长柄双花木叶性状异速生长关系随发育阶段和海拔梯度的变化
2019-09-10耿梦娅陈芳清吕坤王玉兵官守鹏刘杨赟
耿梦娅 陈芳清 吕坤 王玉兵 官守鹏 刘杨赟
摘 要:長柄双花木 (Disanthus cercidifolius var. longipes) 是一种仅分布于我国东南地区的珍稀濒危植物。为研究该物种叶性状异速生长关系和叶片资源利用策略及其随发育阶段和海拔梯度的变化规律,该文以分布于江西省不同海拔梯度的长柄双花木群落为研究对象,调查分析了群落中不同发育阶段长柄双花木植株的叶片面积、叶片体积以及叶片含水量与叶片干重之间的异速关系。结果表明:不同发育阶段植株之间叶性状异速生长关系有着显著差异。成树叶片面积的增长速度低于或等于叶片干重的增长速度,幼树、幼苗叶片面积的增长速度低于叶片干重的增长速度;成树叶片体积与叶片干重呈等速增长,幼树、幼苗叶片体积的增长速度高于叶干重的增长速度;成树叶片含水量的增长速度低于叶干重的增长速度,幼树、幼苗两性状间保持等速增长。海拔梯度对长柄双花木叶性状异速生长关系也有影响,植株叶体积和叶含水量与叶干重的异速生长指数在不同海拔间有显著性差异。在低海拔区域,叶体积与叶干重呈等速增长,叶含水量的增长速度低于叶片干重的增长速度。在高海拔区域,叶体积的生长速度低于叶干重的生长速度,叶含水量和叶片干重呈等速增长。这说明长柄双花木叶片资源投资策略随着发育阶段和海拔梯度的不同发生变化。成树主要将叶生物量投资于光捕获面积和同化结构,幼树和幼苗则主要投资于维管组织的建设。由于海拔升高会引起风力增大、光强增强和土壤理化性质改变,长柄双花木在中低海拔倾向于增大叶体积以抢占资源,在高海拔倾向于加强机械组织和维管组织的建设来抵抗外界因子干扰。
关键词: 长柄双花木, 发育阶段, 异速生长, 资源利用策略, 江西省
Changes of allometric relationships among leaf traits of Disanthus cercidifolius var. longipes in different ontogenetic stages and altitude gradients
GENG Mengya1, CHEN Fangqing1*, L Kun1, WANG Yubing1, GUAN Shoupeng1, LIU Yangyun2
( 1. Hubei International Scientific and Technological Cooperation Center of Ecological Conservation and Management in Three Gorges Area, Yichang 443002, Hubei, China; 2. Hubei Zhengjiang Environmental Science & Technology Co. Ltd., Yichang 443002, Hubei, China )
Abstract:Disanthus cercidifolius var. longipes is a rare and endangered plant distributed only in the southeast area of China. In order to research the allometric relationship of leaf trait, characteristic of leaf resource utilization, and their changes with ontogenetic development and altitude gradient, we investigated and analyzed allometric relationships among the lamina mass, lamina area, lamina volume and lamina water content of D. cercidifolius var. longipes plants at different developmental stages in D. cercidifolius var. longipes communities distributed along altitude gradient in Jiangxi Province. The results showed that there were significant different allometric relationships among plants at different deve-lopmental