乌蒙山峨眉栲、华木荷群落特征研究
2019-09-10周杰彭明春王崇云曹文馨等
周杰 彭明春 王崇云 曹文馨 等


摘 要:乌蒙山位于中国西部半湿润常绿阔叶林与东部湿润常绿阔叶林交界区,是云南省唯一分布峨眉栲、华木荷群系( Formation Castanopsis platyacantha,Schima sinensis)的地区,该群系处于中国西部常绿阔叶林向东部常绿阔叶林过渡的地带,群落更加复杂多样。为了更深入地了解该群系的群落特征,同时找出其与东部湿润常绿阔叶林的联系,该文对该类型群落进行了数量分类研究,分析了聚类归并的各群落类型的乔灌优势种特征(径级结构、高度结构和重要值),同时采用DCA,CCA和DCCA排序分析进行群落特征分异的环境解释,并与四川峨眉山同类群落进行特征对比。结果表明:(1)乌蒙山峨眉栲、华木荷群落可划分为6个类型:峨眉栲群落,峨眉栲、华木荷群落,峨眉栲、十齿花群落,水青冈(Fagus longipetiolata)、华木荷、峨眉栲群落,华木荷、硬斗石栎(Lithocarpus hancei)群落,十齿花(Dipentodon sinicus)、华木荷群落。(2)乌蒙山峨眉栲、华木荷群落中,峨眉栲大树比例较高,华木荷小苗数量巨大,灌木層常以竹类为优势种。(3)海拔、坡度和太阳年辐射值是影响群落分布的主导因素。(4)峨眉山同类群落中优势种种群呈稳定型或增长型,而乌蒙山峨眉栲、华木荷群落中,峨眉栲和水青冈呈现下降型的特点。
关键词:乌蒙山,峨眉栲,华木荷群落,聚类分析,排序
Abstract:Wumeng Mountain is located in the joint zonation between western semi-humid evergreen broad-leaved forest and eastern humid evergreen broad-leaved forest in China. Formation Castanopsis platyacantha, Schima sinensis is a typical forest in this area with complex and diverse types that are only occurrence of Yunnan. In order to understand the community characteristics of this formation in more details,and to find out its relationship with the eastern moist evergreen broad-leaved forest,in this paper,we made clustering analysis on the basis of canopy and shrub layer species. And then,we analyzed the dominant species in tree and shrub layers by DBH class structure,height class structure,and importance value,and carried out ordination analysis by DCA,CCA,and DCCA to reveal the relationship between community features and environmental gradients. Finally,we compared the communities with the similar ones in Emei Mountain. The results were as follows:(1) Castanopsis platyacantha,Schima sinensis community in Wumeng Mountain could be divided into six types:Castanopsis platyacantha community,Castanopsis platyacantha,Schima sinensis community,Castanopsis platyacantha,Dipentodon sinicus community,Fagus longipetiolata,Schima sinensis,Castanopsis platyacantha community,Schima sinensis,Lithocarpus hancei community,and Dipentodon sinicus,Schima sinensis community. (2) There was a large proportion of Castanopsis platyacantha trees in old age,and a great number of seedlings of Schima sinensis in the community. Meanwhile,bamboo species were tyipical dominant in the shrub layer. (3) Community patterns were mainly influenced by elevation,slope,and annual solar radiation. (4) Dominate tree species’ population structures of similar communities in Emei Mountain were stationary or expansive type,but those were characteristics of constrictive type in Wumeng Mountain.
