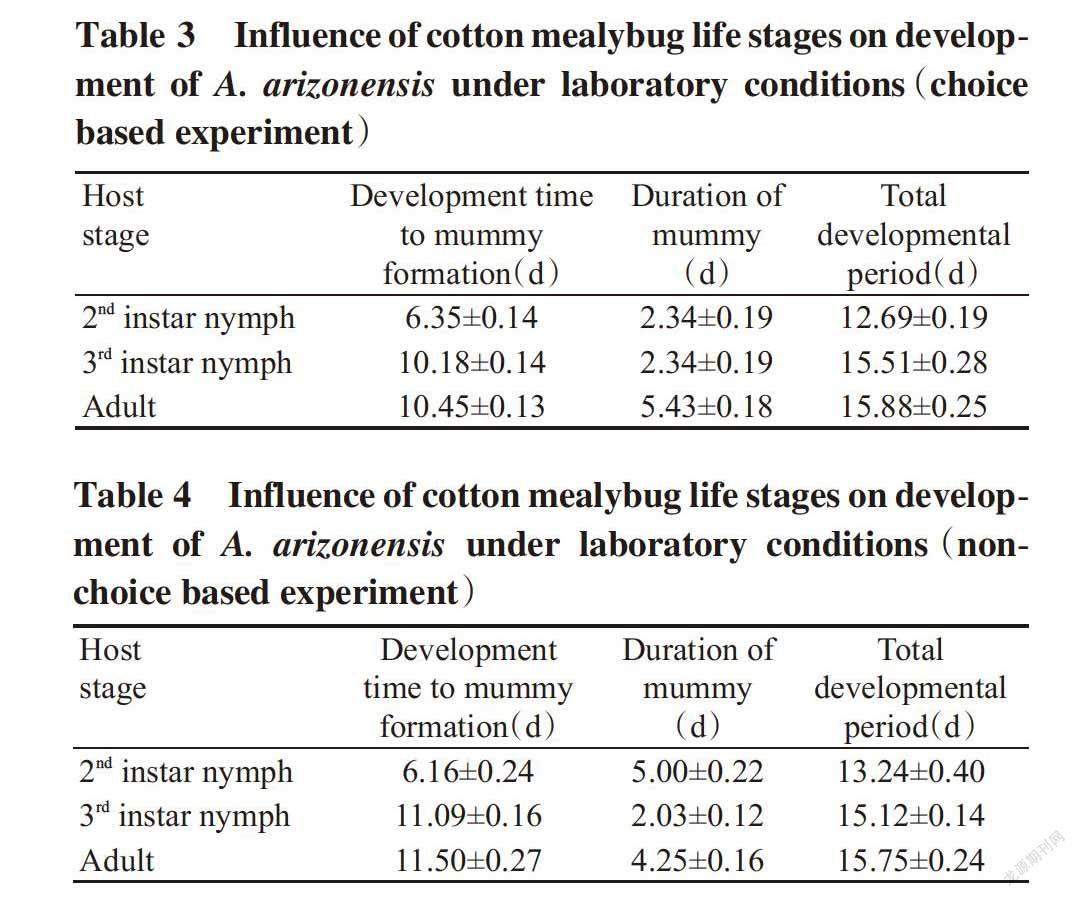
The results depicted that,under choice based experiment,the development time to mummy formation was the shortest(6.35±0.14 d) when 2nd instar nymphs of P. solenopsis was offered for parasitisation to A. arizonensis,while the longest time to mummy formation(10.45±0.13 d) was recorded for adult mealybugs,followed by 3rd instar nymphs of mealybug(10.18±0.14 d)(Table 3). The maximum(51%) mummies formed within one week as compared to mummies(49%) formed within two weeks for 2nd instar mealybug after parasitisation by A. arizonensis. In 3rd instar nymphs,the maximum mummies(57%) formed within one week as compared to mummies(43%) formed within two weeks after parasitisation by A. arizonensis. In adult mealybugs,the maximum mummies(55%) formed within one week as compared to mummies(45%) formed within two weeks after parasitisation by A. arizonensis. Whereas,under non-choice based experiment,the development time to mummy formation was the shortest(6.16±0.24 d) when 2nd instar nymphs of P. solenopsis was offered for parasitisation,while the longest time to mummy formation(11.50±0.27 d) was recorded for adult me-alybugs,followed by 3rd instar nymphs of mealybugs(11.09±0.16 d)(Table 4). The maximum mummies(67%) formed within two weeks as compared to mummies(33%) formed within one week for 2nd instar mealybug after parasitisation by A. arizonensis. In 3rd instar nymph,the maximum mummies(54%) formed within one week as compared to mummies formed within two weeks after parasitisation by A. arizonensis. In adult mealybug,the maximum mummies(52%) formed within two weeks as compared to mummies formed within one week after parasitisation by A. arizonensis(Fig.3).
