三沙湾夏季大型底栖动物群落结构及其和水产养殖活动关系
2019-09-03纪炜炜
王 楠,纪炜炜,付 婧,周 进
(1.中国水产科学研究院东海水产研究所,上海 200090;2.上海海洋大学水产与生命学院,上海 201306)
大型底栖动物是海洋生态系统中最为重要的生物类群之一,其具有较高的物种多样性、较强的次级生产能力和较为理想的环境指示功能。在中国近岸水域,针对此类群已有较多研究,内容涵盖种群、群落和生态系统等多个层次。其中,群落研究仍是目前底栖生物研究中的重要内容之一,本研究的区域三沙湾也是如此,针对此水域内的底栖群落结构已有多篇研究[1-9]。因底栖生态学研究中常用的抽样调查方法特点所限,基于有限样本的群落结构描述和实际群落组成之间存在一定差距,因此针对三沙湾水域底栖群落结构的进一步研究仍有较为重要的学术意义。
本研究区域三沙湾是我国东海区最为典型的水产养殖海湾之一,主要网箱养殖大黄鱼(Pseudosciaena crocea),是我国养殖产量最大的单品种海水鱼类。近年来,三沙湾大黄鱼养殖产量约占全国的70%[10]。网箱养殖引发养殖衍生有机物(aquaculture-derived organic matter,AOM)沉降,其在沉积物环境中的过度积累可通过营养关系影响底栖生物群落,该假设在国内外均已被证实[11-15]。同时,区域内存在的贝类养殖也已被证明可能影响底栖群落或环境[16-17]。然而,养殖活动对于底栖生物群落的影响受养殖强度和区域水文动力、沉积物特性等自然地理状况等多方面因素共同影响,故已有的针对不同海湾的研究结果之间存在差异。目前对于三沙湾内水产养殖活动对底栖生态环境效应的认识尚不全面,已有相关研究多通过对比分析不同类型水域(养殖和非养殖以及不同养殖类型)内群落时间变化趋势的差异[6,8],部分关于群落空间差异的分析结论通常基于较为有限的采样站位数据[4,8],故基于较大空间范围和较多采样站位数据的空间差异研究亟待补充。
本研究根据2016年夏季在三沙湾水域内大面积采样的生物群落和环境因子数据,描述区域内大型底栖动物群落结构特征,探究群落和典型环境因子的相关性,揭示典型群落参数在鱼类网箱、海带吊绳、鲍鱼筏式养殖水域以及自然水域共4种不同类型区域间的差异性,以期在描述区域群落结构现状的基础上,揭示水产养殖活动对于底栖生物群落的影响。
1 材料与方法
1.1 三沙湾养殖状况
三沙湾水产养殖活动始于20世纪80年代,伴随大黄鱼人工养殖的成功,90年代开始海湾内网箱养殖规模迅速增加。2017年三沙湾大黄鱼网箱养殖产量已达1.47×105t[10]。同时,该水域内海带(Laminaria japonica)、龙须菜(Gracilaria lemaneiformis)和鲍鱼(Haliotis discus hannai)的养殖产量也维持在较高水平,2014年三者湿重产量分别为1.3×105t、1.9×104t和 1.6×105t。大黄鱼养殖方式主要为标准网箱养殖(尺寸:3 m×3 m×4 m),网箱遍布海湾各区。网箱养殖周期为1—2年,幼鱼通常在4—5月和10—12月期间繁育,培养至体长10 cm后按照1500尾/网箱密度进行养殖,约8—13个月后长至商品鱼尺寸后出售。养殖饵料主要来源于我国近海捕获的小型杂鱼,通常包括鳀(Engraulis japonicus)、沙丁鱼(Sardine pilchardus)、龙头鱼(Harpadon nehereus)和带鱼(Trichiurus lepturus)等。海带和龙须菜采用吊绳方式养殖,遍布海湾中部及内湾各区,12月至次年5月为海带养殖期,6—9月为龙须菜养殖期。鲍鱼使用筏式养殖,仅限于湾口区小范围水域,主要饵料为湾内养殖的海带和龙须菜。