Key words:Wumeng Mountain,Castanopsis platyacantha,Schima sinensis community,clustering analysis,ordination analysis
中国是世界上常绿阔叶林分布最辽阔的区域,类型也最为多样。云南的常绿阔叶林因受西南季风和高原地貌的深刻影响,其生态表现和种类组成与我国东部的常绿阔叶林既有紧密的联系,又有显著的差别(金振洲,1979;吴征镒和朱彦丞,1987)。中山湿性常绿阔叶林是云南山地植被垂直带的主要类型之一,主要分布于哀牢山、无量山、镇康大雪山、高黎贡山、乌蒙山中部和北部等地,与云南的水平地带性植被半湿润常绿阔叶林和季风常绿阔叶林不同,这一植被亚型以“湿”为主要特点,属于西部典型常绿阔叶林中的西部山地柯集群(宋永昌,2013)。
乌蒙山保护区位于云南省东北部,地处四川盆地的边缘,恰好处在西部半湿润常绿阔叶林向东部湿润常绿阔叶林过渡的地带(Song,1988)。这一区域的主要植被类型属于金振洲划分的“中山湿性常绿阔叶林”(金振洲,1979),但与无量山(彭华和吴征镒,1998)、高黎贡山(孟广涛等,2013)、哀牢山(巩合德等,2011)、澜沧江保护区(丁涛等,2006)等地的中山湿性常绿阔叶林不同,乌蒙山自然保护区是我省唯一分布峨眉栲、华木荷为优势种的常绿阔叶林地区,也是我国峨眉栲、华木荷群落分布的最西边缘。
峨眉栲、华木荷群落在湖北、重庆、贵州、四川等地均有分布,是西部常绿阔叶林向东部常绿阔林过渡的类型,在种类上兼有两者特点(宋永昌,2004),该群落是四川亚热带常绿阔叶林最主要的类型之一(四川植被协作组,1980),云南主要分布于滇东北大关、永善、彝良、镇雄等地。这类植被在云南垂直分布可达海拔2 300 m(杨一川等,1994)。《云南植被》中已有乌蒙山峨眉栲、华木荷群落的相关记载,但在《中国植被志》栲林、木荷林植被志编研工作中,发现以往资料无法满足编研需要,所以对这一类型进行了更广泛的深入调查,从数量分类、群落排序、优势种种群结构等方面进一步揭示群落特征和环境分异规律,并与四川峨眉山同类群落进行对比,为乌蒙山自然保护区峨眉栲、华木荷群落的管理与保護提供理论指导。
1 研究区概况
乌蒙山国家级自然保护区位于云南省东北部昭通市境内,在云贵川三省交界处,地跨大关、彝良、盐津、永善和威信5县,总面积为26 186.65 hm2,由三江口、朝天马、海子坪三个片区组成。地理位置介于103°51′47″—104°45′04″ E和27°47′35″—28°17′42″ N之间,其主要保护对象为亚热带湿性常绿阔叶林森林生态系统、珍稀濒危特有物种及其栖息地、云贵高原湿地代表类型,属于森林生态系统类型自然保护区。
乌蒙山地势崎岖不平,坡度大多在20°左右,属于深至中等切割的中山山地。年均温为11~18 ℃,年降水量在1 000~1 300 mm 之间,降水量北部多于南部,迎风坡多于背风坡。区内日照时数约为 1 000 h,年日照率约为 25%,无霜期为210~220 d。乌蒙山是四川盆地冷空气进入云南的主要通道,在昆明准静止锋的控制下,气候与云南大部分地区不同,光照少、阴雨日数多,湿度大,干湿季分明但干季不干。
2 研究方法
2.1 群落调查
2.1.1 样地位置的确定 据文献记载和当地林业局提供的资料,实地调查典型的峨眉栲、华木荷群落,调查地为彝良县山羊坪、高木桥、横河头上,大关县罗汉坝、三江口、小岩坊附近,镇雄县首家坪子、场坝罗汉,地理位置介于103°55′—104°42′ E和27°24′—28°15′N之间,共调查400~900 m2的样地26个。样地信息见表1。