2. 4 Emergence of adult A. arizonensis after formation of mealybug mummies
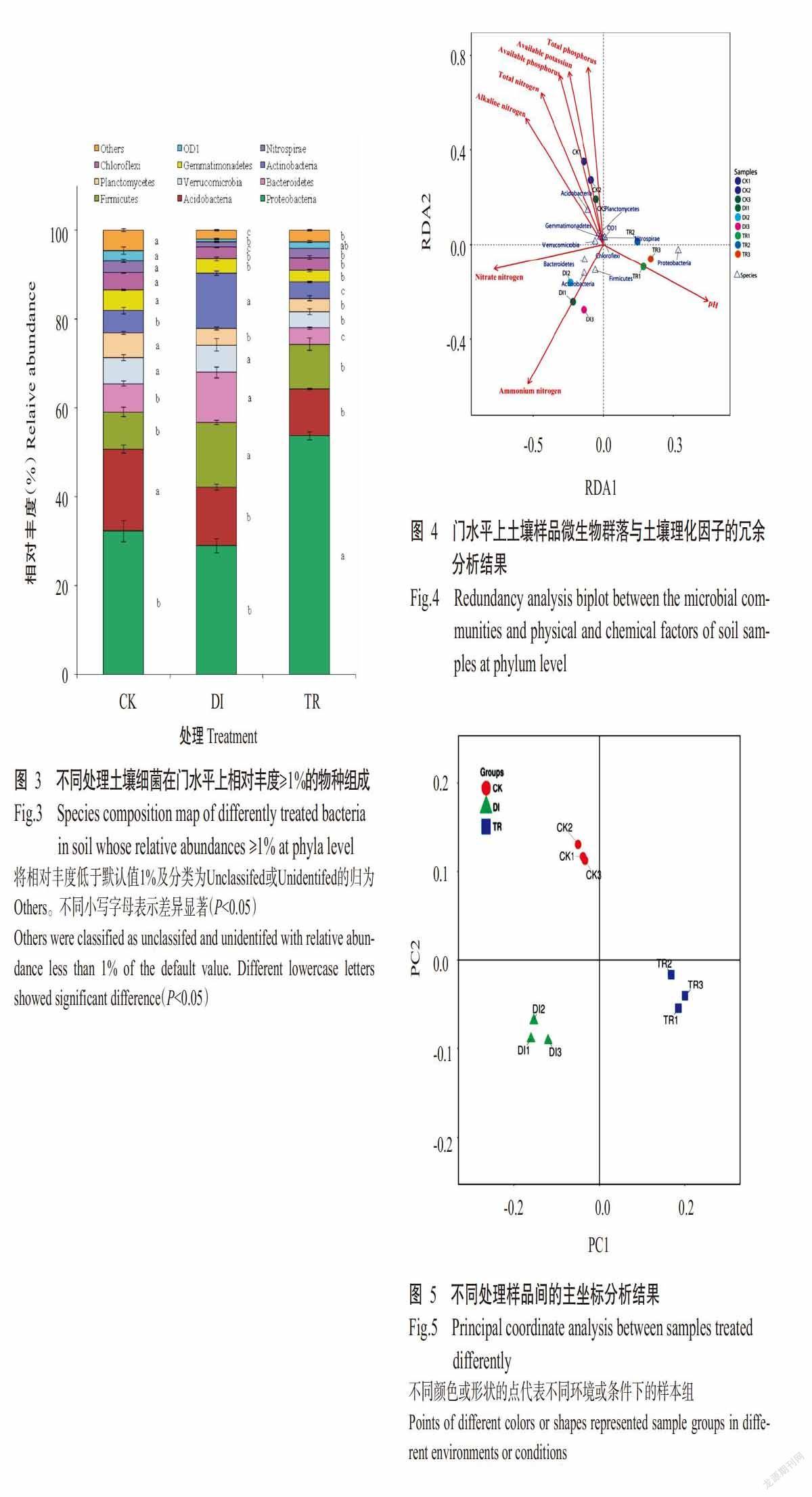
The results further showed that,the timeline to emergence of adult A. arizonensis(mummies duration) after formation of mealybug mummies was recorded the shortest(2.34±0.19 d) for both 2nd and 3rd instar nymph mealybugs and the longest mummy duration(5.43±0.18 d) was observed in adult mealybugs under choice based experiment(Table 3). In 2nd instar nymphs,the highest percentage(57%) of mu-mmies lasted up to 3 d,while minimum mummies took up to 6 d and adults of A. arizonensis emerged from those mummies. In the 3rd instar nymph and adult mealybugs,the results showed that,65% mummies lasted up to 3 d and 35% lasted up to 6 d. On the other hand,under non-choiced base experiment,the timeline to emergence of adult A. arizonensis,after formation of mealybug mummies was recorded the shortest(2.03±0.12 d) for 3rd instar nymph mealybugs,and the longest mummy duration(5.00±0.22 d) was observed in the 2nd instar nymph mealybug fo-llowed by adult mealybugs(4.25±0.16 d)(Table 4). In the 2nd instar nymphs,the highest percentage(53%) of mummies lasted up to 6 d,while minimum mummies(47%) took up to 6 d and adults of the 3rd instar nymph and adult mealybugs,the results showed that,65% mummies lasted up to 3 d and 35% lasted up to 6 d(Fig.4).