1.2 样品采集时间和站位
本研究于2016年8月在三沙湾采集大型底栖动物样品及同步环境数据,共设置4种类型共41个采样站位(图1)。距离养殖设施5 m范围以内区域定义为养殖水域,其内共包括23个大黄鱼网箱养殖站位(6、7、9~12、16~24、28~30、32、36、38~40号)、9个海藻吊绳养殖站位(2、3、5、13、27、31、33、37、41号)和 2个鲍鱼筏式养殖站位(34、35号)。距离养殖设施600 m以外区域为自然水域[18],水域内共包括7个采样站位(1、4、8、14、15、25、26号)。
1.3 底栖生物采集及处理方法
使用面积为0.04 m2箱式采泥器采集底泥样品,每站成功取样2次合并为1个样品。使用最小孔径为0.50 mm筛网冲洗样品,所获大型底栖生物样品使用75%酒精现场固定并保存。固定样品在实验室内使用1%虎红(rose bengal)溶液染色,静置24 h后进行粗分。粗分后样品进行分类学鉴定、个体计数及称重(湿重),计算丰度(个·m-2)和生物量(g·m-2)。具体操作方法参考《海洋调查规范》(GB/T 12763.6-2007)相关要求进行。
1.4 环境因子数据采集及检测
温度、盐度、pH和溶解氧等水文环境因子使用多功能水质参数仪(YSIEX02)现场获取。沉积物环境因子重点选择与水产养殖活动关系较为密切的要素,包括 δ15N、δ13C、C/N比、总氮、总有机碳、总磷、含水率、硫化物和粉砂-黏粒含量。环境因子和生物样品同步采集,环境因子检测方法参照唐盟[19]相关叙述。
1.5 参数计算
分析三沙湾底栖动物群落结构与环境因子关系时,本文选取最为典型的6种群落参数:物种数(S)、丰度(N)、生物量(B)、Shannon-Wiener多样性指数 (H′)、Pielou均匀度指数 (J′)、Margalef丰富度指数(d)。
1.5.1 Shannon-Wiener多样性指数(H′)
各采样站位 Shannon-Wiener多样性指数(H′)公式为:
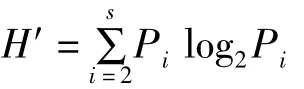
式中,Pi为第i种的个数与该样方总个数之比值,S为样方种数。

图1 三沙湾2016年8月航次大型底栖动物和环境因子采样站位图Fig.1 M ap of the study area
1.5.2 Margalef丰富度指数(d)和Pielou均匀度指数(J′)
各采样站位 Margalef丰富度指数(d)和Pielou均匀度指数(J′)公式分别为:
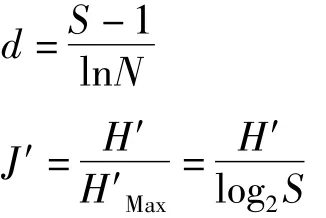
式中,S为种类数,N为总丰度,H′为 Shannon-Wiener多样性指数。
1.6 数据处理