2.1.2 样地设置和记录 根据具体的地形情况采用最小面积法(Kikvidze & Ohsawa,2002;王睿智和国庆喜,2016)设置样地,记录样地内的坡度、坡向、海拔和经纬度。乔木树种在1.3 m以上,测量每株基径、胸径、冠幅、株高、干高等数据,1.3 m以下的记入灌木层中;灌木和草本从样地4个角落各取2 m × 2 m的小样地,计录物种的株高和丛径,每个样地都采集了标本,野外未识别出的物种带回室内进行鉴定。
2.2 分析方法
2.2.1 相对基部面积(relative basal area,RBA) 物种相对基部面积=该物种基部面积之和/所有物种基部面积之和(Tang et al.,2011);乔木树种高度在1.3 m以上的记作乔木,1.3 m以下的记作灌木;灌木层根据高度进行基径的划分,高度0.5 m以下的基径记为0.5 cm,0.5 m以上的基径记为1 cm。
2.2.2 群落聚类 运用SYN-TAX2000的层次聚类(hierarchical classification)模块进行群落聚类分析。层次聚类主要通过一定规则自下而上的对样地进行合并,并在达到一定条件时终止合并。以样地编号为行,物种相对基部面积为列进行汇总,得到群落数据矩阵,将矩阵输入SYN-TAX2000软件进行聚类分析,并绘制聚类分析树状图。
2.2.3 径级结构、高度结构和重要值 依据群落聚类分析的结果,对同一组群落的乔木层和灌木层分别计算物种的相对基部面积(RBA),然后对每一组乔木层的前3个优势物种进行径级结构、高度级结构分析,灌木层的前三个优势物种进行重要值分析。(1)径级结构(diameter at breast height,DBH)。按照“1960年森林专业调查办法(草案)”中的规定:林分平均直径在12 cm 以上时,以4 cm及以下为第I级,胸径大于24 cm的划为第Ⅶ级,单位为株·hm-2。(2)高度结构。根据样地的具体情况,高度划分五级:1.3 m及以下;1.3~8 m(含8);8~12 m(含12);12~16 m(含16);16 m以上,单位为株·hm-2。(3)重要值。选取RBA最大的3个物种作为灌木层优势种,重要值=(相对密度+相对频度+相对显著度)/3。相对密度=该种的株数/总的株数;相对频度=该种出现的样地数/总的样地数;相对显著度=该种的RBA /所有种的RBA和。
2.2.4 数量排序 用Canoco for windows 4.5进行数量排序分析。以相对基部面积矩阵进行去趋势对应分析(detrended correspondence analysis,DCA)分析。用1∶5万DEM栅格数据提取土壤水分饱和度、土壤稳定性(Robert et al.,2005)和太阳年辐射值(Mccune & Keon,2002),加上样地采集的海拔、坡度、坡向数据,创建环境因子矩阵,结合相对基部面积矩阵进行典范对应分析(canonical correspondence analysis,CCA)和去趋势典范对应分析(detrended canonical correspondence analysis,DCCA)。
3 结果与分析
3.1 聚类分析及优势种分析
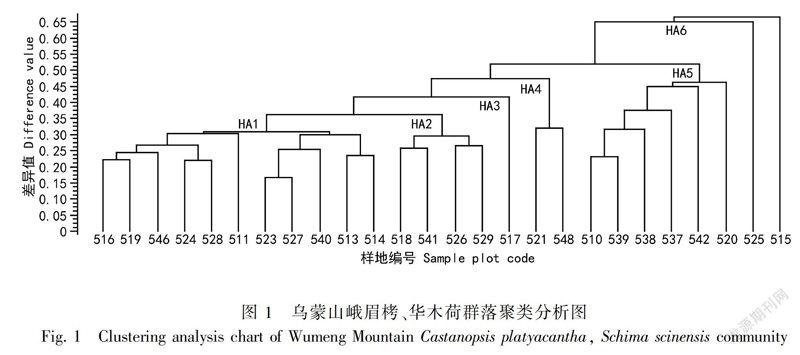
3.1.1 群落聚类 以各样地乔木和灌木物種的相对基部面积(RBA)作为分类依据,通过SYN-TAX2000进行层次聚类分析,以差异值0.35为起始,逐层向上聚类,到差异值0.65停止聚类,共得到了6个类别。聚类结果按由小到大的顺序进行显示,其结果如图1所示。