2. 5 Total development period from parasitization to adult emergence
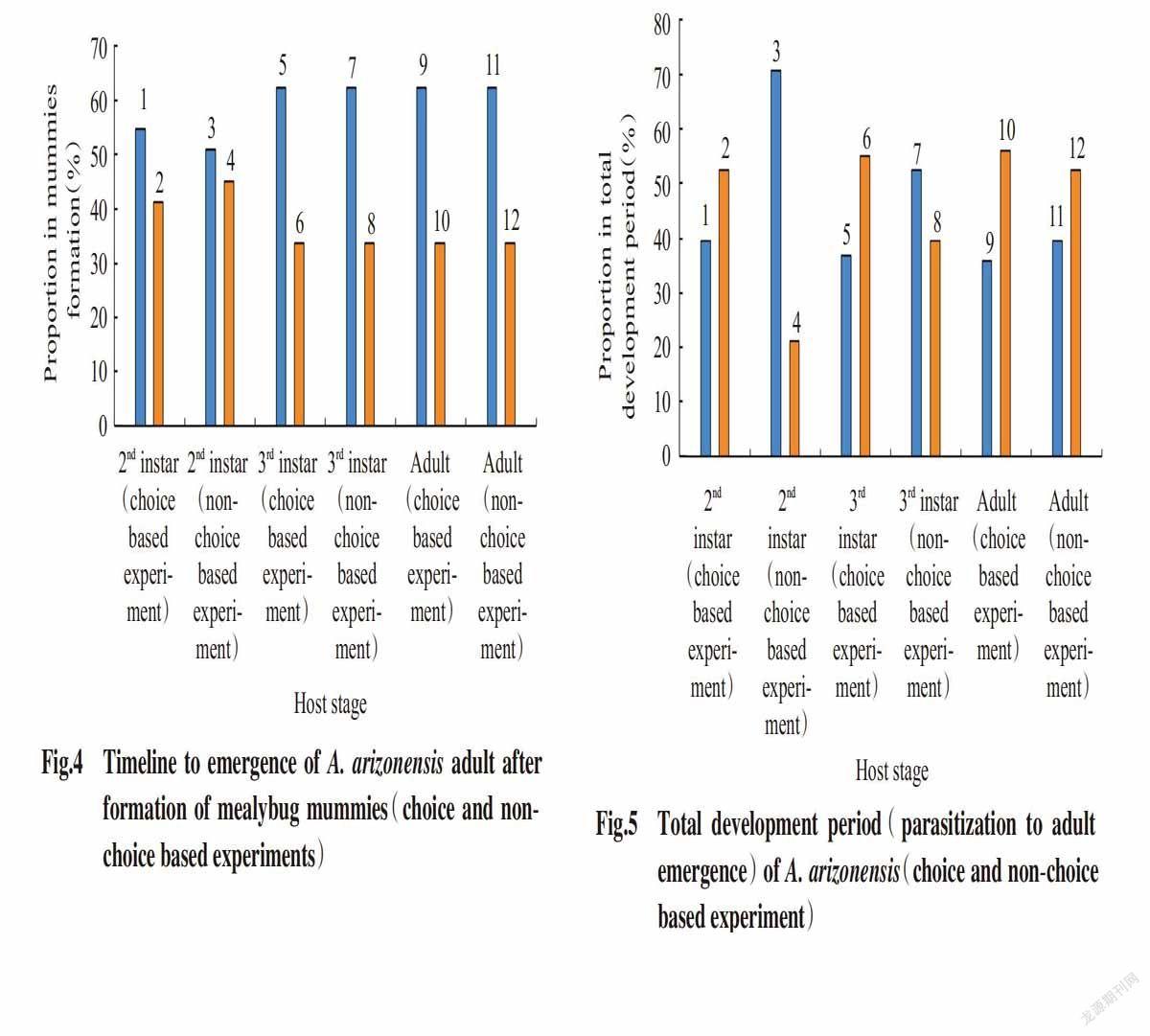
The total development period from parasitization to adult emergence of A. arizonensis was the shortest(12.69±0.19 and 13.24±0.40 d respectively) when 2nd instar nymphs of P. solenopsis was offered for parasitisation to A. arizonensis,while the longest time to total development period(15.88±0.25 and 15.75±0.24 d) was recorded at the 3rd instar nymph mealybugs,followed by the 3rd instar nymphs of mealybugs(15.51±0.28 and 15.12±0.14 d respectively),under both choice and non-choice based experiments(Table 3 and Table 4). The results revealed that,under choice based experiment,57% adults of A. arizonensis emerged within 15-17 d and 43% emerged within 12-14 d from the mummies formed in the 2nd instar nymph of mealybugs,60% adults of A. arizonensis emerged within 15-17 d and 40% emerged within 12-14 d from the mummies formed in 3rd instar nymph of mealybugs,61% adults of A. arizonensis emerged within 15-17 d and 39% emerged within 12-14 d from the mummies formed in adult mealybugs. Under non-choice based experiment,23% adults of A. arizonensis emerged within 15-17 d and 77% emerged within 12-14 d from the mummies formed in the 2nd instar nymph of mealybugs,43% adults of A. arizonensis emerged within 15-17 d and 57% emerged within 12-14 d from the mummies formed in the 3rd instar nymph of mealybugs,57% adults of A. arizonensis emerged within 15-17 d and 43% emerged within 12-14 d from the mummies formed in adult mealybugs(Fig.5).
2. 6 Adult emergence rate of A. arizonensis
The maximum adult emergence rate(96% and 93%) of A. arizonensis was recorded from mummies formed in the 3rd instar nymph mealybug,whereas the minimum(72% and 65%) was reported from mummies formed in the 2nd instar nymph mealybugs,fo-llowed by adult emergence from mummies formed in adult host stage(89% and 85%) under choice based and non-choic based experiments(Fig.6).
3 Discussion
Many pest-controlling strategies have been used in various parts of the world to check the population of mealybug under threshold levels,many chemicals were also used to control the cotton mealybug,the pesticides used against the cotton mealybug were quick,result-oriented and economic whereas on the other hand they possessed several environmental ha-zards to the natural enemies,livestock and farmer community(Noureen et al.,2016). The use of chemicals for the control of this pest was not only expensive but also disturbed the ecosystem of natural enemies. The continuous use of chemical insecticides resulted serious threats like development of resistance in insects,environmental pollution and side effects on human health.