stages. The increasing rate of lamina area was lower than or equal with that of lamina mass in adult trees, meanwhile the growth rate of lamina area was lower than that of lamina mass in saplings and seedlings; The lamina vo-lume and lamina mass of adult trees grew at the same rate, but the lamina volume of saplings and seedlings grew faster than the lamina mass of them; The growth rate of lamina water content was slower than that of lamina mass in adult trees, meanwhile the growth rate of both characters grew at the same rate in saplings and seedlings. Altitude gradient also influenced the allometric relationship among leaf traits. The allometric relationships between the lamina volume, lamina water content and lamina mass differed significantly among altitude gradients. At the low elevation, the lamina volume and lamina mass grew at the same rate, meanwhile lamina water content grew slower than lamina mass. At the high elevation, the lamina volume grew slower than lamina mass, and the lamina water content and lamina mass grew at the same rate. These results suggested that the investment strategy of leaf resources of D. cercidifolius var. longipes changed with the change of ontogenetic development and altitude gradient. The adult trees of D. cercidifolius var. longipes invested more lamina biomass in the development of light-harvesting area and assimilation structure, meanwhile the saplings and seedlings mainly invested lamina biomass in the development of vascular tissue. As the increase of altitude would increase wind and light intensity, and change physicochemical properties, D. cercidifolius var. longipes plants tends to increase the lamina volume to capture more resources at the middle and lower altitudes, and tends to strengthen the construction of machinery and vascular tissue to resist external interference at the higher altitude.
Key words: Disanthus cercidifolius var. longipes, developmental stage, allometry, resource utilization strategy, Jiangxi Province