所有样地共分为6组。HA1:峨眉栲群落,包括516、519、546、524、528、511、523、527、540、513、514号样地;HA2:峨眉栲、华木荷群落,包括518、541、526、529号样地;HA3:峨眉栲、十齿花群落,仅有517号样地;HA4:水青冈、华木荷、峨眉栲群落,包括521、548号样地;HA5:华木荷、硬斗石栎群落,包括510、539、538、537、542、520号样地;HA6:十齿花、华木荷群落,包括525、515号样地。
3.1.2 乔木层和灌木层优势种分析 聚类分析划分出来的6组群落的乔木层物种RBA见表2,灌木层优势种RBA见表3。从表2可以看出,聚类分析所得的6个群落类型都以峨眉栲或者华木荷为优势种,除了这两个优势种外,水青冈(HA4水青冈、华木荷、峨眉栲群落)、十齿花(HA6十齿花、华木荷群落)也会有超过它们的情况;其余乔木物种中出现次数较多且RBA较大的有:硬斗石栎、薄叶山矾(Symplocos anomala)、海桐山矾(S. heishanensis)、峨眉柯(Lithocarpus oblanceolatus)、五裂槭(Acer oliverianum)、细梗吴茱萸叶五加(Acanthopanax evodiaefolius var. gracilis)、木瓜红(Rehderodendron macrocarpum)、粗脉杜鹃(Rhododendron coeloneurum)、木姜子(Litsea pungens)、珍珠花(Lyonia ovalifolia)、西南山茶(Camellia pitardii)、房县槭(Acer franchetii)、叶萼山矾(Symplocos phyllocalyx)、康定冬青(Ilex franchetiana)等。从表3可以看出,筇竹(Qiongzhuea tumidinoda)、箭竹(Disporum cantoniense)和方竹(Chimonobambusa quadrangularis)在5类群落中都占了绝对的优势,HA3峨眉栲、十齿花群落中红叶木姜子(Litsea rubescens)占了绝对优势。竹类要么单优,要么混合占有优势。在红叶木姜子为优势的HA3群落中,没有竹类植物。除了竹类和红叶木姜子,其余灌木物种中出现次数较多且RBA较大的物种有:五裂槭、西南绣球(Hydrangea davidii)、野八角(Illicium majus)、华木荷、峨眉栲、多变石栎(Lithocarpus variolosus)、西南山茶、薄叶山矾、木姜子、散毛樱桃(Cerasus patentipila)、滇瑞香(Daphne feddei)。华木荷和峨眉栲的幼苗都出现在4类群落中,而且华木荷幼苗的RBA略高于峨眉栲。
3.2 径级结构、高度结构和重要值分析
乔木层优势种的径级结构和高度结构从水平和垂直两个方向上反映了优势种在群落中的组合情况以及数量动态及其发展趋势,灌木层优势种的重要值则能进一步反映其在群落中的优势程度。
3.2.1 乔木层优势种的径级结构 乔木层优势种(RBA最大的3种)的径级结构如图2所示。从图2可以看出,峨眉栲第Ⅶ级数量最多,第Ⅱ级基本没有,第Ⅰ、第Ⅲ~第Ⅵ级有一定数量;华木荷第Ⅰ级数量最多,在HA5中甚至接近600株·hm-2,其余各级都相对较少;薄叶山矾主要集中在第Ⅱ~第Ⅵ级,第Ⅰ、第Ⅶ级都没有;五裂槭、硬斗石栎和野核桃都是第Ⅰ级数量最多,其余各级都相对较少;水青冈在HA2中各级数量都较少,在HA4中第Ⅶ级数量最多;十齿花第Ⅳ级数量最多,其余各级均较少。
3.2.2 乔木层优势种的高度结构 乔木层优势种(RBA最大的3种)的高度结构如图3所示。从图3可以看出,峨眉栲从低到高数量总体呈上升趋势,其中16 m以上大树数量最多;华木荷1.3 m以下幼苗最多,其余各高度数量分布均匀;五裂槭、硬斗石栎、野核桃都是1.3 m以下数量最多,只有野核桃没有16 m以上大树; 水青冈除了1.3 m以下数量较少外,其余各高度级数量均较多,其中1.3~8 m数量最多;十齿花高度主要集中在12 m以下,其中1.3~8 m数量最多。
