水分胁迫对柠条基质栽培黄瓜幼苗生长及光合特性的影响
2019-09-10曲继松张丽娟朱倩楠田永强高丽红
曲继松 张丽娟 朱倩楠 田永强 高丽红



摘要:研究水分胁迫对柠条基质栽培黄瓜幼苗光合作用及相关生理指标的影响,为利用新型园艺基质培育优质黄瓜幼苗提供理论参考。采用72穴标准穴盘进行育苗,测定正常供水(CK)、中度水分胁迫(5%PEG6000,DR-M)和重度水分胁迫(10% PEG-6000,DR-S)处理下黄瓜幼苗的株高、茎粗、根冠比、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、光合参数、叶绿素荧光参数的变化。结果表明,水分胁迫显著抑制了黄瓜幼苗的生长,处理后第17d,与CK相比,DR-M处理的株高降低8.30%,DR-S处理的株高降低16.20%,DR-M处理的黄瓜幼苗茎粗增加0.85%,DR-S处理的茎粗下降7.67%。与CK相比,DR-M处理下黄瓜幼苗干物质积累量增加,根冠比下降,叶绿素含量降低,丙二醛含量增加,SOD和POD活性升高,净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、水分利用效率(WUE)、初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、单位反应中心捕获的用于电子传递的能量(ETo/RC)均下降。对夏季柠条基质培育黄瓜幼苗而言,中度水分胁迫处理可抑制因温度过高、水分充足引起的徒长,有利于提高抗逆能力,增加干物质积累量,培育优质壮苗。
关键词:柠条基质;黄瓜;育苗;水分生理;光合作用
中图分类号:S642.2
文献标识码:A
文章编号:1000-4440(2019)02-0384-07
黄瓜(Cucumis sativus Linn)喜温暖,是设施蔬菜生产主栽作物种类之一。由于黄瓜易管理,生长速度快,茬口安排靈活,因此形成了周年栽培和供应的生产模式”。培育壮苗是园艺作物生产的关键环节,因为健壮的幼苗不仅对提高作物抗逆性十分有利,也是果实生产的基础和丰产的保证。穴盘育苗的出现革新了育苗方式,其基质成分和相对比例均显著影响幼苗的生长。以柠条粉作为育苗基质的试验已经取得了初步成效,尤其是在西瓜[2]、甜瓜(3]茄子[4]、辣椒[5]等的育苗上均有较好表现。水分管理是穴盘基质育苗的关键环节,水分高低影响着幼苗的质量好坏。有研究结果表明,土壤含水量为田间最大持水量的90%时,黄瓜幼苗茎粗最大,光合速率最高,叶绿素含量高,根系活力强,幼苗长势健壮[6]。适量的水分有利于保持土壤中水气比例的平衡,增强土壤中微生物的活性,促进根系的良好生长,提高吸收能力,增加干物质和叶绿素的积累量,从而促进光合能力的增强以及净光合速率的提高,为后期的栽培管理奠定基础。目前,有关水分胁迫对黄瓜影响的报道主要集中在模拟水分胁迫对黄瓜幼苗叶绿素、丙二醛(MDA)、脯氨酸、超氧化物歧化酶等生理指标的影响方面,有关水分胁迫对黄瓜幼苗光合特性和叶绿素荧光参数影响的研究较少。光合作用是作物最基本也是最重要的生理功能,对水分胁迫非常敏感,植物受到水分胁迫后会出现光合速率下降的现象171。气孔限制是水分胁迫下植物光合速率下降的主要原因[12],严重胁迫时非气孔因子起主导作用[13]。叶绿素荧光是探测和分析植物光合功能的重要指标,为研究光系统II(PS II)及其电子传递过程提供了丰富信息。牛铁泉等[14]以苹果为试验材料,发现水分胁迫下Fv/Fo和Fv/Fm显著降低,PS II反应中心光合潜能下降。叶片气体交换和叶绿素荧光是光合作用的探针,通过对叶片气体交换参数和叶绿素荧光参数进行分析,可以了解植物叶片对光能的吸收和利用[15-16]因此,研究水分胁迫下黄瓜叶片气体交换参数和叶绿素荧光参数的变化,可以进一步了解水分胁迫对黄瓜光合特性的影响。