本文选取7种最为典型的底栖群落参数进行研究,包括物种数(S)、丰度(N)和生物量(B)、多样性指数 (H′)、Pielou均 匀度 指数 (J′)、Margalef丰富度指数(d)和单纯度指数(C),各参数通过Primer 6.0软件计算获取。分析群落参数在不同类型水域之间的差异时,首先通过Shapiro-Wilk方法检验各参数是否符合正态分布,再通过Bartlett检验进行数据方差齐性检验。结果表明本研究计算出的7种群落参数大多数数据组不能同时满足正态分布和方差齐性的要求。此外,由于鲍鱼养殖区样本有限,仅包括2个采样站位,故本研究使用非参数Kruskal-Wallis方法检验群落参数的组间差异,显著性水平设为P=0.05。同时,利用Behrens-Fisher方法进行两两比较,两两比较的显著性检验进行Bonferroni校正。此部分计算通过R软件完成。
为分析三沙湾夏季底栖动物的群落相似性,文章根据丰度数据对41个采样站位的底栖群落进行聚类分析(cluster analysis)和非度量多维尺度排序分析(nMDS),此分析利用Primer 6.0软件完成。因Bray-Curtis similarity模型计算样方之间的半几何距离可避免样品间相似性受到样品中均不存在物种的影响,比较符合本研究数据的特性,故本研究使用此种模型方法构建样方相似性矩阵。同时对于原始丰度数据进行平方根转化,以对少数优势种(大个体种)或多数稀有种(小个体种)进行平衡权重。聚类计算时,采用层次分析法中较为温和的average-linkage算法,以避免出现基于过度相似性或非相似性的聚类结果。
研究三沙湾底栖动物群落结构和环境因子关系时,首先对三沙湾水域群落数据进行除趋势对应分析 (deternded correspondence analysis,DCA),以确定群落数据的分布类型(单峰型或线性分布)。在本研究中,DCA分析结果显示4个轴重最大梯度超过4,因此选择基于单峰模型的典范对应分析方法(canonical correspondence analysis,CCA)。上述分析通过Canoco 4.5软件完成。
2 结果与分析
2.1 群落结构现状
2016年8月三沙湾海域采样共鉴定大型底栖动物6门75种(表1,附表1)。其中,环节动物物种数占据绝对优势,共计48种,占总物种数64.00%;其他门类物种数按从高到低顺序分别为节肢动物、软体动物、棘皮动物、纽形动物和腔肠动物。
三沙湾大型底栖动物的丰度和生物量均值分别为371.34个·m-2和21.82 g·m-2(表1)。环节动物对于底栖生物丰度和生物量的贡献量均为最大,类群丰度和生物量均值分别为292.68个·m-2(占底栖生物总丰度78.82%)和13.06 g·m-2(59.87%)。丰度和生物量在站位间存在较大差异,丰度最大值出现在网箱养殖站位第17号站,为1 075.00个·m-2;最小值出现在海带养殖站位第13号站,为25.00个·m-2。生物量最大值出现在网箱养殖站位第9号站,为128.23 g·m-2,最小值出现在海带养殖站位第41号站,为 0.15 g·m-2。
三沙湾大型底栖动物群落多样性(H′)均值为2.56(变化范围0.00~3.51),单纯度(C)均值为0.26(0.10~1.00),均匀度(J′)均值为 0.83(0.00~0.97),丰富度(d)均值为0.97(0.16~1.75)。
2.2 群落优势物种的空间分布