表型可塑性是指生物体同一基因型为适应不同环境而产生不同表现型的特性(Huey et al., 2000)。某一性状的可塑性既可以遗传,也可以单独通过自然选择进行进化(Pigliucci,2001)。异速生长指的是生物体某一特征的相对生长速率不等于第二种特征相对生长速率的现象(Bertalanfy,1999),是由遗传所决定的一种稳定特性。植物总是朝最优的异速生长曲线进化(La Barbera,1989)。植物本身的异速生长只是“外观可塑性”,不是真正意义上的表型可塑性(McConnaughay & Coleman,1999),而不同环境下,异速生长曲线的变化,才是真正的表型可塑性(陆霞梅等,2007)。植物的异速生长与生物量分配、植物对资源的利用、植物对异质环境的形态适应都有着十分密切的关系(Bertalanfy,1999)。对植株异速生长的探究由生物量的分配开始,现在已经渗透到生态系统范围内的各个层面,有些研究甚至到了基因组水平(West & Brown,2005)。叶片是植物进行光合的重要器官,其所合成的有机化合物是生长发育的物质基础。叶片同时又是植物对外界资源条件变化反应最为敏感的器官,环境条件的变化常常引起叶片生长及其形态结构的变化(何春霞等,2013;杨国一等,2018)。因此,叶片性状异速生长关系成为了植物异速生长特性研究的热点。目前,关于叶片性状在种间水平上比较常见(Wright et al., 2004;祝介东等,2011),而种内水平的研究并不多见(Milla & Reich,2007)。植物叶片形态的变化可能来自植株大小的变化,也可能是由环境或资源的变化造成的(Weiner,2004;朱恕英等,2019)。研究不同发育时期植物叶性状间的异速生长能客观反映其生长关系随植物发育的变化,而研究植物在不同环境条件下叶性状间生长关系的差异则有助于揭示植物叶片对资源条件变化是否存在可塑性变化,以及植物叶片对资源条件变化的响应(Lleonart et al., 2000)。
长柄双花木(Disanthus cercidifolius var. longipes)是金缕梅科(Hamamelidaceae)双花木属(Disanthus)的一种多年生落叶灌木。双花木属系金缕梅科孑遗的单种属,其原种仅分布于日本,长柄双花木是该属中国-日本植物区系的替代种,该属在探索长柄双花木系统发育和东亚长柄双花木区系地理方面具有重要的科学意义(傅立国,1992)。该物种分布区域狭小,仅见于赣浙湘一带部分地区,被列为国家二级保护树种(高浦新等,2013)。长柄双花木高为2~6 m,4月上旬展叶,10 月下旬落叶(魏来,2015),主要生长于海拔为600 ~ 1 100 m的常绿阔叶林和针阔叶混交林中(张嘉茗等,2013),为湿生耐阴种(廖飞勇,2010)。目前,关于长柄双花木生态学方面的研究,主要包括群落区系特征及种群遗传多样性(张嘉茗等,2013;谢国文等,2014)、种群生物学(肖宜安等,2004)等,未见长柄双花木叶性状间异速生长关系随发育过程与种群环境的不同而变化的相关研究。本研究在野外勘察基础上,选择了4个分布于江西省不同海拔的长柄双花木群落作为研究对象,调查分析了这些群落中不同发育阶段长柄双花木植株的叶片面积、叶片体积、叶片含水量与叶片干重之间的异速生长关系。旨在:(1)分析不同生长发育阶段长柄双花木叶片性状异速生长关系的差异,揭示该物种在不同发育阶段对资源利用方式的转变;(2)分析分布于不同海拔梯度长柄双花木植株叶性状间异速生长曲线的变化,探讨这种变化能否展现叶片性状的可塑性,揭示该物种对资源状况变化的响应及投资策略的变化。
1 材料与方法
1.1 研究地概况
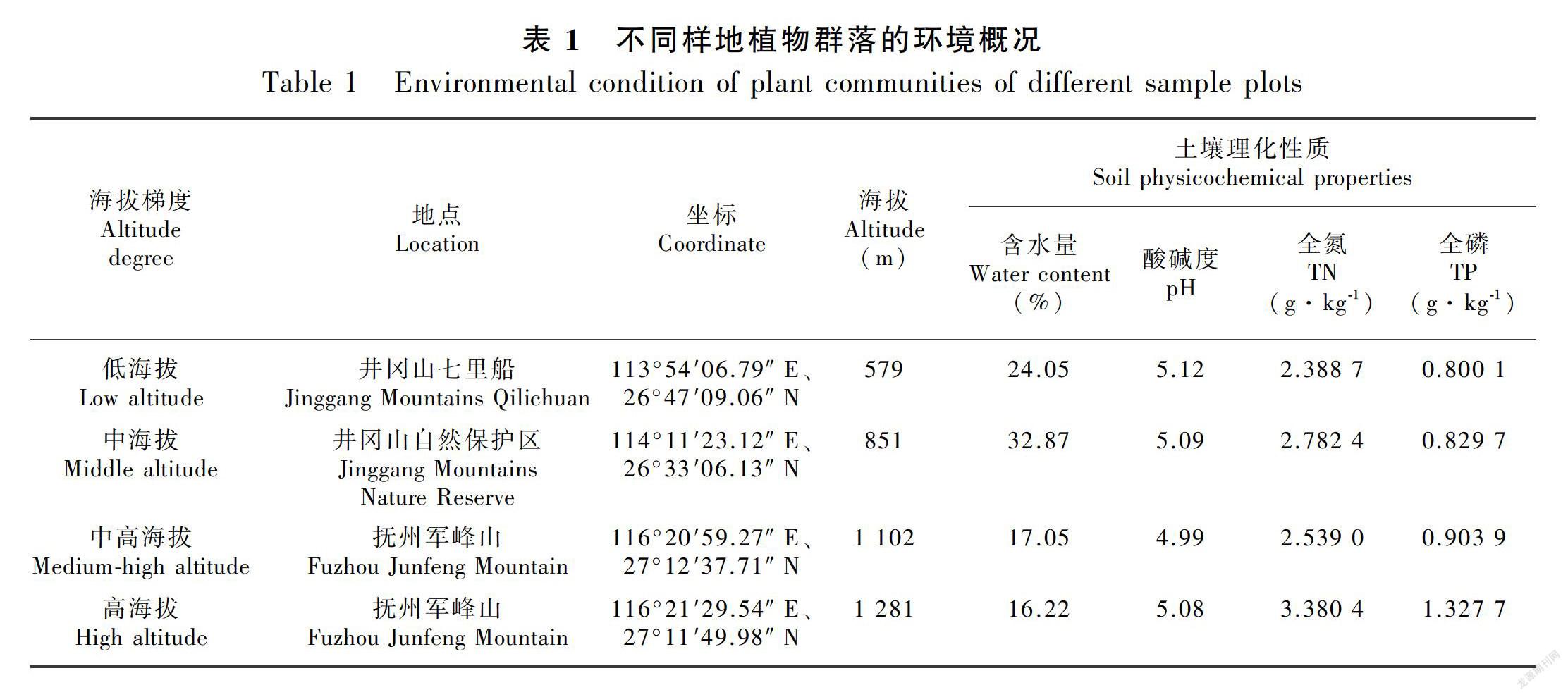
本研究选择江西省井冈山市和江西省抚州市南丰县军峰山作为研究地点。所在地区属于亚热带季风气候,年均温度为 14.2~17 ℃,最低温度为-10 ~ -6 ℃,最高温度为 38 ~ 39 ℃,年均降雨量为 1 852.6~1 875 mm。土壤为黄壤,一般为酸性(李象钦等,2014)。区域内的典型植被为亚热带常绿阔叶林(谢国文等,2016)。
1.2 样地调查
通过野外勘察,分别在井冈山和军峰山不同海拔梯度选择组成与结构较为典型的长柄双花木群落为研究对象(表1),2017年8月分别对各群落的长柄双花木种群的叶功能性状进行了野外调查。在每个长柄双花木群落中,分别选取生长在光照充沛条件下长势良好的长柄双花木成树(基径>4 cm,树高>3 m)和幼樹(树高1~2 m)各10 棵,幼苗(树高0.3~1 m)15棵(肖宜安等,2004;姚婧等,2013)。每棵树从东西南北四个不同方向的枝条上采集长于树冠外、完全伸展、生长良好的叶子10 片(幼苗采集5 片),每个群落共采集长柄双花木叶片275 片。其中,成树100 片,幼树100 片,幼苗75 片。将采集到的长柄双花木叶片放置在两片湿润的滤纸中间,用自封袋保存。带回室内,进行测量。