本试验拟以德尔99黄瓜为试验材料,研究水分胁迫对柠条基质栽培黄瓜幼苗主要生理指标的影响,旨在了解黄瓜幼苗的抗旱机制,揭示水分胁迫与黄瓜幼苗光合作用及相关生理指标之间的关系,探索利于幼苗生长的柠条基质水分含量,以期为柠条基质培育黄瓜幼苗的水分管理提供理论参考和技术支撑。
1 材料与方法
1.1 试验地概况
试验在宁夏农林科学院园林场试验基地的育苗专用温室内进行,位于东经106°9'0.55",北纬38° 38'57.89",海拔1117m,属于典型的中温带大陆性气候,年平均气温8.5℃左右,年平均日照时数2800~3000 h,年平均降水量200 mm左右,无霜期185d左右。
1.2 试验材料
供试黄瓜品种为德尔99(DR),种子购自天津德瑞特公司,柠条粉购自宁夏回族自治区盐池县源丰草产业有限公司。1m3柠条粉中加入2 kg尿素和5 kg商品有机肥(N:P20,:K20=12:8:9,质量比),高温静态厌氧-好氧交替发酵90d后,加入珍珠岩和蛭石(柠条粉:珍珠岩:蛭石=7:2:1,体积比),作为育苗基质。使用29 cmx58 cm标准72穴穴盘。
柠条复配基质物理性状为:干体积质量0.2117g/cm3、湿体积质量0.5376g/cm3、总孔隙度60.25%、通气孔隙度27.66%、持水孔隙度32.59%、大小孔隙比0.8487。
柠条复配基质化学性质为:pH5.50、电导率9.46mS/cm、全盐21.50 g/kg、有机质402.00 g/kg、全氮18.06 g/kg、速效氮1128.00 mg/kg、速效磷127.50mg/kg速效钾2450.00 mg/kg、碳氮比12.90。
1.3 试验设计
试验于2017年5月20日-2017年8月30日在9m跨度育苗专用日光温室内进行,育苗床架上的每个穴盘按照南北方向摆放,所有穴盘横向并一字排开,与温室前沿平行,距离温室前沿3m。
试验设3个处理,分别为:对照(CK,用清水浇灌)、中度水分胁迫处理(DR-M,用体积分数为5%的PEG-6000溶液浇灌)、重度水分胁迫处理(DR-S,用体积分数为10%的PEG-6000溶液浇灌)。每个处理3穴盘,重复3次[17]。当幼苗长到1叶1心(6月27日)时开始处理,之后每7d(7月4日、7月11日)处理1次,每次处理后的第3d进行生长指标和生理指标的测量。第1次处理后的第10d测定气体交换参数和叶绿素荧光参数。
1.4 测定项目及方法
1.4.1 生长指标用直尺测量幼苗株高,游标卡尺测量幼苗茎粗,采用氯化三苯基四氮唑(TTC)法测定根系活力,根冠比=地下部干质量/地上部干质量,每个处理随机选择3株幼苗进行测量。
1.4.2 生理指标乙醇比色法测定叶片叶绿素(Chl.)含量,硫代巴比妥酸(TBA)显色法测定MDA含量,氯化硝基四氮唑蓝(NBT)光化还原法测定超氧化物歧化酶(SOD)活性,愈创木酚比色法测定过氧化物酶(POD)活性[18]。以顶部最大的2片完全展开功能叶片为材料,每个样本测量3次,结果取平均值。
