10个冷季型草坪草品种在高温胁迫下的抗性
2019-09-02徐庆国梁东鸣
赵 娜,徐庆国,苏 鹏,梁东鸣,唐 瑶
(1. 湖南涉外经济学院体育学院,湖南 长沙 410205;2. 湖南农业大学农学院,湖南 长沙 410128)
全球气候变化背景下,各种极端天气层出不穷,极端高温天气也增多,使得原本存在越夏困难的南方冷季型草坪草面临新的挑战[1],尤其是交播草坪中的多年生冷季型草坪草,极端高温会导致交播草坪建植过程中冷季型草坪草成坪不理想或者返青受阻。另外,有研究发现,中高纬度的内陆地区更容易遭到高温胁迫的侵害[2]。我国南方地区位于中纬度的内陆地,高温是限制冷季型向南方地区引种推广以及草坪维护的重要因子。目前,已对冷季型草坪草的高温抗性及其耐热性表现开展了相关研究[3]。早期的研究主要着重于冷季型草坪草在过渡气候带的适应性研究[4-5],随后进行了冷季型草坪草的高温抗性的生理生化、基因表达以及代谢产物研究[6-7],以及在南方若干地区的冷季型草种筛选研究[3],以期得到更耐高温、更适宜在南方种植的冷季型草坪草种。但目前对草坪草引种筛选和耐热性鉴定指标的研究、选育抗热品种及配套技术措施方面的工作还比较少,特别是针对湖南地区的耐热品种筛选工作更少,且已有的评价冷季型草坪草种耐热性的指标数量虽多,但没有进一步划分不同指标对冷季型草坪草种响应高温胁迫时的类型。
本研究以6种(10个品种)冷季型草坪草坪用性状与5个生理生化指标作为综合判断依据,进行了室内培养和大田种植试验观测,以期了解冷季型草坪草耐热性鉴定指标的类型,筛选出适宜湖南种植的冷季型草坪草种,解决冷季型草坪草在湖南地区越夏的问题,为今后草坪建设中的草种选择和草坪草遗传育种研究提供参考。
1 材料与方法
1.1 供试材料
试验所用材料来自于6个冷季型草坪草种中的10个品种,分别为,3个高羊茅(Festuca arundinacea)品种:锐步(Barrera)、易凯(Ikay)、维加斯(Vegas);1个紫羊茅(Festuca rubra)品种:桥港Ⅱ(Bridgeport Ⅱ);2个黑麦草(Lolium pernne)品种:百舸 (Bartwingo)、爱神特 (Accent);2个早熟禾(Poa pratensis)品种:威罗(Wellow)、百思特(Barrister);1个白三叶(Trifolium repens)品种:考拉 (Koala);1个匍匐剪股颖(Agrostis stolonifera)品种:摄政王(Regent)。供试草种均来自北京中种草业有限公司。
1.2 试验地概况
试验于2015年10月至2016年7月在湖南农业大学草业科学系教学科研基地进行。试验地位于湖南省东北部,处在湘江下游和长沙盆地西边,111°53′-114°15′ E,27°51′-28°41′ N。年平均气温17.2 ℃,从5月下旬起,气温显著升高,夏季日平均气温在30 ℃以上有85 d,气温高于35 ℃的炎热日,每年平均大概30 d。试验期间日最高和最低气温变化如图1所示。
1.3 试验设计与指标检测