2016年8月三沙湾底栖群落中平均丰度位居前6位的物种皆为环节动物(表2),按数量从高到低顺序分别为丝异须虫(Heteromastus filiformis)、不倒翁虫(Sternaspis scutata)、欧努菲虫属一种(Onuphis sp.)、索沙蚕科未定种(Lumbrineridae)、角海蛹(Ophelina acuminata)和丝鳃虫属一种(Cirratulus sp.)。此外,软体动物胡桃蛤属一种(Nucula sp.)和焦河篮蛤(Potamocorbula ustulata)以及棘皮动物倍棘蛇尾属一种(Amphioplus sp.)在群落中也较具数量优势。数量优势物种通常在鲍鱼筏式养殖和网箱养殖水域中形成较高种群数量。

表1 2016年夏季三沙湾大型底栖动物群落组成Tab.1 Species com position ofmacrobenthic community in Sansha Bay in summer,2016
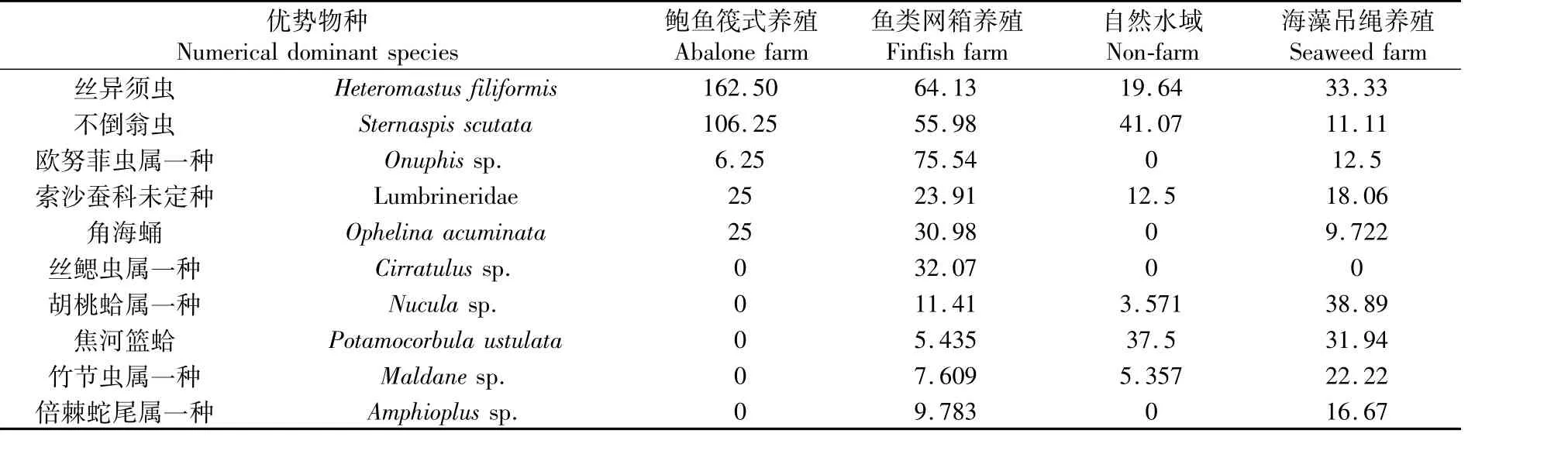
表2 2016年夏季三沙湾数量优势大型底栖动物栖息密度的空间分布(丰度值位居前10位物种)Tab.2 Spatial distribution pattern of abundance of top ten numerical dom inant macrobenthos in Sansha Bay in summer,2016 (个·m-2)
2.3 典型群落参数在不同类型养殖水域之间的差异
Kruskal-Wallis检验结果表明,本文研究的7个群落参数在鱼类网箱养殖水域、海藻吊绳养殖水域、鲍鱼筏式养殖水域和无养殖水域之间均无显著差异(P>0.05,图2)。后续两两比较显示显著性差异仅出现1次,即群落多样性指数(H′)在鱼类网箱养殖水域和无养殖水域之间存在显著差异。
2.4 三沙湾底栖动物群落结构的聚类分析
基于丰度数据的聚类结果显示,三沙湾大型底栖动物群落结构的空间异质性较高。按照30%相似性水平,区域内底栖生物可划分成11个群落(图3)。在同等类型的水体中,不同采样站位内的群落结构也呈现出较大差异,例如无养殖水域、海藻吊绳养殖等水域的站位均体现出此种空间差异。

图2 2016年夏季三沙湾大型底栖动物群落参数在不同类型水域间的差异Fig.2 Differences of indices ofmacrobenthic community between different waters in Sansha Bay in summer,2016
nMDS分析结果显示压力系数为0.19。在二维排序空间中,采样站位之间的位置较为分散(图4)。在各种类型的采样站位中,无养殖水域和海藻吊绳养殖水域站位之间空间差异性较强。
2.5 群落结构和环境因子的相关性分析
物种丰度值的除趋势对应分析(DCA)结果显示最长轴长度为6.330,故采用典范对应分析方法(CCA)进行排序,结果显示前两轴特征值分别为0.411和0.307,种类和环境因子排序轴的相关系数高达0.974和0.913,物种和环境关系累计百分比显示前两轴环境因子对物种分布特征的解释量可达71.8%,排序轴较好地反映底栖生物群落与环境因子之间的关系。
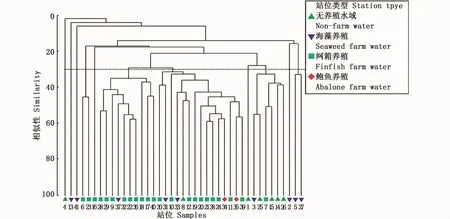