1.4.3 光合参数选取幼苗2叶2心时的最大功能叶片为试验材料,每个处理随机选择3片。采用TPS-2便携式光合作用测定系统对净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)胞间CO,浓度(Ci)、气孔限制值(Ls)、水分利用效率(WUE)等光合参数进行测定,育苗温室内部光照度为(1000+50)μmol/(m2·s),CO2浓度为(400+20)μmol/mol。
1.4.4 叶绿素荧光参数选取幼苗2叶2心时的最大功能叶片为材料,每个处理随机选择3片。使用Hansatech公司生产的连续激发式荧光仪Handy PEA测定黄瓜幼苗叶片的荧光参数,测定前暗适应30min,利用配套软件对数据进行处理,测试指标有:初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、PSII最大光化学效率(Fv/Fm)、性能指数(PI)、单位反应中心吸收的光能(ABS/RC)、单位反应中心捕获的用于还原结合有质体醌的蛋白质(QA)的能量(TRo/RC)、单位反应中心捕获的用于电子传递的能量(ETo/RC)、单位反应中心耗散掉的能量(DIo/RC)[19]测定时间为10:00-12:00。
1.5 数据分析
数据处理和制图采用Excel 2007软件和DPS 3.01软件Duncan's新复极差法进行。
2 结果与分析
2.1 水分胁迫对黄瓜幼苗生长发育的影响
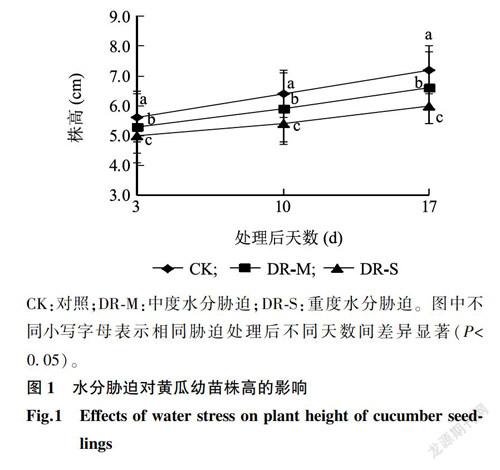
2.1.1 生长状况图1显示,CK处理的黄瓜幼苗生长较快。水分胁迫对黄瓜幼苗株高的影响较大,PEG-6000处理后第17d,DR-M处理和DR-S处理的黄瓜幼苗株高均显著低于CK,分别降低了8.30%、16.20%。说明水分胁迫抑制了幼苗的正常生长,而且胁迫程度越大抑制效果越明显。
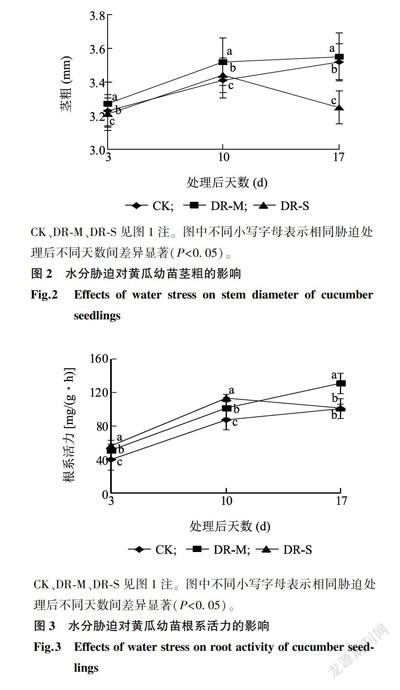
图2显示,PEG-6000处理后第17d,DR-M处理的黄瓜幼苗茎粗比CK高0.85%,但DR-S处理的黄瓜幼苗茎粗却显著低于CK,下降了7.67%,表明适当的水分胁迫处理可以使幼苗茎粗增大。
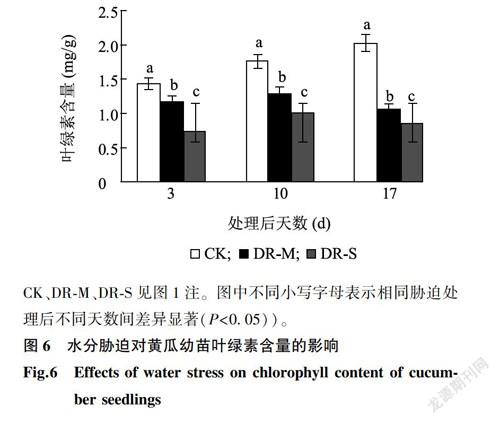
2.1.2 根系状况图3显示,处理后第10d,与CK相比,DR-M处理下黄瓜幼苗的根系活力提高了16.22%,DR-S处理下黄瓜幼苗的根系活力提高了29.29%。表明短期水分胁迫有利于幼苗根系活力的提升,而且胁迫强度越高根系活力提高越多。
随着水分胁迫时间的延长,DR-M处理根系活力持续增强,处理后第17d,较CK提升29.69%,较7d前增加28.68%。DR-S处理的水分胁迫程度最高,随着水分胁迫时间的延长,黄瓜幼苗出现萎蔫状况,根系活力出现了下降趋势,处理后第17d,较CK提高0.78%,较7d前下降10.10%。说明中度水分胁迫可以增强幼苗的根系活力,但重度且长时间的水分胁迫会伤害幼苗的根系,导致根系活力降低。
2.1.3 干物质积累量图4显示,处理后第3d,与CK相比,DR-M处理下的干物质积累量增加9.75%,DR-S处理下的干物质积累量减少1.43%。处理后第10d,各处理干物质积累量之间的差异加大,处理后第17d,DR-M处理的干物质积累量较CK增加21.43%,而DR-S处理较CK下降19.55%,DR-M处理较DR-S处理增加50.94%,差异均达到显著水平。表明中度水分脅迫有利于黄瓜幼苗干物质的积累。