3.2.3 灌木层优势种的重要值分析 从图4可以看出,RBA和重要值基本成正比,即RBA增加,重要值也随之增加,RBA减少,重要值也减少;所有类型群落除HA3外都是竹类为主要优势种,竹类要么混合占据优势,要么单独占据优势;当竹类混合占据优势时,优势种间的差距相对较小,当竹类单独为优势时,其优势程度则更加明显;在没有竹类的HA3群落中,红叶木姜子占优势,且优势程度也很明显。
3.3 DCA排序分析
对26个样地进行DCA排序分析,前四个排序轴的特征值分别为0.773、0.462、0.284、0.185。第一二轴特征值较大,说明这两个排序轴包含的生态信息量多,具有更重要的生态意义。以前两轴为轴绘制二维排序图,结果如图5。从图5可以看出,样地在第二轴上有明显的梯度变化,说明DCA排序较好的反映了各样地之间及样地与环境之间的关系。从排序轴来看,第一轴基本反映了水分梯度变化,从左到右为湿润到干旱的过渡(王景升等,2016),第二轴反映了温度梯度变化(海拔),即从下往上,海拔逐渐升高,年均温逐渐降低。由此看出海拔对群落分布影响最大,DCA排序结果与聚类所得结果基本一致。HA6为次生性群落,与干扰相关。
3.4 CCA排序分析
典范对应分析(CCA)是一种基于单峰模型的排序方法。它可将样地、物种及环境因子排序表示在一个图上,從而直观地看出它们之间的关系(张金屯,2004)。对26个样地和6个环境因子进行CCA排序分析,结果如图6所示。
排序图中箭头表示环境因子,箭头越长表示其对研究对象的分布影响越大;箭头连线与排序轴夹角余弦值代表其与排序轴的相关性大小。从图6中可以看出,6个环境因子中海拔对群落的分布影响最大,坡度和太阳年辐射值次之,坡向最小;海拔与第一排序轴具有最大的相关性,坡度则和第二排序轴有较强的相关性;将样地投影于海拔变量延长线上可以看出明显的梯度关系,HA5所在区域海拔最高,HA1整体分布的海拔最低;HA1分布的区域坡度较大,而HA5华木荷、硬斗石栎群落所在区域坡度相对较小。HA6属于次生生境,相对较为开阔,因此太阳辐射值也更强。CCA排序结果与聚类结果基本一致,相比DCA排序而言能更清楚地展示群落与各环境因子之间的关系。
3.5 DCCA排序分析
DCCA采用与DCA相同的除趋势方式,但去除了弓形效应的影响,可以看成是CCA和DCA的结合(张金屯,2004)。对样地和环境因子做DCCA分析,结果如图7所示。
从图7中可以看出,海拔对群落的分布影响最大,坡度和太阳年辐射值次之,坡向最小,整体结果与CCA相似,但样地的分离度不如CCA。
4 讨论与结论
乌蒙山峨眉栲、华木荷群落在《云南植被》中属于中山湿性常绿阔叶林,但这一群落在地理分布和物种组成等方面都更接近东部的湿润常绿阔叶林,因此可以看作是湿润常绿阔叶林。从生境气候、聚类分析、优势种种群三个方面看,和峨眉山同类型群落相比既有联系,又有区别。
(1)气候比较:本研究中,乌蒙山峨眉栲、华木荷群落平均海拔在2 000 m左右,因此选择海拔最接近的镇雄县作为气候比较的代表,峨眉山则以峨眉山市为代表。乌蒙山和峨眉山海拔相差500 m左右;年均温相差不大,但是乌蒙山月平均温度均未低于0 ℃,峨眉山12、1、2、3月均温都低于0 ℃;峨眉山的年降水量差不多是乌蒙山的一倍多,两地降水都主要集中在夏季(图8)。