Encrytids were the most successful natural enemies used in the biological control programs against mealybugs(Noyes and Hayat,1994). A. arizonensis(Girault) was an important koinobiont nymphal endoparasitoid of P. solenopsis reported in India(Hayat,2009),Pakistan(Mahmood,2008),China(Chen et al.,2010),Iran(Zain-ul-Abdin et al.,2012) and Australia(Khan et al.,2012). It was a key mortality factor of P. solenopsis under field conditions(Ram et al.,2009). Field parasitism by A. arizonensis to an extent of 95% has been recorded in P. solenopsis(Khuhro et al.,2012). It was reported by Mahmood(2008) for the first time in Pakistan,and later was described and named as A. bambawalei by Hayat(2009). However,Fallahzadeh et al.(2014) from Iran reported that this species was closer to A. arizonensis(Girault) and reported on A. bambawalei as a junior synonym of A. arizonensis. Host life stage was one of the key factors influencing oviposition preference,sex allocation and offspring fitness in solitary parasitoids(King,1989;Hagvar and Hofsvang,1991;Harvey et al.,1994;Kant et al.,2012). Furthermore,the progeny fitness was positively related to host resources;consequently,the parasitoid tended to allocate the offspring sexes according to host-size-dependent sex allocation principle(Charnov,1982; Godfray,1994;West and Sheldon,2002;West,2009). Parasitoids made a series of oviposition decisions while locating their hosts to achieve maximum fitness gains which included the number of eggs laid,the ratio of ferti-lized to unfertilised eggs and whether to super parasi-tize the host or not. Such decisions were relatively influenced by the quality of hosts encountered by the parasitoids(Kant et al.,2012). Earlier studies by many workers on different parasitoids have reported that host stage at the time of parasitism affected their reproductive fitness,resulting in altered deve-lopmental time(Sequeira and Mackauer,1993),body size(Lampson et al.,1996) and progeny sex ratio(Godfray,1994;Napoleon and King,1999).
The result of the present findings showed that,the parasitism rate of A. arizonensis was highly influenced by host’s developmental stages,under choice based experiment,the maximum parasitism rate was recorded for 3rd instar host stage followed by at adult host stage,whereas the minimum parasitism was noticed when 2nd instar mealybugs was offered as host for parsitization. The longevity of female parasitoid was recorded maximum(24 d) on 3rd host stage,while minimum(16 d) reported on adult host stage,followed by 2nd instar nymph mealybugs(18 d). Under non-choice based experiment,the highest parasi-tism rate was recorded for 3rd instar host stage fo-llowed by 2nd instar host stage,whereas the minimum parasitism was noticed when adult stage mealybugs were offered as host for parsitization. The longevity of female parasitoid with an average was 21,23 and 17 d for 2nd,3rd and adult stage mealybug,respectively. These results are agreement with the research ca-rried out by Solangi and Mahmood(2011) and Zain ul-Abdin et al.(2012) who discovered that in laboratory females lived 10-37 d and parasitized 32-170 mealy bugs,whereas male died within one week,their findings showed that only single parasitoid adult was emerged from each mummy. The results further depicted that,under choice based experiment,the development time to mummy formation was the shor-test(6.35±0.14 d) when 2nd instar nymphs of P. solenopsis was offered for parasitisation to A. arizonensis,while the longest time to mummy formation(10.45±0.13 d) was recorded for adult mealybugs,followed by 3rd instar nymphs of mealybug(10.18±0.14 d). The maximum mummies formed within one week as compared to that formed within two weeks for 2nd instar mealybug after parasitisation by A. arizonensis. In 3rd instar nymphs,the maximum mu-mmies formed within one week as compared to mu-mmies formed within two weeks after parasitisation by A. arizonensis. In adult mealybugs,the maximum mummies formed within one week as compared to that formed within two weeks after parasitisation by A. arizonensis under non-choice based experiment,the development time to mummy formation was shortest(6.16±0.24 d) when 2nd instar nymphs of P. solenopsis was offered for parasitisation,while