图5显示,CK的黄瓜幼苗根冠比较高,经过PEG-6000处理的幼苗根冠比较低,随着PEG-6000体积分数增大,根冠比降低。随着水分胁迫时间的延长,CK根冠比逐渐升高,而DR-M和DR-S处理的根冠比在处理后第10d小幅上升,处理后第17d又下降。中度水分胁迫可以降低黄瓜幼苗的根冠比,防止幼苗徒长。高度水分胁迫同样降低黄瓜幼苗的根冠比,而且根冠比值更低。
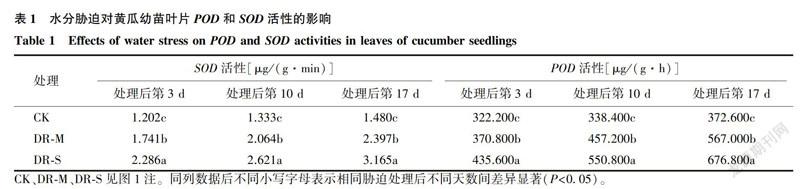
2.1.4 叶绿素含量图6显示,CK的黄瓜叶片叶绿素含量较高,而且随着时间的延长,叶绿素含量逐渐升高。水分胁迫下,幼苗叶片叶绿素含量均较CK降低。PEG-6000处理第3d,CK组的叶绿素含量为1.427 mg/g,DR-M处理组的叶绿素含量为1.168 mg/g,DR-S处理组的叶绿素含量为0.727 mg/g。DR-M处理后第10d的叶绿素含量较处理后第3d增加10.19%,DR-S处理后第10d较处理后第3d增加38.93%,DR-M处理后第17d较处理后第10d降低18.03%,DR-S处理后第17d较处理后第10d降低15.64%。处理后第17d,DR-M和DR-S处理组黄瓜幼苗叶片的叶绿素含量与CK相比,分别降低47.80%和57.84%(P<0.05)。说明黄瓜幼苗受到水分胁迫后,随胁迫时间延长叶绿素含量下降,幼苗叶片变黄并趋于死亡。
2.1.5 MDA含量图7显示,CK的黄瓜幼苗叶片的MDA含量较低,随着胁迫时间延长,MDA含量缓慢升高。PEG-6000处理使黄瓜幼苗叶片MDA含量显著增加,处理后第10d,与CK相比,DR-M处理和DR-S处理的黄瓜幼苗叶片MDA含量分别增加16.30%、73.64%(P<0.05)。CK、DR-M和DR-S3个处理的线性相关方程直线斜率分别为0.105、0.248、0.341,斜率的大小反映了幼苗叶片MDA含量增加速率的大小。由此可知,黄瓜幼苗只能忍受短时间的中度水分胁迫,随着水分胁迫时间延长和胁迫程度的增加,黄瓜幼苗的生长发育严重受阻,甚至出现萎蔫。这是因为,黄瓜植株在逆境下保护酶活性下降,细胞内自由基平衡被打破,自由基累积量增加,造成膜脂过氧化,产生较多的MDA,使膜结构受损程度加重,导致植物受害甚至死亡![20]
2.1.6 SOD和POD活性表1显示,随着胁迫时间的延长,DR-M和DR-S处理黄瓜幼苗叶片的SOD和POD活性逐渐升高。处理后第17d,与CK相比,DR-M处理组黄瓜幼苗叶片的SOD和POD活性分别提高了61.96%和52.17%,DR-S处理分别提高113.85%和81.64%。水分胁迫可以提高SOD和POD的活性,降低细胞膜系统的伤害,抗细胞衰老,增强幼苗的木质化程度。
2.2 水分胁迫对黄瓜幼苗叶片光合参数的影响
表2显示,随着水分胁迫程度的加剧,黄瓜幼苗的Pn、Tr、Gs、Ci均显著降低,CK的黄瓜幼苗叶片WUE最高,为2.4063,DR-M处理次之,为2.2541,DR-S处理最小,为1.6509。水分胁迫下,Pn下降,主要是由于植物叶片气孔阻力增大甚至,直接关闭气孔,Gs下降,导致叶肉细胞梭化位点的CO2量减少,Ci减少,而且胁迫程度越大,Pn越低。
2.3水分胁迫对黄瓜幼苗叶绿素荧光参数的影响