图3 2016年夏季三沙湾大型底栖动物群落的聚类分析Fig.3 Cluster analysis ofmacrobenthic community from different samp ling stations in Sansha Bay based on abundance data in summer,2016
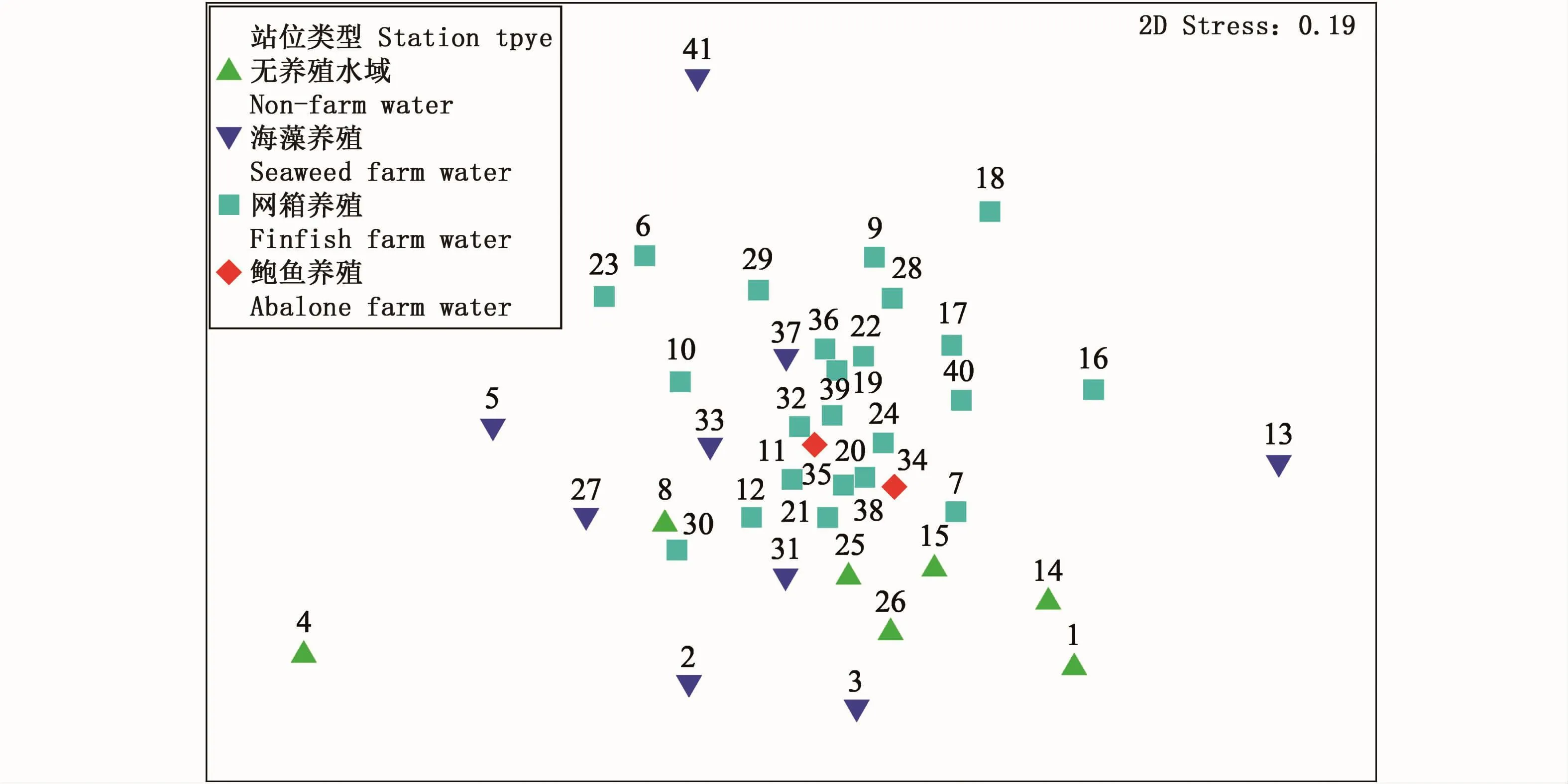
图4 2016年夏季三沙湾大型底栖动物群落nMDS分析Fig.4 The nMDS analysis ofmacrobenthic community in Sansha Bay in summer,2016
水体温度、盐度、水深和沉积物含水率、粉砂-黏粒含量、总氮和总有机碳等环境变量轴长较长,显示三沙湾内此类环境因子对于群落的影响较大(图5)。沉积物含水率、粉砂-黏粒含量、总氮和总有机碳等环境变量之间夹角较小,显示此类因子之间具有较强的正相关关系。大部分数量优势物种和水深以及沉积物总氮、总磷、总有机碳和含水率呈现较为显著的正相关关系。数量绝对优势物种(如1、2、4、5号物种等)多位于排序轴的中央位置。
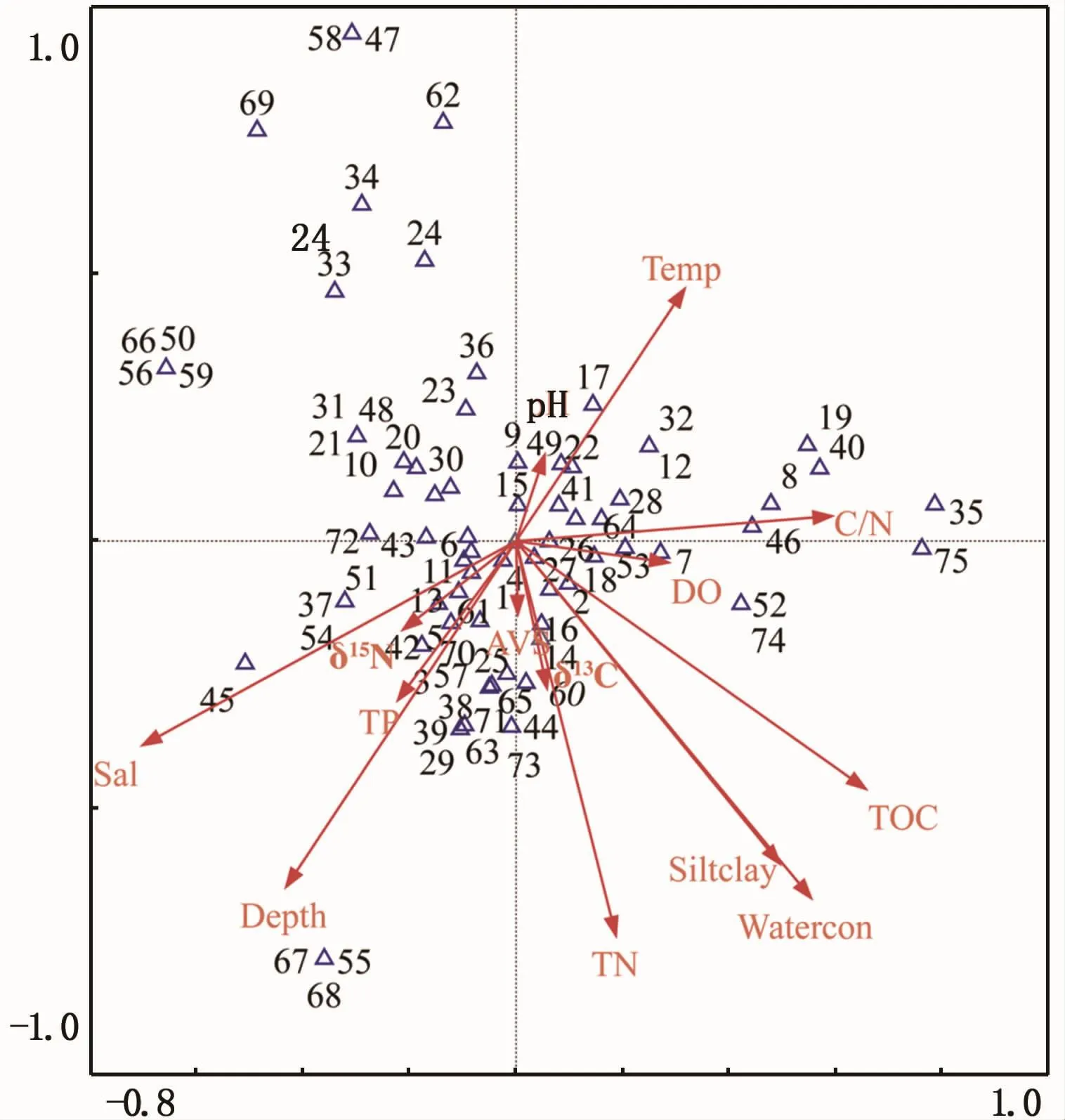
图5 2016年夏季三沙湾大型底栖动物与环境因子典型对应分析排序图Fig.5 Ordination diagrams of species(abundance data)and typical environmental variables in Sansha Bay based on canonical correspondence analysis in summer,2016
3 讨论
3.1 三沙湾大型底栖动物群落结构特征
根据2016年8月的采集样品,三沙湾共出现大型底栖动物75种,涵盖环节动物、节肢动物、软体动物和棘皮动物等浅海水域习见大型底栖动物类群。其中,环节动物物种数占群落物种总数达60%,类群数量优势明显;节肢动物和软体动物次之,其他门类较少。此种群落组成特征与中国近海其他水域内的底栖群落相似,在温带和亚热带软泥底质的生境中,环节动物物种数量在底栖样品中较具优势[20]。三沙湾大型底栖动物的丰度和生物量均值分别为371.34个·m-2和21.82 g·m-2,此种数量水平和东海近海水域内的其他相关报道数据相似[21],符合暖温带浅水水域底栖生物多样性的典型特征。
尽管三沙湾底栖生物在物种数、丰度和生物量等数量特征方面体现出与邻近海域内群落的相似性,但区域内群落具有其独特特征,例如三沙湾的底栖群落物种组成特征。以同属东海区、且同为养殖海湾的象山港和乐清湾为例,象山港底栖群落优势物种为异足索沙蚕(Lumbrineris heteropoda)、长吻沙蚕(Glycera chirori)、多鳃齿吻沙蚕(Nephtys polybranchia)和不倒翁虫等[22],乐清湾群落优势种主要为西格织纹螺(Nassarius siquinjorensis)、白沙箸(Virgularia gustaviana)、不倒翁虫、小头虫(Capitella capitata)、棘刺锚参(Protankyra bidentata)等[15],三沙湾内最具数量优势的物种丝异须虫在上述海湾不具数量优势,另一数量优势物种角海蛹在上述海湾则未有分布。