1.3 叶功能性状指标的测定
本研究选择了长柄双花木叶面积、叶体积、叶干重以及叶含水量共四个指标进行测定,其中叶片面积反映了植物的光合捕获能力(Ackerly et al., 2002),叶片体积代表了植物的综合投资(Roderick et al., 2000),叶片含水量则能够反映叶片代谢的活跃程度(Roderick et al., 1999)。
将采集到的叶片置于水中,在5 ℃的黑暗条件下放置12 h, 取出后快速用滤纸吸干叶片表层水分,在精度为0.01 g的电子天平上称量得到饱和鲜重。用精度为0.01 mm 的电子游标卡尺,沿着主叶脉均匀选择3个点(每个点距叶片主脉约0.25 cm),测其厚度,取3个点的平均值作为一个叶片的厚度(Cornelissen et al., 2003)。用WinRHIZO根系分析系统(Regent Instruments Inc., 加拿大)扫描得到叶面积。将扫描后的叶片置于105 ℃下杀青15 min,于60 ℃下烘干至少72 h,称量获得叶片干重(Pérez-Harguindeguy et al., 2013)。其中,叶体积(LV)=叶面积(cm2)×叶厚度(cm),叶片含水量(LWC)=(叶片饱和鲜重-叶片干重)(g)/叶片饱和鲜重(g)×100%(Cornelissen et al., 2003)。
1.4 数据分析
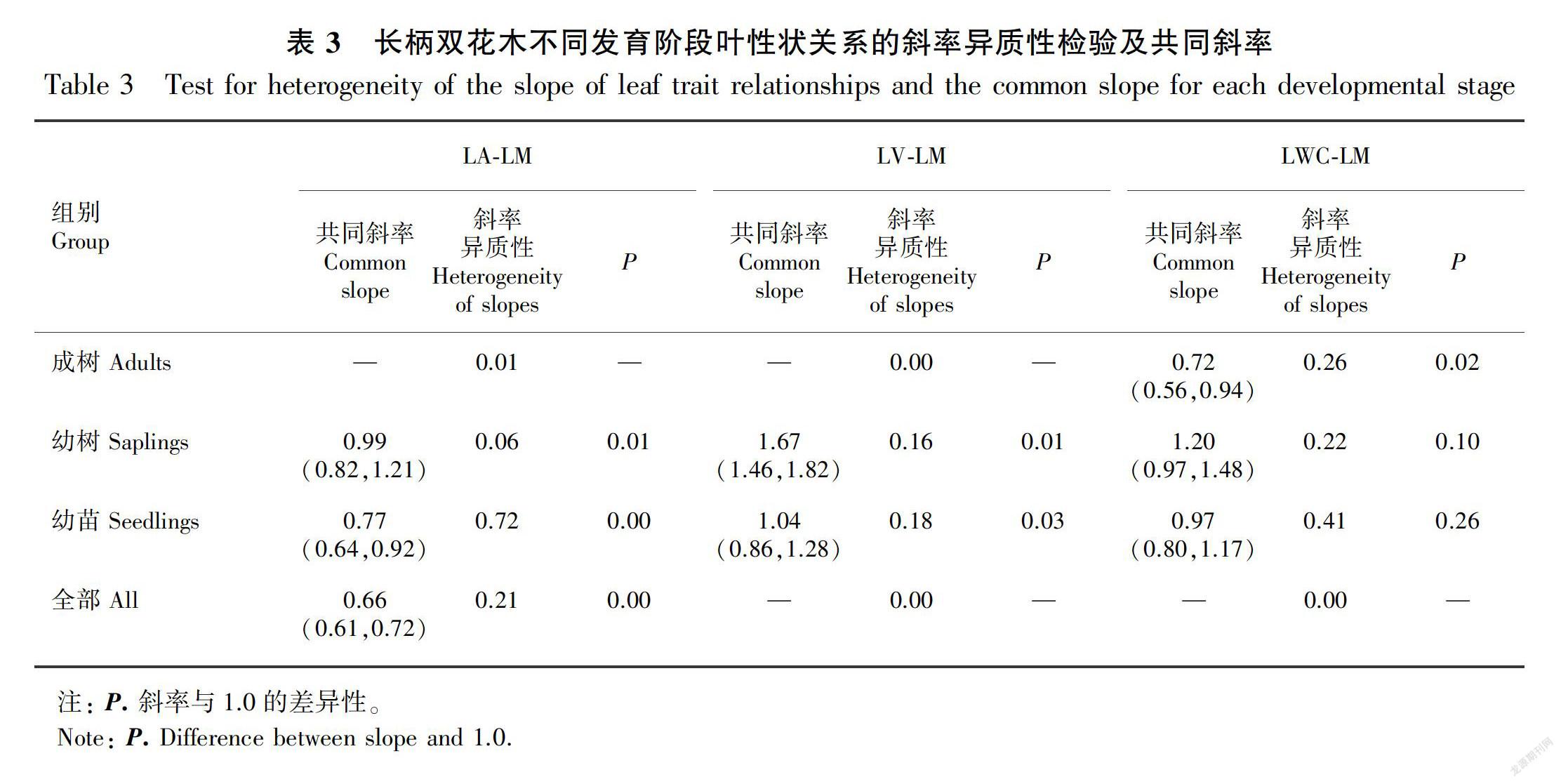
对各海拔每个个体的叶片性状求算术平均值,将其看作该个体叶片的性状值,然后将各叶片性状值以10为底进行对数转换,使其满足正态分布后进行分析。将异速生长方程式y=αxβ转换为对数表达式logy=logα+βlogx,其中x与y表示不同的叶性状值,β表示斜率,即异速生长指数。当β与1.0有显著性差异时,叶片性状间满足异速生长关系。斜率使用标准主轴回归(Standardized Major Axis,SMA)进行计算(Warton et al., 2006),并采用Pitman(1939)的方法计算出斜率的置信区间,同时检验斜率与1.0差异的显著性。使用Warton & Weber(2002)提出的方法计算斜率的异质性,当斜率同质时求出共同斜率(必要时需采用post hoc Tukey进行多重比较)。所有分析均在R软件(3.4.4)的smatr包中进行,所有统计检验的显著水平均使用α=0.05。
2 结果与分析
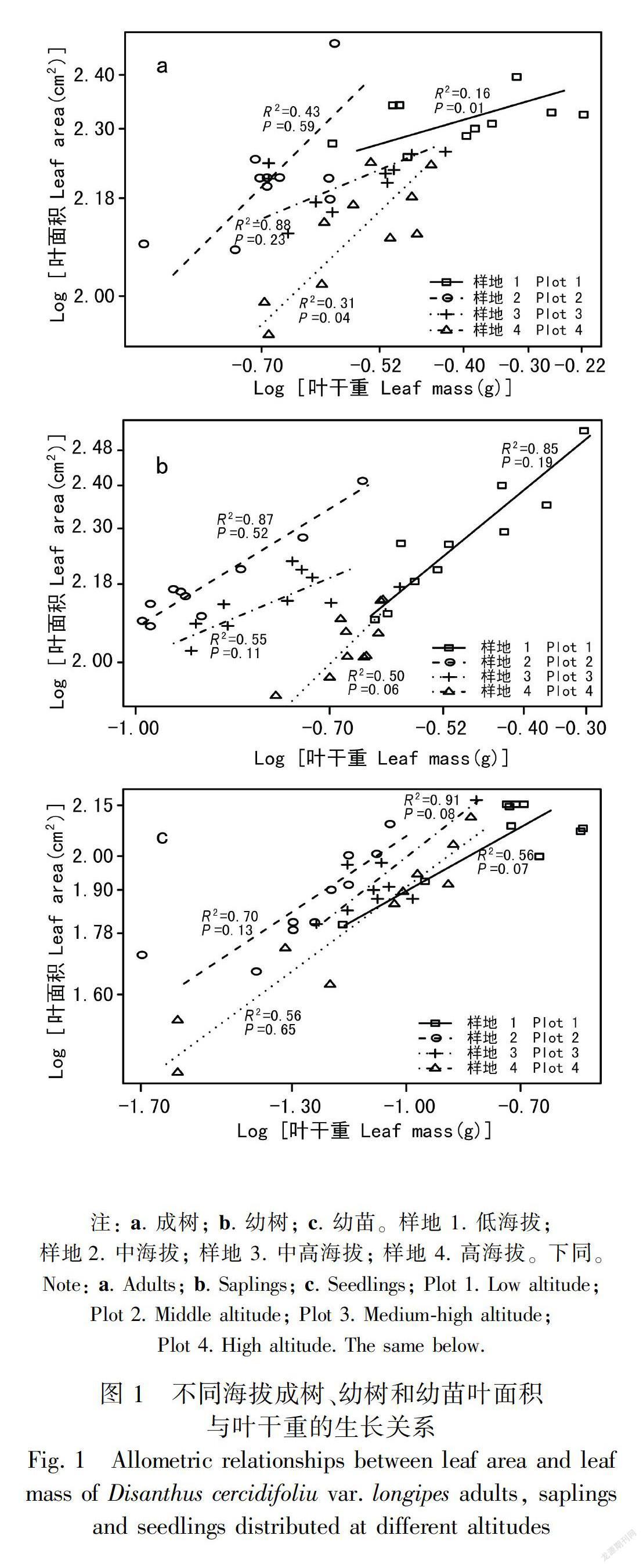
2.1 叶面积与叶干重的生长关系
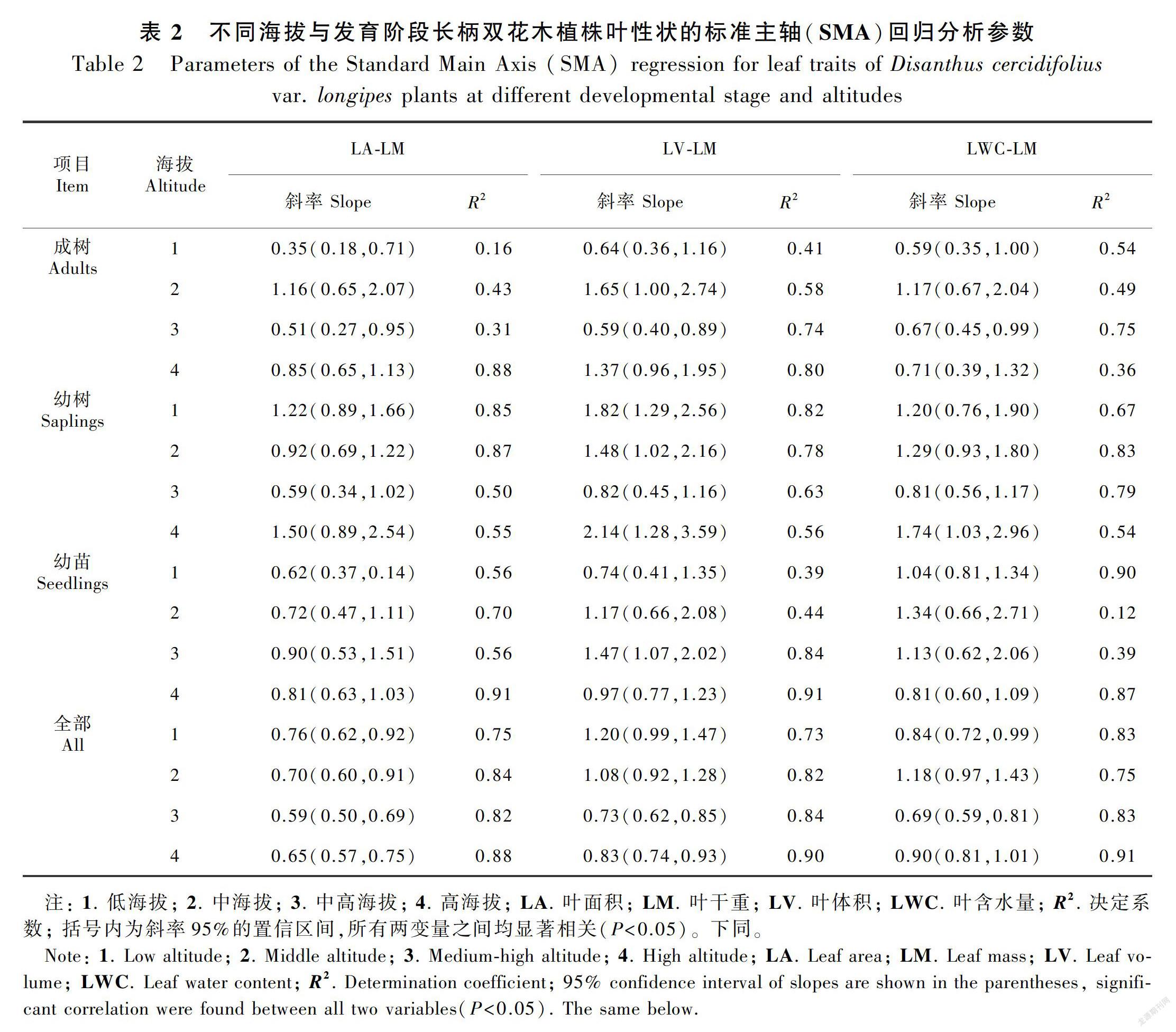
长柄双花木的叶面积与叶干重在不同发育阶段均表现为显著的相关关系(表2),但不同发育阶段植株叶面积与叶干重的异速生长状态随海拔梯度的变化而有所差异。成树叶面积与叶干重的斜率在不同海拔间有显著性差异(P<0.05)(图1,表3),其中低海拔和中高海拔的斜率差异不显著,共同斜率为0.43,与1.0差异显著(P<0.01),表现为异速生长关系;中海拔和高海拔的斜率差异不显著,共同斜率为1.17,与1.0无显著性差异(P>0.05),表现为等速生长关系;幼树和幼苗阶段的叶面积与叶干重的斜率在各海拔间无显著性差异,其中幼树的共同斜率为0.99(95%置信区间:CI=0.82~1.21),幼苗的共同斜率为0.77(95%置信区间:CI=0.64~0.92)(图1,表3),均与1.0差异显著(P<0.05),表现为异速生长关系。
不考虑发育阶段时, 不同海拔间长柄双花木的异速指数无显著性差异,共同斜率为0.66(95%置信区间:CI=0.61~0.72),与1.0差异显著(P<0.05),表现为异速生长关系(表3)。
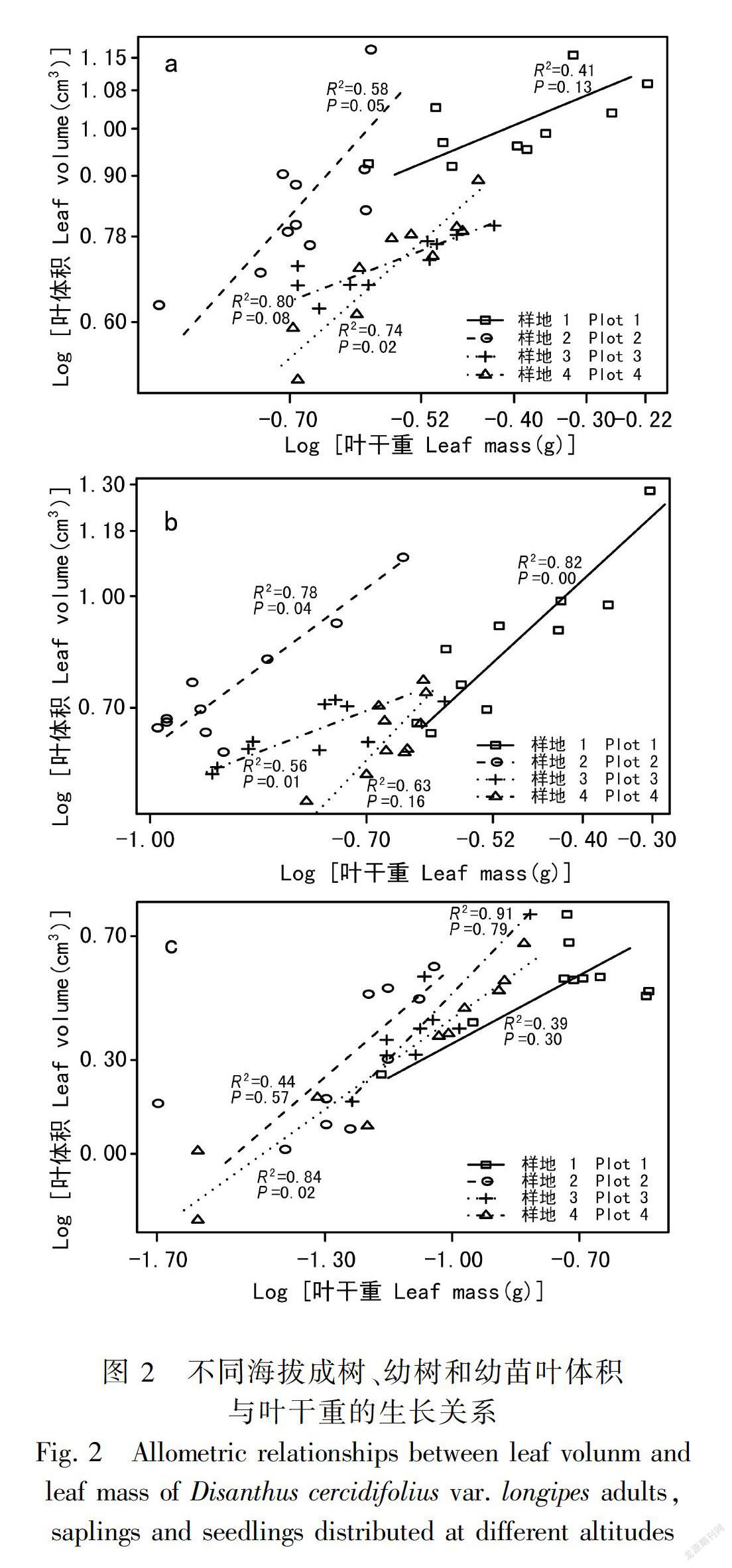
2.2 叶体积与叶干重的生长关系
长柄双花木的叶体积与叶干重在不同发育阶段均表现为显著的相关关系(表2),但不同发育阶段植株叶体积与叶干重的异速生长关系也随海拔梯度的变化而变化。长柄双花木成树的叶体积与叶干重的斜率在不同海拔间虽有显著性差异(P<0.05)(图2,表3),但四个海拔的异速指数分别与1.0无显著性差异(P>0.05),表现为等速生长关系(表2)。幼树和幼苗阶段的叶体积与叶干重的斜率在各海拔间无显著性差异,其中,幼树的共同斜率为1.67(95%置信区间:CI=1.46~1.82),幼苗的共同斜率为1.04(95%置信区间:CI=0.86~1.28)(图2,表3),均与1.0差异显著(P<0.05),表现为异速生长关系。