表3显示,水分胁迫下黄瓜幼苗叶片叶绿素荧光基本参数Fo、Fm和Fv变化较大,且各处理间的差异达到显著水平。CK的黄瓜幼苗Fo、Fm和Fv最大,分别为544.50、2511.50和1995.00。水分胁迫严重影响黄瓜幼苗的性能指数,CK最高,为1.0885,比DR-S处理高118.31%。DR-M和DR-S处理叶片的Fv/Fm降低,说明水分胁迫下,植物发生了光抑制。
2.4 水分胁迫对黄瓜幼苗PSII反应中心活性参数的影响
表4显示,3个处理的黄瓜幼苗均表现为ABS/RC>TRo/RC>ETo/RC,表明随着电子传递链的延伸,热耗散增加,光能利用率降低。DR-M和DR-S处理的ETo/RC均比DIo/RC小,CK则相反,说明水分胁迫使得PSII反应中心用于热耗散的能量高于用于电子传递的能量。随着水分胁迫程度的加重,单位反应中心吸收的能量增加,但在碳同化降低的情况下,易导致PSII的过量激发,造成PSII的进一步损害。
3 讨论
水分胁迫程度越大,对株高的抑制越显著。中度水分胁迫有利于增加黄瓜幼苗干物质的积累量,降低黄瓜幼苗的根冠比,增加幼苗茎粗,增强根系活力,提高根系吸收能力,但重度水分胁迫以及长时间的水分胁迫会伤害幼苗的根系,导致根系活力显著降低。说明中度水分胁迫有助于提高抗逆能力,益于柠条基质中黄瓜壮苗的培育。
植物受到水分胁迫时,通过渗透调节降低渗透势,维持细胞膨压是保证体内正常生理功能的重要调节方式。SOD和POD是细胞膜活性氧清除酶系统中的关键酶,在清除超氧自由基,控制膜脂过氧化以及保护细胞膜正常代谢等方面起着重要的作用[13]。本研究结果表明,随着PEG-6000体积分数的增加,黄瓜幼苗叶片中SOD、POD酶活性升高。
中度水分脅迫时,气孔导度的限制是导致光合速率降低的主要原因,重度水分胁迫时,叶肉细胞利用CO2的能力降低,光合能力下降是引起光合速率降低的主要原因[21]水分胁迫会导致叶片气孔导度下降,CO2反应受阻,进而造成叶片光合能力降低,作物可以通过提高叶片水分利用效率来适应这种逆境[22-23]。气孔导度是反映叶片气体交换的重要指标,植物气孔的开闭主要受光照和CO2控制,表现为昼开夜闭,但当植物缺水时,水分就成为决定气孔开闭的决定性因素,水分过少会导致植物叶片气孔关闭,使植物避免因继续大量失水而受到伤害,进而导致光合速率下降。作物对水分胁迫最早、最敏感的响应是气孔的关闭[24],本研究结果中气孔导度大小与水分胁迫程度呈负相关,即随着水分胁迫程度加剧,气孔导度值逐渐降低。水分胁迫明显降低了植株的气孔导度、蒸腾速率净光合速率。
作物受到水分胁迫后会出现光合速率下降的现,象[25-27]黄瓜幼苗叶绿素荧光参数中的Fo、Fm和Fv受水分胁迫的影响较大。作物水分利用效率是其生存的关键因子[28]和抗旱策略的重要组成部分[29]。
参考文献:
[1]张曼义,杨再强,侯梦媛.水分胁迫下黄瓜叶片光响应过程的模拟[J].中国农业气象,2017,38(10):644-654.
[2]曲继松,郭文忠,张丽娟,等.柠条粉作基质对西瓜幼苗生长发育及干物质积累的影响[J].农业工程学报,2010,26(8):291-295.
[3]张丽娟,曲继松,冯海萍,等.利用柠条粉发酵料作为育苗基质对甜瓜幼苗质量的影响[J].北方园艺,2010(15):165-167.
[4]曲继松,张丽娟,冯海萍,等.混配柠条复合基质对茄子幼苗生长发育的影响[J].西北农业学报,2012,21(11):162-167.
[5]曲继松,张丽娟,冯海萍,等.发酵柠条粉混配基质对辣椒幼苗生长发育的影响[J].江苏农业学报,2012,28(4):846-850.