此种相邻区域内底栖群落组成呈现显著差异的现象说明大型底栖动物是海洋生态系统中物种多样性水平较高的生态类群之一,且底栖群落形成机制较为复杂。同时,本文的聚类分析结论显示,三沙湾大型底栖动物群落结构空间异质性较高。即使按较低的30%相似性标准,水域内底栖生物仍可被划分为十余个群落。聚类分析重点旨在组群划分而非连续尺度上样品间关系的呈现,此方法比较适用于环境条件空间异质性较高、样品可被明确划分成组的情况。相比之下,排序方法可更好地表达生物群落对于比较连续的环境梯度的响应特点。nMDS标序是近年来底栖生态学中较为常用的方法之一,即按照样品间的非相似性等级顺序将样品排放在二维标序图中,同时引入压力系数以反映相似性等级与标序图中相应的距离等级的不一致程度。本文nMDS分析中压力系数为0.19,说明相关排序结果基本可信。本文的nMDS分析和聚类分析结论相似,均显示采样站位之间的相似性较低。
三沙湾内如此高的群落结构空间异质性较为特殊,可能源于其存在的多重外源胁迫。首先,区域内存在鱼类网箱、鲍鱼筏式和海藻吊绳等多种水产养殖方式,不同生产方式对于底栖生境的影响可能存在显著差异。同时,在水产养殖活动影响之外,海湾内频繁的船只航行、海洋工程建设等人类活动也可能对底栖生境产生扰动,此类胁迫容易造成底栖生境的不稳定性,从而影响区域内的底栖群落。例如无养殖水域4号站位丰度仅为75个·m-2,此站位较浅的水体(1.5 m)导致区域内底栖群落易受船只航行的影响。此站位中的寡鳃齿吻沙蚕(Nephtys oligobranchia)和双鳃内卷齿蚕(Aglaophamus dibranchis)在三沙湾其他站位中出现频率极低,显示该站位独特的物种组成。寡鳃齿吻沙蚕和双鳃内卷齿蚕同属多毛纲齿吻沙蚕科,此科物种对于物理扰动具有较强的适应能力,例如在长江口洋山深水港码头建设工程水域多毛类物种多样性极低,但此科物种仍可生存[23]。
3.2 三沙湾底栖动物群落对于水产养殖活动的响应
本研究结果显示三沙湾水域大型底栖动物物种数、丰度及生物量在采样站位之间变化范围较大,如丰度和生物量高值可数百倍于低值(两者变化范围分别为25.00~1 075.00个·m-2和0.15~128.22 g·m-2)。然而,站位之间的差异并非由水域类型的差别决定。例如三沙湾底栖生物丰度在网箱养殖、海带养殖、鲍鱼养殖和无养殖水域均值分别为430.98个·m-2、367.19个·m-2、437.50个·m-2和 223.21个·m-2,4种类型水域间无显著差异,两两比较结果显示任意两种水域之间也无统计学差异。在三沙湾内,丰度最大值出现在网箱养殖站位第17号站,可达1 075.00个·m-2,此种高丰度并非由少数机会物种所致,而是由于站位内高数量的丝异须虫、不倒翁虫、欧努菲虫属和索沙蚕科物种共同作用而成。同时,此站位Shannon-Wiener多样性指数为2.82,印证了站位内底栖生物维持在较高的物种多样性水平。生物量最高值也出现在网箱养殖站位(第9号站位),站位内的甲壳动物和棘皮动物对于总生物量贡献较大。