不区分发育阶段时,长柄双花木叶片体积与叶片干重的异速生长指数在不同海拔间有差异(表3)。其中低海拔和中海拔下叶体积与叶干重的异速指数无显著性差异(P>0.05),共同斜率为1.13,与1.0无显著性差异(P>0.05),呈等速生长关系;高海拔和中高海拔的异速指数无显著性差异(P>0.05),共同斜率为0.79,与1.0有显著性差异(P<0.01),体现出异速生长趋势(图2)。
2.3 叶含水量与叶干重的生长关系
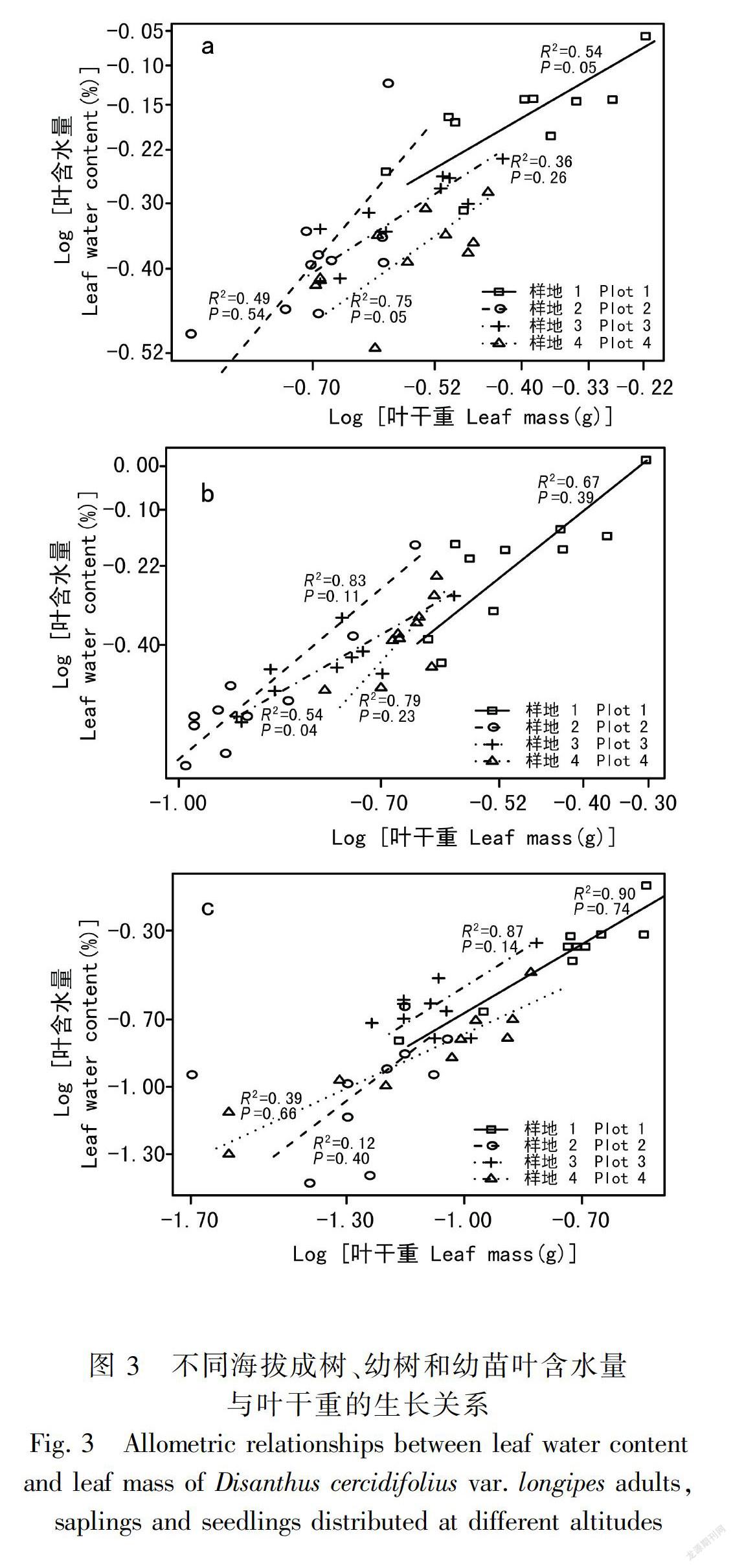
长柄双花木的叶含水量与叶干重在不同发育阶段均表现为显著的相关关系(表2),且每个发育阶段的异速指数在不同海拔间均无显著性差异(P>0.05)。成树的共同斜率为0.72(95%置信区间:CI=0.56~0.94),幼树的共同斜率为1.20(95%置信区间:CI=0.97~1.48),幼苗的共同斜率为0.97(95%置信区间:CI=0.80~1.17)(图3,表3)。其中成树的共同斜率与1.0差异显著(P<0.05), 呈异速生长关系;幼树与幼苗的异速指数与1.0无显著性差异(P>0.05),呈等速生长关系。也就是说,长柄双花木叶片含水量与叶片干重的异速生长关系表现为在成树阶段呈异速增长趋势且斜率小于1.0,在幼树与幼苗阶段呈等速增长趋势。
不区分发育阶段时,长柄双花木叶片含水量与叶片干重的异速生长指数在不同海拔间有差异(表3)。其中低海拔和中高海拔叶含水量与叶干重的异速指数无显著性差异,共同斜率为0.76,显著小于 1.0(P<0.01),体现出异速生长趋势;中海拔和高海拔的异速指数无显著性差异,共同斜率为0.88,与1.0无显著性差异(P>0.05),表现出等速生长关系。
3 讨论
3.1 叶性状异速生长关系与生物量投资策略随发育阶段的变化
植物异速生长关系能反映植物对资源利用方式和同化产物分配的特征(杨琼等,2016)。由于植物在不同生长发育阶段的生长发育的重点有所不同,因此植物性状异速生长关系与生物量投资策略常随着植株的发育而变化(姚婧等,2013)。植物以叶面积为基础进行光合作用,以叶体积为基础进行气体交换(Charles-Edwards,1979; Westoby et al., 2002),且叶片中的液态体积能表征叶片代谢的活跃程度(Roderick et al., 1999),因此叶面积、叶片体积和叶片含水量随叶干重变化的综合分析,能更好地掌握植物葉片的生长投资策略(祝介东等,2011)。杨琼等(2016)对胡杨的研究发现不同发育阶段植株叶性状间的异速生长关系不同。姚婧等(2013)对五角枫叶性状异速生长关系随发育阶段变化的研究表明五角枫植株叶资源利用方式在不同发育阶段发生了转变。
本研究中,长柄双花木叶面积与叶干重、叶体积与叶干重以及叶含水量与叶干重的异速生长状态和生物量投资策略在各发育阶段有较大差异。在幼苗和幼树阶段,叶面积与叶干重呈异速生长关系且斜率小于1.0,叶体积与叶干重呈异速增长趋势且斜率大于1.0,而叶片含水量与叶片干重呈等速增长趋势。若将叶片面积看作收益,叶片干重看作支出(祝介东等,2011),本发育阶段所出现叶面积与叶干重的异速生长关系表明叶片的收支不平衡。其原因是因为在资源不充足的林下环境中,长柄双花木幼苗和幼树的生长面临着与草本以及其他灌木植物的竞争。由于该物种是典型的喜荫喜湿植物,对光照要求较低,对水分需求较高(李晓红等,2013)。为了更有效地将水分输送到叶肉细胞以及更好地进行光合,该物种在幼苗和幼树会将较多的叶生物量用于投资建设维管组织(Poorter & Evans,1998,Garnier et al., 1999),而用于叶片同化的投资减少,空腔逐渐增大(Ishida et al., 2005)。最终导致叶片面积增长的速度小于叶片干重增加的速度,而叶体积的增长速度却大于叶干重的增长速度。在成树阶段,长柄双花木叶面积与叶干重在中海拔和高海拔下出现等速生长关系,叶体积与叶干重呈等速增长,叶片含水量与叶片干重呈异速增长趋势且斜率小于1.0。这可能是由于这一阶段长柄双花木需要同化大量养分输出到其他营养器官及生殖器官生长与构建。为了达到这一目的,植株将较多的叶生物量投资用于叶面积的增大和同化与支撑结构的构建,以提高叶片的光捕获能力,进而增加光合效率以获取更多的有机质(Ackerly et al., 2002;Ishida et al., 2005;Niinemets et al., 2007)。最终使得叶体积、叶面积与叶干重的增长速度相近,而叶含水量的增长低于叶干重。