(2)群落聚类分析:依据峨眉山峨眉栲、华木荷群落特征研究(杨一川等,1994),对峨眉山峨眉栲、华木荷群落所有样地进行层次聚可分为3组:EH1.峨眉栲、华木荷群落,包括10、9、8、7号样地;EH2.峨眉栲群落,包括6、5、1号样地;EH3.华木荷群落,包括4、2、3号样地(图9)。
由此可见,相比峨眉山同类群落,乌蒙山峨眉栲、华木荷群落乔木层优势树种种类更多,群落分化更明显,这可能是乌蒙山地处东西部常绿阔叶林过渡区域的原因。
(3)群落优势种种群对比:种群是构成群落的基本单位,其结构能客观反映出群落的发展演变趋势,本研究中,将乌蒙山所有群落进行合并,得出的乌蒙山峨眉栲、华木荷群落中4个主要优势种种群的高度和径级结构。
从高度结构看,乌蒙山华木荷种群高度主要集中在1.3 m及以下,峨眉山华木荷种群则集中在6~22 m,两地16 m以上大树都有一定数量;乌蒙山峨眉栲种群高度主要集中在12~16 m,峨眉山峨眉栲种群高度则在4 m和16 m两个高度级;乌蒙山另外两个优势种是水青冈和十齿花,这两个种高度主要集中在1.3~8 m,十齿花没有16 m以上的大树,峨眉山另外的优势种是山矾(Symplocos
caudata)和峨眉四照花(Dendrobenthamia capitata),其中山矾多数个体仅高2~6 m,峨眉四照花可达16 m(图10)。
从径级结构看,乌蒙山华木荷种群是典型的增长型种群,峨眉山的华木荷种群是增长型或稳定型;乌蒙山峨眉栲群落第Ⅶ级比率最大,属于下降型种群,峨眉山的峨眉栲种群则是增长型或稳定型;乌蒙山水青冈种群属于下降型种群,十齿花种群则属于增长型或稳定型,峨眉山的山矾种群是稳定型,西南山茶(Camellia pitardii)种群为增长型、峨眉四照花、油樟(Cinnamomum longepaniculatum)种群可能成为增长型或稳定型(图10)。
综上所述,乌蒙山峨眉栲种群大树较多呈现出下降型种群特点,华木荷种群幼苗数量巨大,种群呈增长型;峨眉山的峨眉栲和华木荷种群则都处于增长型或稳定型阶段。
乌蒙山峨眉栲、华木荷群落具有以下特征:(1)相比其他地区,乌蒙山峨眉栲、华木荷群系类型多样,可划分为6个类型。乔木层常绿与落叶树种混杂,峨眉栲种群大树数量多,华木荷小苗数量巨大;灌木层常以竹类为优势种,反映出偏湿性的生境特征。(2)海拔、坡度和太阳年辐射值是影响群落分布格局的主导因素。(3)相比峨眉山同类群落,乌蒙山优势种种群中峨眉栲和水青冈种群呈下降型,峨眉山优势种种群都呈稳定型或增长型。
致谢 本研究野外调查得到乌蒙山国家级自然保护区管理局的大力支持和协助,谨以致谢!
参考文献:
DING T,DU F,WANG J,et al.,2006. Analysis on life form characteristics of the mid-montane humid evergreen broad-leaved forest in lancangjiang nature reserve [J]. J SW Coll,26(2):19-23.[丁涛,杜凡,王娟,等,2006. 澜沧江自然保护区中山湿性常绿阔叶林生活型特征研究 [J]. 西南林业大学学报,26(2):19-23.]
GONG HD,YANG GP,LU ZY,et al.,2011. Diversity and spatial distribution patterns of trees in an evergreen broad-leaved forest in the Ailao mountains,Yunnan [J]. Biodivers Sci,19(2):143-150. [巩合德,杨国平,鲁志云,等,2011.哀牢山常绿阔叶林树种多样性及空间分布格局[J]. 生物多样性,19(2):143-150.]