KAPSAR等[24]对饵料的研究表明,网箱养殖饵料的3/4总氮和总磷将排入水域,其中65%总氮和10%总磷沉于海底。海湾内的鲍鱼和海藻养殖则无需投入动物性蛋白。根据PEARSON等[25]提出的经典底栖群落演替理论,三沙湾高强度网箱养殖活动引发AOM的过量沉降应会造成区域内底栖群落呈现较低的生物量和丰度,并引发区域内甲壳动物和棘皮动物等类群数量较少、环节动物多毛类动物数量较多等状况。然而,本研究结果表明三沙湾网箱养殖并未对底栖群落产生显著影响。因此,本文结果与之前较为普遍的网箱养殖为高污染养殖方式的认知存在差异。特别是本研究采集的站位中,丰度和生物量的最大值均出现在网箱养殖站位。同时,本研究的聚类结果表明,即使在相同类型的养殖水体中,底栖群落结构也呈现出较大空间差异,例如无养殖水域、海藻养殖等水域内的站位皆是如此。唐盟[19]对三沙湾底栖环境因子的空间分布特征进行研究,结果显示三沙湾内典型环境因子呈现较为均质性分布的特征,其空间格局并未体现出水域类型之间的差异性,因此本研究揭示的群落参数空间分布特点与环境因子相符。彭广海等[9]利用AMBI、M-AMBI和Shannon-Wiener多样性指数等方法评价水产养殖对于底栖环境影响,结果表明网箱养殖活动并未对底栖环境造成明显的负面影响,本文结果与之一致。
笔者部分未发表的数据表明,三沙湾内氮、磷的主要来源包括水产养殖和来自毗邻陆域的农业面源污染和畜禽养殖排放,水产养殖活动承担了外源营养物质输入的主要贡献量。以2014年数据为例,养殖活动使三沙湾内总氮和总磷分别增加了1.2×104t和2.4×103t。因此,本文认为区域内网箱养殖的环境影响客观存在,且维持在较高强度。但因三沙湾内良好的水动力条件和复合养殖等客观条件,扰动被有效减缓,从而未对底栖群落产生显著影响。
一些研究表明较高强度水产养殖活动对于底栖群落未产生显著影响,如象山港网箱养殖区底栖生物物种数、丰度和生物量均高于自然水域,显示出AOM对于底栖生物多样性的支持功能[26];黄河口较大规模的海水池塘养殖对于邻近区域的影响也限于较小范围和较低程度[27]。然而,也有研究显示,网箱养殖对于底栖群落会产生显著不利影响,如大亚湾大鹏澳内的网箱养殖活动降低了底栖动物的生物量和丰度[14];乐清湾养殖和非养殖区内群落结构差异显著[15]。此种差异性结果的出现表明底栖群落和养殖活动胁迫之间的关系较为复杂,养殖对于群落的影响程度受养殖强度和区域水文动力、沉积物特性等自然地理状况等多方面因素共同影响。
海藻养殖是三沙湾内重要的生产方式之一。本文和象山港的相关研究[22]均表明,部分海藻养殖水域内底栖群落较为退化,显示此种养殖方式对于底栖群落可能产生潜在的负面影响。目前,在国内海藻养殖因其具有消除富营养化的作用,因而多被认为具有积极的生态作用,相关负面的生态效应报道较少。然而,在国外关于海藻养殖引发的负面生态结果已有报道,如在坦桑尼亚沿岸水域,海藻养殖导致底栖生物丰度和生物量降低[28]。因此,本文认为海水藻类大规模养殖对于区域底栖生态环境的影响值得深入研究。
3.3 群落结构和环境因子的相关性