3.2 叶性状异速生长关系随海拔的变化及可塑性
环境对叶表型变异有很大影响(张翠仙等,2014),海拔等环境因子的变化常引起植物叶性状异速生长关系发生变化(杨贺雨等,2016),而异速生长曲线的变化说明植物对环境变化具有最大程度上的可塑性(陆霞梅等,2007)。海拔主要影响环境的温度和光照(王志恒等,2004),进而对叶性状的异速生长关系及其生物量投资策略产生影响。杨贺雨等(2016)对华中五味子叶表型可塑性的研究表明不同海拔叶面积和叶柄长的可塑性较强,姚婧等(2013)对五角枫的研究表明不同海拔叶含水量的可塑性较强。
本研究中,不同海拔下长柄双花木叶面积与叶干重、叶体积与叶干重以及叶含水量与叶干重的异速生长状态在同一发育阶段基本没有显著差异。不同海拔下叶面积与叶干重、叶体积与叶干重以及叶含水量与叶干重的异速指数分别在幼苗和幼树阶段均没有显著差异,有共同斜率;不同海拔下叶含水量与叶干重的异速指数在成树阶段没有显著差异,有共同斜率。因而进一步研究了在不区分发育阶段的情况下,各异速生长关系随海拔的变化。在不区分发育阶段的情况下,对长柄双花木叶片各性状之间总体的异速生长关系的分析表明,不同海拔植株的叶面积与叶干重的异速指数无显著差异,但其叶体积和叶含水量分别与叶干重的异速生长指数在不同的海拔间都具有显著性差异。具体表现为叶体积与叶干重在低海拔和中海拔中呈等速增长趋势,在中高海拔和高海拔下叶体积的增长速度小于叶干重的增长速度;叶含水量的增长速度在低海拔和中高海拔下小于叶干重增长速度的趋势,在中海拔和高海拔中两者呈等速增长趋势。上述结果表明叶片体积和叶片含水量是可塑性较强的两个叶性状,它们对叶片干重的增加产生了可塑性响应。叶片体积代表了植株的综合投资(Roderick et al., 2000)。叶体积和叶干重异速生长关系在不同海拔的变化可能是由于不同海拔之间的环境条件的差异引起的。由于高海拔和中高海拔温度低,风力大,年降水量多,光强较强(Krner,2003),该海拔梯度的长柄双花木减少了对叶体积的投资,转而将更多的叶生物量投资于机械组织和维管组织,用于抵抗风力与强光。党晶晶等(2015)对甘肃臭草的研究也证明了光强越強,叶体积增长速度越慢。叶片含水量能够反映叶片代谢的活跃程度(Roderick et al., 1999),本研究中叶片含水量与叶片干重在不同海拔异速生长关系的变化表明植株叶片对代谢活性的投资可能也发生了改变。长柄双花木低海拔与中高海拔分布地土壤氮的含量较低,该海拔梯度植株叶氮含量也相对较低。由于叶氮含量与光合能力显著相关(Evans,1989),低海拔与中高海拔长柄双花木叶片相对较低的叶氮含量与光合能力可能使得其代谢活动相对减少,其叶含水量的增长速度相应地低于高海拔。
4 结论
本研究分析了不同发育阶段以及不同海拔长柄双花木植株的叶片面积、叶片体积以及叶片含水量与叶片干重之间的异速关系,发现不同发育阶段植株之间叶性状异速生长关系有着显著差异。成年树叶片面积的增长速度低于或等于叶片干重的增长速度,幼树、幼苗叶片面积的增长速度低于叶片干重的增长速度;成树叶片体积与叶片干重呈等速增长,幼树、幼苗叶片体积的增长速度高于叶干重的增长速度;成树叶片含水量的增长速度低于叶干重的增长速度,幼树、幼苗两性状间保持等速增长。同时确定了海拔梯度对长柄双花木叶性状异速生长关系也有影响,植株叶体积和叶含水量与叶干重的异速生长指数在不同海拔间有显著性差异。在低海拔和中海拔区域,叶体积与叶干重呈等速增长,叶含水量的增长速度低于叶片干重的增长速度。在中高海拔和高海拔区域,叶体积的生长速度低于叶干重的生长速度,叶含水量和叶片干重呈等速增长。本研究结果表明,长柄双花木叶片资源投资策略随着发育阶段和海拔梯度的不同而变化。成树主要将叶生物量投资于光捕获面积和同化结构,幼树和幼苗则主要投资于维管组织的建设。由于海拔升高导致风力增大、光强增强、土壤理化性质改变,因此长柄双花木在中低海拔倾向于增大叶体积以抢占资源,在高海拔倾向于加强机械组织和维管组织的建设。
致谢 感谢江西省井冈山国家级自然保护区管理局以及向琳、谭向前和张行等同志在野外调查中给予的帮助。
参考文献:
ACKERLY DD, KNIGHT CA, WEISS SB, et al., 2002. Leaf size, specific leaf area and microhabitat distribution of cha-parral plants: Contrasting patterns in species level and community level analyses [J]. Oecologia, 130(3): 449-457.