the longest time to mummy formation(11.50±0.27 d) was recorded for adult mealybugs,followed by 3rd instar nymphs of mealybugs(11.09±0.16 d). The maximum mummies formed within two weeks as compared to that formed within one week for 2nd instar mealybug after parasitisation by A. arizonensis. In 3rd instar nymph,the maximum mummies formed within one week as compared to mummies formed within two weeks after parasitisation by A. arizonensis. In adult mealybug,the maximum mummies formed within two weeks as compared to that formed within one week after parasitisation by A. arizonensis. The results further showed that,the timeline to emergence of adult Aenasius arizonensis(mummies duration) after formation of mealybug mummies was recorded the shortest(2.34±0.19 d) for both 2nd and 3rd instar nymph mealybugs,and the longest mummy duration(5.43±0.18 d) was observed in adult mealybugs under choice based experiment. In 2nd instar nymphs,the highest percentage of mummies(57%) lasted up to 3 d,while minimum mummies(34%) took up to 6 d and adults of A. arizonensis emerged from those mummies. In 3rd instar nymph and adult mealybugs,the results showed that,65% mummies lasted up to 3 d and 35% lasted up to 6 d,on the other hand,under non-choice based experiment,the timeline to emergence of adult A. arizonensis after formation of mealybug mummies was recorded the shortest(2.03±0.12 d) for 3rd instar nymph mealybugs,and longest mummy duration(5.00±0.22 d) was observed in 2nd instar nymph mealybug followed by adult mealybugs(4.25±0.16 d). In 2nd instar nymphs,the highest percentage(53%) of mummies lasted up to 6 d,while minimum mummies(47%) took up to 3 d and adults of A. arizonensis emerged from those mummies. In 3rd instar nymph and adult mealybugs,the results showed that,65% mummies lasted up to 3 d and 35% lasted up to 6 d. The total development period from parasitization to adult emergence of Aenasius arizonensis was shortest the(12.69±0.19 and 13.24±0.40 d) when 2nd instar nymphs of P. solenopsis was offered for parasitisation to A. arizonensis,while longest time to total development period was recorded for 3rd instar nymph mealybugs,followed by(15.51±0.28 and 15.12±0.14 d) 3rd instar nymphs of mealybugs(15.88±0.25 and 15.75±0.24 d) under both choice and non-choice based experiments(respectively).
The results revealed that,under choice based experiment,57% adults of A. arizonensis emerged within 15-17 d and 43% emerged within 12-14 d from the mummies formed in 2nd instar nymph of mealybugs,60% adults of A. arizonensis emerged within 15-17 d and 40% emerged within 12-14 d from the mummies formed in 3rd instar nymph of mealybugs,61% adults of A. arizonensis emerged within 15-17 d and 39% emerged within 12-14 d from the mummies formed in adult mealybugs. Under non-choice base experiment,23% adults of A. ari-zonensis emerged within 15-17 d and 77% emerged within 12-14 d from the mummies formed in 2nd instar nymph of mealybugs,43% adults of A. arizonensis emerged within 15-17 d and 57% emerged within 12-14 days from the mummies formed in 3rd instar nymph of mealybugs,57% adults of A. arizonensis emerged within 15-17 d and 43% emerged within 12-14 d from the mummies formed in adult mealybugs,The results of the present study are in conformity with those conducted by Zain-ul-Abidin et al.(2012) and Sangle et al.(2013) with slight deviations due to changes in environmental conditions. These results are also in accordance with the findings of Cross and Moore(1992) and Bokonon-Ghanta et al.(1995) who discovered that mostly encyrtids prefer to parasitize late(3rd) instars and adult mealybugs.
4 Conclusion
It is concluded from the present findings that,the host stage is climacteric factor affecting the parasitization development,female longevity and adult emergence of A. arizonensis. As the results showed that,the maximum parasitization was recorded on 3rd instar mealybug,the female A. arizonensis lived longer on 3rd instar host stage,compared to 2nd instar nymphs and adult mealybugs under choice and non-choice base experiment. Adult emergence of A. arizonensis was higher on 3rd instar nymph hosts compared to 2nd instar nymphs and adult mealybugs under both choice and non-choice based experiments.