[6]董绪兵,毕焕改,刘业霞,等.黄瓜幼苗干旱-低温交叉适应与渗透调节的关系[J].中国农业科学,2011,44(2):335-340.
[7]李晨,李秀杰,韩真,等.非生物胁迫对葡萄光合作用的影响研究进展[J].山东农业科学,2017,49(12):144-148.
[8]翁亚伟,张磊,张姗,等.盐旱复合胁迫对小麦幼苗生长和水分吸收的影响[J].生态学报,2017,37(7):2244-2252.
[9]马梦茹,王占林,贺康宁,等.不同土壤含水量与光照对山杏和四翅滨藜光合作用的影响[J].江苏农业科学,2017,45(22):126-129.
[10]JEYARAMRAJA P R,MEENAKSHIS N,JOSHISD,etal. Water deficit induced oxidative damage in tea(Camellia sinensis)plants[J].Journal of Plant Physiology,2005,162:413-419.
[11]李佳,刘立云,李艳,等.保水剂对干旱胁迫下槟榔幼苗生.理特征的影响[J].南方农业学报,2018,49(1):104-108.
[12]张志焕,韩敏,张逸,等.水分胁迫对不同抗旱性砧木嫁接番茄生长发育及水气交换参数的影响[J].中国农业科学,2017,50(2):391-398.
[13]桑子阳,马履一,陈发菊.干旱胁迫对红花玉兰幼苗生长和生理特性的影响[J].西北植物学报,2011,31(1):109-115.
[14]牛铁泉,田给林,薛仿正,等.半根及半根交替水分胁迫对苹果幼苗光合作用的影响[J].中国农业科学,2007,40(7):1463-1468.
[15]睢晓蕾,毛胜利,王立浩,等.弱光条件下辣椒幼苗叶片的气体交换和叶绿素荧光特性[J].园艺学报,2007,34(31):615-622.
[16]MAXWELL K,JOHNSON G N.Chlorophyll fluorescence —— apractical guide[J].Journal of Experimental Botany,2000,51(345):659-668.
[17]吴顺,张雪芹,蔡燕.旱胁迫对黄瓜幼苗叶绿素含量和光合特性的影响[J].中国农学通报,2014,30(1):133-137.
[18]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[19]李鹏民,高辉远,RETOJ S.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J].植物生理学与分子生物学学报,2005,31(6):559-566.
[20]肖姣娣.不同强度干旱胁迫对刺槐幼苗生理生化特性的影响[J].中南林业科技大学学报,2015,35(8):23-26.
[21]ZHOU S,LI M,GUAN Q,et al.Physiological and proteome analysis suggest critical roles for the photosynthetic system for high water-use eficiency under drought stress in Malus[J].Plant Science,2015,236:44-60.
[22]高玉紅,牛俊义,徐锐,等.不同覆膜方式对玉米叶片光合、蒸腾及水分利用效率的影响[J].草业学报,2012,21(5):178-184.
[23]于文颖,纪瑞鹏,冯锐,等.不同生育期玉米叶片光合特性及水分利用效率对水分胁迫的响应[J].生态学报,2015,35(9):2902-2909.
[24]孟凡超,张佳华,郝翠,等.CO2浓度升高和不同灌溉量对东北玉米光合特性及产量的影响[J].生态学报,2015,35(7):2126-2135.
[25]GALLE A,HALDIMANN P,FELLER U.Photosynthetic performance and water relations in young pubescent oak(Quercus pubescens)trees during drought stress and recovery[J].New Phytologist,2007,174(4):799-810.
[26]FLEXAS J,BOTA J,GALMES J,et al.Keeping a positive carbon balance under adverse conditions:responses of photosynthesis andrespiration to water stress[J].Physiologia Plantarum,2006,127(3):343-352.
[27]孙东宝,王庆锁.水分对苜蓿叶片光合特性的影响[J].植物生态学报,2012,36(1):72-80.
[28]ZHANG G C,XIAJ B,SHAO H B,et al.Grading woodland soil water productivity and soil bioavailability in the semiarid loess plateau of China[J].Clean-Soil Air Water,2012,40(2):148-153.
[29]SAUD S,YAJUN C,FAHAD S,et al.Silicate application increases the photosynthesis and its associated metabolic activities inKentucky bluegrass under drought stress and post-drought recovery[J].Environmental Science and Pollution Research,2016,23(17):17647-17655.