三沙湾内底栖生物和典型环境因子的典范对应分析结果表明,区域内水体温度、盐度、水深和沉积物含水率、粉砂-黏粒含量、总氮和总有机碳等环境因子对于群落的影响较大(图5)。上述环境因子可归纳为两类,第一类环境因子为底栖生物生境的基本理化因子。底栖生境通常包括底层水体和沉积物环境两种类型,水体温度、盐度和水深是水体环境中影响底栖群落最为重要的环境因子,温度和盐度影响生物基本新陈代谢特征,不同物种在长期演化过程中形成独特的温度和盐度适应性,温度和盐度基本决定区域内生物多样性和区系特点;水深是影响水体水文动力特征的重要因素,可影响水动力交换水平,同时也影响有效光照等关键生态条件,通过初级生产途径进而影响底栖群落特征。沉积物含水率、粉砂-黏粒含量为沉积物环境的基本物理特征,影响沉积物中营养物质和污染物质的赋存,从而影响底栖生物群落。第二类环境因子作用于底栖生物的营养功能。例如,总氮和总有机碳的数量特征通常表征生物的食源范围,是影响生物群落的核心过程[29]。
本研究结果和已有研究结果存异,例如ZHOU[6]研究结果表明,在众多环境因子中,水体溶解氧含量、沉积物酸性可挥发性硫化物含量、沉积物氧化还原电位和大型底栖动物群落结构之间相关性较强,而沉积物中总氮和总磷与底栖生物群落无显著相关性。此种区域相同但结果存异的现象表明不同数据来源、不同实验设计方法对于生物群落和环境因子相关性结论的获取存在较大的影响。
在自然条件下,各种环境因子之间协同和拮抗作用复杂,使得影响生物群落的关键环境因子难以甄别。本文揭示在三沙湾水域非核心环境因子也具有较为重要的生态作用,例如底层水体溶解氧和沉积物酸性可挥发性硫化物含量等。日本国内曾颁布“Law to Ensure Sustainable Aquaculture”,该条例将养殖底层水体溶解氧、沉积物酸性可挥发性硫化物特征和大型底栖动物群落作为判定底栖生境质量的3条标准,三者之间具有较强的相关性[30]。在东海区养殖海湾中,象山港网箱养殖区沉积物中总碳、总磷和总有机质和对照区近似相等,但其内的硫化物含量显著较高[26]。三沙湾内底层水体溶解氧和沉积物硫化物对群落影响较小的原因应在于此两种环境因子的分布特点,与日本典型养殖海湾内报道的相关数值相比[11],三沙湾内沉积物硫化物含量较低,而底层水体溶解氧含量较高,同时两者在海湾内的分布皆较为均匀。
三沙湾内大部分数量优势物种和水深、沉积物总氮、总磷、总有机碳和含水率呈现显著的正相关关系,显示三沙湾内营养物质的累积尚未造成区域的过度营养化,目前有机物质的输入对于维持区域生物多样性和种群数量具有积极的支持作用。数量绝对优势物种(如图5中的1、2、4、5号物种等)多位于二维排序轴的中央,体现此类物种对于特定种类环境要素的依赖性较弱,同时也说明此类物种具有较为宽广的生态适应能力。
因采样条件等客观因素限制,本研究选取的环境因素虽较为典型,但数量较为有限(共13个环境因子),虽能基本反映养殖活动的胁迫状况,但未能很好地体现其他外源胁迫的影响程度,例如区域内水上交通、海洋工程建设等人类活动对于底栖环境的扰动作用,此方面研究在后续工作中有待补充。
致谢:日本京都大学横山寿教授对于本研究实验方案设计、样品采集及处理方法制定等工作提供重要指导,中国水产科学研究院东海水产研究所唐盟和温州大学彭广海两位研究生同学协助野外样品采集工作,谨致谢忱。

附表1 三沙湾大型底栖动物物种名录及序号(按优势度降序排列)Appendix 1 Directories and sequence number ofmacrobenthos species in Sansha Bay(descending sort w ith dom inance)