BERTALANFY LV (Translated by WU XJ), 1999. Problems of life: An evaluation of modern biological thought [M]. Beijing: The Commercial Press: 141-168.
CHARLES-EDWARDS DA, 1979. Photosynthesis and cropgrowth [M]// MARCELLE R, CLIJSTERS H, van POUCKE M. Photosynthesis and Plant Development. The Hague: Dr. W. Junk Publishers: 11-24.
CORNELISSEN JHC, LAVOREL S, GAMIER E, et al., 2003. A handbook of protocols for Standardized and easy measurement of plant functional traits worldwide [J]. Aust J Bot, 51(4): 335-380.
DANG JJ, ZHAO CZ, LI Y, et al., 2015. Relationship between leaf traits of Melica przewalskyi and slope aspects in alpine grassland of Qilian Mountains, China [J]. Chin J Plant Ecol, 39(1): 23-31.[党晶晶, 赵成章, 李钰, 等, 2015. 祁连山高寒草地甘肃臭草叶性状与坡向间的关系 [J]. 植物生态学报, 39(1):23-31.]
EVANS J, 1989. Photosynthesis and nitrogen relationships of leaves in C3 plants [J]. Oecologia, 78(1): 9-19.
FU LG, 1992. Chinese plant red book—rare and endangered plants(volume One) [M]. Beijing: Science Press: 324-325.[傅立国, 1992. 中国植物红皮书——珍稀濒危植物(第一册) [M]. 北京:科学出版社:324-325.]
GAO FX, LI MQ, ZHOU SX, et al., 2013. Distribution and current status of the endangered shrub Disanthus cercidifolius var. longipes [J]. Plant Sci J, 31(1): 34-41.[高浦新, 李美琼, 周赛霞, 等, 2013. 濒危植物长柄双花木(Disanthus cercidifolius var. longipes) 的资源分布及濒危现状 [J]. 植物科学学报, 31(1):34-41.]
GARNIER E, SALAGER JL, LAURENT G, et al., 1999. Relationships between photosynthesis, nitrogen and leaf structure in 14 grass species and theirdependence on the basis of expression [J]. New Phytol, 143(1): 119-129.
HE CX, LI JY, MENG P, et al., 2013. Changes of leaf traits and WUE with crown height of four tall tree species [J]. Acta Ecol Sin, 33(18): 5644-5654.[何春霞, 李吉躍, 孟平, 等, 2013. 4种高大树木的叶片性状及WUE随树高的变化 [J]. 生态学报, 33(18):5644-5654.]
HUEY RB, GILCHRIST GW, CARLSON ML, et al., 2000. Rapid evolution of a geographic cline in size in an introduced fly [J]. Science, 287: 308-309.
ISHIDA A, YAZAKI K, HOE AL, et al., 2005. Ontogenetic transition of leaf physiology and anatomy from seedlings to mature trees of a rain forest pioneer tree, Macaranga gigantean [J]. Tree Physiol, 25(5): 513-522.
KRNER C, 2003. Alpine plant life: Functional plant ecology of high mountain ecosystems[M]. 2nd ed. Springer Berlin, New York.
LA BARBERA M, 1989. Analyzing body size as a factor in ecology andevolution [J]. Annu Rev Ecol Syst, 20: 97-117.
LI XH, ZENG JJ, ZHOU B, 2013. Endangering reasons and conservation strategy of Disanthus cercidifolius var. longipes, an endemic plant to China [J]. J Jinggangshan Univ (Nat Sci Ed), 34(6):100-106.[李晓红, 曾建军, 周兵, 2013. 特有濒危植物长柄双花木濒危原因及其保育对策 [J]. 井冈山大学学报(自然科学版), 34(6):100-106.]
LI XQ, LI LK, XIE GW, et al., 2014. A study on the floristic diversity of the communities with national endangered plant Disanthus cercidifolius var. longipes [J]. Acta Agric Univ Jiangxi, 36(5): 948-957.[李象钦, 李丽卡, 谢国文, 等, 2014. 江西濒危植物长柄双花木群落区系多样性研究 [J]. 江西农业大学学报, 36(5):948-957.]
LIAO FY, 2010. The physiological characters of Disanthus cercidifolius and its application in the landscape and architecture [J]. N Hortic, (8): 69-71.[廖飞勇, 2010. 长柄双花木的生理习性及其在园林中的应用 [J]. 北方园艺, 8:69-71.]
LLEONART J, SALAT J, TORRES GJ, et al., 2000. Removing allometric effects of body size in morphological analysis [J]. J Theoret Biol, 205: 85-93.
LU XM, ZHOU CF, AN SQ, et al., 2007. Phenotypic plasticity, allometry and invasiveness of plants [J]. Chin J Ecol, 9:1438-1444.[陆霞梅, 周长芳, 安树青, 等, 2007. 植物的表型可塑性、异速生长及其入侵能力 [J]. 生态学杂志, 9:1438-1444.]
MCCONNAUGHAY KDM, COLEMAN JS, 1999. Biomass allocation inplants:Ontogeny or optimality? A test along three resource gradients [J]. Ecology, 80: 2581-2593.
MILLA R, REICH PB, 2007. The scaling of leaf area and mass: The cost of light interception increases with leafsize [J]. P Roy Soc Lond B Bio, 274(1622): 2109-2114.