References:
Ashfaq M,Shah G S,Noor A R, Ansari S P,Mansoor S. 2010. Report of a parasitic wasp(Hymenoptera:Encyrtidae) parasitizing cotton mealybug(Hemiptera:Pseudococcidae) in Pakistan and use of PCR for estimating parasitism levels[J]. Biocontrol Science and Technology,20(6): 625-630.
Bokonon-Ganta A H,Neuenschwander P,Alphen J J M,Vos M. 1995. Host stage selection by Anagyrus mangicola (Hymenoptera: Encyrtidae),a parasitoid of the mango mealybug,Rastrococcus invadens(Homoptera: Pseudococcidae)[J]. Biological Control, 5(4): 479-486.
Charnov E L. 1982. The theory of sex allocation[M]. Princeton: Princeton University Press.
Chen H,Cao R,Xu Z. 2010. First record of Aenasius bambawalei Hayat(Hymenoptera: Encyrtidae),a parasitoid of the mealybug,Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae) from China[J]. Journal of Environmental Entomology,32(2): 280-282.
Cross A E,Moore D. 1992. Developmental studies on Anagyrus mangicola(Hymenoptera: Encyrtidae),a parasitoid of the mealybug,Rastrococcus invadens(Homoptera: Pseudococcidae)[J]. Bulletin of Entomological Research,82(3): 307-312.
Fallahzadeh M,Japoshvili G,Abdimaleki R,Saghaei N. 2014. New records of Tetracneminae(Hymenoptera,Chalcidoidea,Encyrtidae) from Iran[J]. Turkish Journal of Zoology,38(4): 515-518.
Fand B B,Suroshe S S. 2015. The invasive mealybug Phenacoccus solenopsis Tinsley,a threat to tropical and subtropical agricultural and horticultural production systems a review[J]. Journal of Crop Protection,69: 34-43.
Godfray H C J. 1994. Parasitoids: Behavioral and evolutiona-ry ecology[M]. Princeton: Princeton University Press.
Hagvar E B,Hofsvang T. 1991. Aphid parasitoids(Hymenoptera:Aphidiidae): Biology,host selection,and use in bio-logical control[J]. Biocontrol News Information,12:3-41.
Harvey J A, Harvey I F, Thompson D J. 1994. Flexible larval growth allows use of a range of host sizes by a parasitoid wasp[J]. Ecology,75(6): 1420-1428.
Hayat M. 2009. Description of a new species of Aenasius Walker(Hymenoptera:Encyrtidae),parasitoid of the mealybug,Phenacoccus solenopsis(Homoptera:Pseudococcidae) in India[J]. Biosystematica,33(3): 21-26.
Hodgson C,Abbas G,Arif M J,Saeed S,Karar H. 2008. Phenacoccus soleapsis Tinsley(Sternorrhyncha:Caccoidae: Pseudococcidae),an invasive mealybug damaging cotton in Pakistan and India with a discussion on seasonal morphological variation[J]. Zootaxa,1913: 1-35.
Kant R,Minor M A,Trewick S A. 2012. Fitness gain in a koinobiont parasitoid Diaeretiella rapae(Hymenoptera: Braconidae) by parasitising hosts of different ages[J]. Journal of Asia-Pacific Entomology,15(1):83-87.
Khan M,Quade A,Byers K,Hall Z,Grams R. 2012. Record of Aenasius bambawalei Hayat,a parasitoid of solenopsis mealybug,in Australia[C]//Proceedings of 16th Australian Cotton Conference. Cotton Research and Development Corporation.
Khuhro S N,Kalroo A M,Mahmood R. 2012. Present status of mealy bug Phenacoccus solenopsis(Tinsley) on co-tton and other plants in Sindh(Pakistan)[C]//Book of Papers. World Cotton Research Conference-5. https://www.icac. org/meetings/wcrc/wcrc5/Proceedings.pdf.