JIN ZZ,1979. The type and characteristic of evergreen broad-leaf forest in Yunnan [J]. Acta Bot Yunnan,(1):92-107. [金振洲,1979. 云南常绿阔叶林的类型和特点[J]. 云南植物研究,(1):92-107.]
KIKVIDZE Z,OHSAWA M,2002. Measuring the number of co-dominants in ecological communities [J]. Ecol Res,17(4): 519-525.
MCCUNE B,KEON D,2002. Equations for potential annual direct incident radiation and heat load [J]. J Veg Sci,13(4):603-606.
MENG GT,CHAI Y,YUAN CM,et al.,2013. Community characteristics of the mid-montane humid ever-green broad-leaved forest in gaoligong mountains,Yunnan [J]. Sci Silv Sin,49(3):144-151. [孟广涛,柴勇,袁春明,等,2013. 云南高黎贡山中山湿性常绿阔叶林的群落特征 [J]. 林业科学,49(3):144-151.]
PENG H,WU ZY,1998. The preliminary floristical study on mid-montane humid evengreen broad-leaved forest in Wuliang mountain[J]. Acta Bot Yunnan,20(1):12-22. [彭华,吴征镒,1998. 无量山中山湿性常绿阔叶林及其植物区系的初步研究[J]. 云南植物研究,20(1):12-22.]
ROBERT P,TARBOTON DG,GOODWIN CN,2005. Sinmaps uses manual [Z].
Sichuan Vegetation Cooperation Group,1980. The vegetation of Sichuan [M]. Chengdu:Sichuan Peoples Publishing Press:101-104. [四川植被协作组,1980. 四川植被 [M]. 成都:四川人民出版社:101-104.]
SONG YC,1988. The essential characteristics and main types of the broad-leaved evergreen forest in China [J]. Phytocoenologia,16(1):105-123.
SONG YC,2004. Tentative classification scheme of evergreen broad-leaved forests of China [J]. Chin J Plant Ecol,28(4):435-448. [宋永昌,2004. 中国常绿阔叶林分类试行方案[J]. 植物生态学报,28(4):435-448.]
SONG YC,2013. Evergreen broad-leaved forest in China [M]. Beijing:Science Press:61-77,181-189. [宋永昌,2013. 中国常绿阔叶林 [M]. 北京:科学出版社:61-77,181-189.]
TANG CQ,YANG Y,OHSAWA M,et al.,2011. Population structure of relict metasequoia glyptostroboides and its habitat fragmentation and degradation in south-central China [J]. Biol Conserv,144(1):279-289.
WANG JS,YAO SC,PU Q,et al.,2016. Quantitative classification and ordination of grassland communities on the northern Tibetan plateau[J]. Acta Ecol Sin,36(21):6889-6896. [王景升,姚帅臣,普穷,等,2016. 藏北高原草地群落的数量分类与排序[J]. 生态学报, 36(21):6889-6896.]
WANG RZ,GUO QX,2016.Woody plants species-area relationships in a broad-leaved korean pine forest in the Xiaoxing’an mountains [J]. Acta Ecol Sin,36(13):4091-4098. [王睿智,國庆喜,2016. 小兴安岭阔叶红松林木本植物种-面积关系[J]. 生态学报,36(13):4091-4098.]
WU ZY,ZHU YC,1987. The vegetation of Yunnan[M]. Beijing:Science Press:193-196. [吴征镒,朱彦丞,1987. 云南植被[M]. 北京:科学出版社:193-196.]
YANG YC,ZHUANG P,LI XR,1994. Ecological studies on the forest community of Castanopsis platyacantha-Schima sinensis on Emei Mountain [J]. Chin J Plant Ecol,18(2):105-120. [杨一川,庄平,黎系荣,1994. 峨眉山峨眉栲、华木荷群落研究 [J]. 植物生态学报,18(2):105-120.]
ZHANG JT,2004. Quantitative ecology [M]. Beijing:Science Press:189-193. [张金屯,2004. 数量生态学 [M]. 北京:科学出版社:189-193.]