NIINEMETS, PORTSMUTH A, TENA D, et al., 2007. Do we underestimate the importance of leaf size in plant economics? Disproportional scaling of support costs within the spectrum of leaf physiognomy [J]. Ann Bot, 100(2): 283-303.
P-REZ-HARGUINDEGUY N,D AZ S, GARNIER E, et al., 2013. New handbook for standardised measurement of plant functional traits worldwide [J]. Aust J Bot, 61: 167-234
PIGLIUCCI M, 2001. Phenotypic plasticity: Beyond nature andnurture [M]. Baltimore and London: Johns Hopkings University Press.
PITMAN ETG, 1939. A note on normal correlation [J]. Biometrika, 31(1/2): 9-12.
POORTER H, EVANS JR, 1998. Photosynthetic nitrogen-use efficiency of species that differ inherently in specific leafarea [J]. Oecologia, 116(1/2): 26-37.
RODERICK ML, BERRY SL, NOBLE IR,2000. A framework for understanding the relationship between environment and vegetation based on the surface area to volume ratio of leaves [J]. Funct Ecol, 14: 423-437.
RODERICK ML, BERRY SL, NOBLE IR, et al., 1999. A theoretical approach to linking the composition and morpho-logy with the function of leaves [J]. Funct Ecol, 13(5): 683-695.
WANG ZH, CHEN AP, PIAO SL, et al., 2004. Pattern of species richness along an altitudinal gradient on Gaoligong Mountains, Southwest China [J]. Biodivers Sci, 12: 82-88.[王志恒, 陳安平, 朴世龙, 等, 2004. 高黎贡山种子植物物种丰富度沿海拔梯度的变化 [J]. 生物多样性, 12:82-88.]
WARTON DI, WEBER NC, 2002. Common slope tests forbivariate errors-in-variables models [J]. Biom J, 44(2): 161-174.
WARTON DI, WRIGHT I, FALSTER DS, et al., 2006. Biva-riate line-fitting methods for allometry [J]. Biol Rev, 81(2): 259-291.
WEI L, 2015. Disanthus cercidifolius var. longipes [J]. Bull Biol, 50(5): 3.[魏來, 2015. 长柄双花木 [J]. 生物学通报, 50(5):3.]
WEINER J, 2004. Allocation, plasticity andallometry in plants [J]. Perspect Plant Ecol, 6(4): 207-215.
WEST GB, BROWN JH, 2005. The origin of allometric scaling laws in biology from genomes to ecosystems: Towards a quantitative unifying theory of biological structure and organization [J]. J Exp Biol, 208: 1575-1592.
WESTOBY M, FALSTER DS, MOLES AT, et al., 2002. Plant ecological strategies: Some leading dimensions of variation between species [J]. Ann Rev Ecol Syst, 33: 125-159.
WRIGHT J, REICH PB, WESTOBY M, et al., 2004. The worldwide leaf economicsspectrum [J]. Nature, 428: 821-827.
XIAO YA, HE P, LI XH, et al., 2004. Study on numeric dynamics of natural populations of the endangered species Disanthus cercidifolius var. longipes [J]. Chin J Plant Ecol, 28(2): 252-257.[肖宜安, 何平, 李晓红, 等, 2004. 濒危植物长柄双花木自然种群数量动态 [J]. 植物生态学报, 28(2):252-257.]
XIE GW, LI LK, ZHAO JJ, et al., 2014. Genetic diversity of national endangered plant Disanthus cercidifolius var. longipes in Nanling [J]. J Guangzhou Univ(Nat Sci Ed), 13(4): 47-52.[谢国文, 李丽卡, 赵俊杰, 等, 2014. 南岭国家濒危植物长柄双花木遗传多样性研 [J]. 广州大学学报(自然科学版), 13(4):47-52.]
XIE GW, XU HM, LI XQ, et al., 2016. The species community diversity of the endangered plant Disanthus cercidifolius var. longipes in Jiangxi and Zhejiang Province [J]. J Guangzhou Univ(Nat Sci Ed), 15(4): 33-38.[谢国文, 徐惠明, 李象钦, 等, 2016. 濒危植物长柄双花木赣浙群落的物种多样性研究 [J]. 广州大学学报(自然科学版), 15(4):33-38.]
YANG GY, YU WT, ZHENG J, et al., 2018. Scanning electron microscopy observation on foliar micromorphology of Oolong tea germplasms [J]. J S Agric, 49(10):2020-2027. [杨国一, 于文涛, 郑晶, 等,2018. 乌龙茶种质叶片微形态特征的扫描电镜观察 [J]. 南方农业学报, 49(10):2020-2027.]
YANG HY, WEI HY, SANG MJ, et al., 2016. Phenotypic plasticity of Schisandra sphenanthera leaf and the effect of environmental factors on leaf phenotype [J]. Chin Bull Bot, 51(3): 322-334.[杨贺雨, 卫海燕, 桑满杰, 等, 2016. 华中五味子叶表型可塑性及环境因子对叶表型的影响 [J]. 植物学报, 51(3):322-334.]
YANG Q, LI ZZ, FU Q, et al., 2016. Relationship among leaf trait and developing process in Populus euphratica [J]. J Desert Res, 36(3): 659-665.[杨琼, 李征珍, 傅强, 等, 2016. 胡杨(Populus euphratica)叶异速生长随发育的变化 [J]. 中国沙漠, 36(3):659-665.]
YAO J, LI Y, WEI LP, et al., 2013. Changes of allometric relationships among leaf traits in different ontogenetic stages of Acer mono from different types of forests in Donglingshan of Beijing [J]. Acta Ecol Sin, 33(13): 3907-3915.[姚婧, 李颖, 魏丽萍, 等, 2013. 东灵山不同林型五角枫叶性状异速生长关系随发育阶段的变化 [J]. 生态学报, 33(13):3907-3915.]
ZHANG CX, NI ZG, CHEN HR, et al., 2014. Genetic diversity of mango (Mangiferca indica L. ) in nujiang dry-hot valley revealed by morphological characters and AFLP marker [J]. J plant genet resour, 15(4): 753-758.[張翠仙, 尼章光, 陈华蕊, 等, 2014. 怒江干热河谷杧果种质资源的表型和AFLP遗传多样性分析 [J]. 植物遗传资源学报, 15(4):753-758.]
ZHANG JM, LIAO YY, XIE GW, et al., 2013. Population characteristics of national rare and endangered plant Disanthus cercidifolius var. longipes [J]. J Trop Biol, 4(1): 74-80.[张嘉茗, 廖育艺, 谢国文, 等, 2013. 国家珍稀濒危植物长柄双花木的种群特征 [J]. 热带生物学报, 4(1):74-80.]
ZHU JD, MENG TT, NI J, et al., 2011. Within-leaf allometric relationships of mature forests in different bioclimatic zones vary with plant functional types [J]. Chin J Plant Ecol, 35(7): 687-698.[祝介东, 孟婷婷, 倪健, 等, 2011. 不同气候带间成熟林植物叶性状间异速生长关系随功能型的变异 [J]. 植物生态学报, 35(7):687-698.
ZHU SY, YAN LB, PI FJ, et al., 2019. Stoichiometric characteristics and seasonal variation of soils and dominant plant leaves in secondary forest in karst area [J]. J S Agric, 50(1):90-96. [朱恕英,严令斌,皮发剑,等,2019. 喀斯特区次生林土壤和优势树种叶片的化学计量特征及季节变化 [J]. 南方农业学报, 50(1):90-96.]