King B H. 1989. A test of local mate competition theory with a solitary species of parasitoid wasp,Spalangia cameroni[J]. Oikos,55(1): 50-54.
Kumar R, Kranthi K R, Monga D,Jat S L. 2009. Natural parasitization of Phenacoccus solenopsis Tinsley(Hemiptera: Pseudococcidae) on cotton by Aenasius bambawalei Hayat(Hymenoptera: Encyrtidae)[J]. Journal of Biological Control,23(4): 457-460.
Kumashiro B R,Heu R A,Nishida G M,Beardlsley J W. 2001. New state records of immigrant insects in the Hawaiian Islands for the year 1999[C]//Proceedings of the Hawaiian Entomological society,35: 170-184.
Lampson L J,Morse J G,Luck. R F. 1996. Host selection,sex allocation,and host feeding by Metaphycus helvolus (Hymenoptera:Encyrtidae) on Saissetia oleae(Homoptera:Coccidae) and its effect on parasitoid size,sex,and quality[J]. Environmental Entomology,25(2): 283-294.
Mahmood R. 2008. Breakthrough in biological control of mealybug in Pakistan[J]. Biocontrol News Information,29: 38-39.
Mahmood R,Aslam M N,Solangi G S,Samad A. 2011. Historical perspective and achievements in biological mana-gement of cotton mealy bug Phenacoccus solenopsis Tinsley in Pakistan[C]//The 5th Meeting Asian Cotton Research and Development Network,1-17.
Napoleon M E,King B H. 1999. Offspring sex ratio response to host size in the parasitoid wasp Spalangia endius[J]. Behavioral Ecology and Sociobiology,46(5): 325-332.
Noureen N,HussainM,Fatima S,Ghazanfar M. 2016. Cotton mealybug management: A review[J]. Middle-East Journal of Scientific Research,24(8): 2424-2430.
Noyes J S,Haya M. 1994. Oriental mealybug parasitoids of the Anagyrini(Hymenoptera:Encyrtidae)[M]. Wallingford: CAB International.
Ram P,Saini R K,Vijaya. 2009. Preliminary studies on field parasitization and biology of solenopsis mealybug parasi-toid,Aenasius bambawalei Hayat(Encyrtidae:Hymenoptera)[J]. Journal of Cotton Research and Development,23(2): 313-315.
Sangle P M,Korat D M,Patel B H. 2013. A note on morpho-metry and post larval life stages of Aenasius bambawalei Hayat[J]. Karnataka Journal of Agricultural Sciences,26(1): 152-154.
Sequeira R,Mackaue M. 1993. The nutritional ecology of parasitoid wasp,Ephedrus californicus baker(Hymenoptera: Aphidiidae)[J]. Canadian Entomologist,125(3):423-430.
Solangi G S,Mahmood R. 2011. Biology,host specificity and population trends of Aenasius bambawalei Hayat and its role in controlling mealybug Phencoccus solenopsis Tinsley at Tandojam Sindh[C]//The 5th Meeting Asian Cotton Research and Development Network,1-7.
Tinsley J D. 1898. Notes on Coccidae,with descriptions of new species[J]. Canadian Entomologist,30(12):317-320.
Wang Y,Watson G W, Zhang R. 2010. The potential distribution of an invasive mealybug,Phenacoccus solenopsis and its threat to cotton in Asia[J]. Agricultural and Fo-rest Entomology,12(4): 403-416.
West S A. 2009. Sex allocation[M]. Woodstock: Princeton University Press.
West S A,Sheldon B C. 2002. Constraints in the evolution of sex ratio adjustment[J]. Science,295(5560): 1685-1688.
Williams D J, Granara de Willink M C. 1992. Mealybugs of central and south America[M]. London: CAB Internatio-nal.
Zain-ul-Abdin,Muhammad J A,Muhammad D G,Muhammad A,Hussain F,Abbas S K,Shaina H,Manzoor A. 2012. Biological characteristics and host stage preference of mealybug parasitoid Aenasius bambawalei(Hymenoptera:Encyrtidae)[J]. Journal of Agricultural Entomology. 11(1): 198-204.
(責任编辑 陈德元)