草坪使用条播法建植,于2015年10月10日在每个试验小区挖深为3~5 cm 的浅沟,将草种撒播沟内,覆土。各试验小区面积为2 m × 1.5 m,完全随机区组排列,每个品种重复3次,常规养护。2015年12月至2016年7月每月观测草坪坪用性状指标。草坪颜色使用TCM500草坪颜色分析仪进行测定,所有小区均随机测定3次,最后计算平均值。草坪绿期使用目测法,以50%草坪草枯黄和返青为准。草坪均一性测定使用目测法,按照1~9级进行分级。草坪颜色无差异,无明显病虫害特征,没有裸地,杂草率< 0.05%,为8~9级;裸地和杂草所占据的面积在1%~15%之间为6~7级;裸地和杂草所占据的面积在16%~30%之间为4~5级;裸地和杂草所占据的面积在31%~45%之间为2~3级;裸地和杂草所占据的面积达到45%以上为1级。草坪抗热性使用目测的方法,用安全越夏率作为主要指标,夏季的耐热性级别和对应判定标准为草坪草70%~100%死亡为1~2级,40%~70%枯黄为3~4级,10%~40%为5~6级,0~10%枯黄现象为7~9级。
2016年3月初,挖取生长在同一生育期的带土草坪草植株,按草种分别移栽于20 cm × 12 cm ×10 cm的塑料盒中,置于人工气侯箱内进行高温培养。分别在25(CK)、30、35和40 ℃温度条件下进行培养,期间进行正常的光照(每天10 h,光强5 000 lx),每个温度下处理7 d,每个处理设3次重复。处理结束后检测各草坪草样品叶片各项生理生化指标。叶绿素含量检测使用丙酮浸提法[8],丙二醛(MDA)含量检测使用双组分分光光度计法[9],游离脯氨酸含量检测使用酸性茚三酮法[10],可溶性糖含量检测使用蒽酮比色法[11],叶片膜质透性使用相对电导率法检测[12]。

图1 2016年4月-7月试验地日最高与最低气温Figure 1 The highest and lowest temperatures of site from April to July 2016
1.4 数据统计与分析
所有试验数据均使用Excel 2007整理并绘制表格,使用SPSS 18.0软件进行单因素方差分析与Duncan多重比较,有关气温的数据使用Sigmaplot 12.5整理作图。
2 结果与分析
2.1 田间生长条件下不同冷季型草坪草坪用性状的差异
2.1.1 不同冷季型草坪草叶片绿度的变化
草坪绿度是指草坪草绿色的深浅,表征草坪观赏质量。从2015年12月到2016年4月,各个草种叶片绿度的数值波动较小(表1),到5月底白天平均最高气温开始达到30 ℃,所有草种叶片绿度均开始下降,尤其以黑麦草、早熟禾和剪股颖的5个品种降低最明显。田间观测发现这5个品种草坪草叶片叶尖出现枯黄的情况;从6月到7月底,白天最高气温逐渐升高,温度达到近40 ℃时,所有草种叶片绿度都大幅度降低,特别是黑麦草、早熟禾以及匍匐剪股颖的几个草种,7月17日观测时由于温度过高,地上部分枯死;而高羊茅和紫羊茅的绿度下降幅度比其他草种小,甚至生长良好。白三叶考拉在用作草坪草时,持续高温下也可以维持较好的长势,且比高羊茅易凯表现更好。
2.1.2 不同冷季型草坪部分坪用指标的比较
4月底各个草种建植的草坪均有较高的均一性级别(表2)。田间观测发现,所有草种叶片颜色浓绿、质地好、细软,植株密度大,外观有吸引力。6月-7月气温回升,全部草种草坪均一性都有所降低,其中以黑麦草、剪股颖和早熟禾所有品种建植的草坪均一性降低最快;根据田间观测,发现其叶片蜷缩、干枯,还有一部分死亡,草坪变得稀疏且有不少杂草长出。但是高羊茅和紫羊茅以及白三叶考拉却没有出现上述情况,均一性降幅也较低。
高羊茅的3个品种、白三叶考拉和紫羊茅桥港Ⅱ的绿期较长,比黑麦草、剪股颖和早熟禾平均长50 d左右,田间观测发现,没有草种全部安全度过夏季。不过,各草种的越夏表现有很大的差异,高羊茅的3个品种、白三叶考拉和紫羊茅桥港Ⅱ明显优于其他草坪草,这与其他坪用指标有较高的一致性。
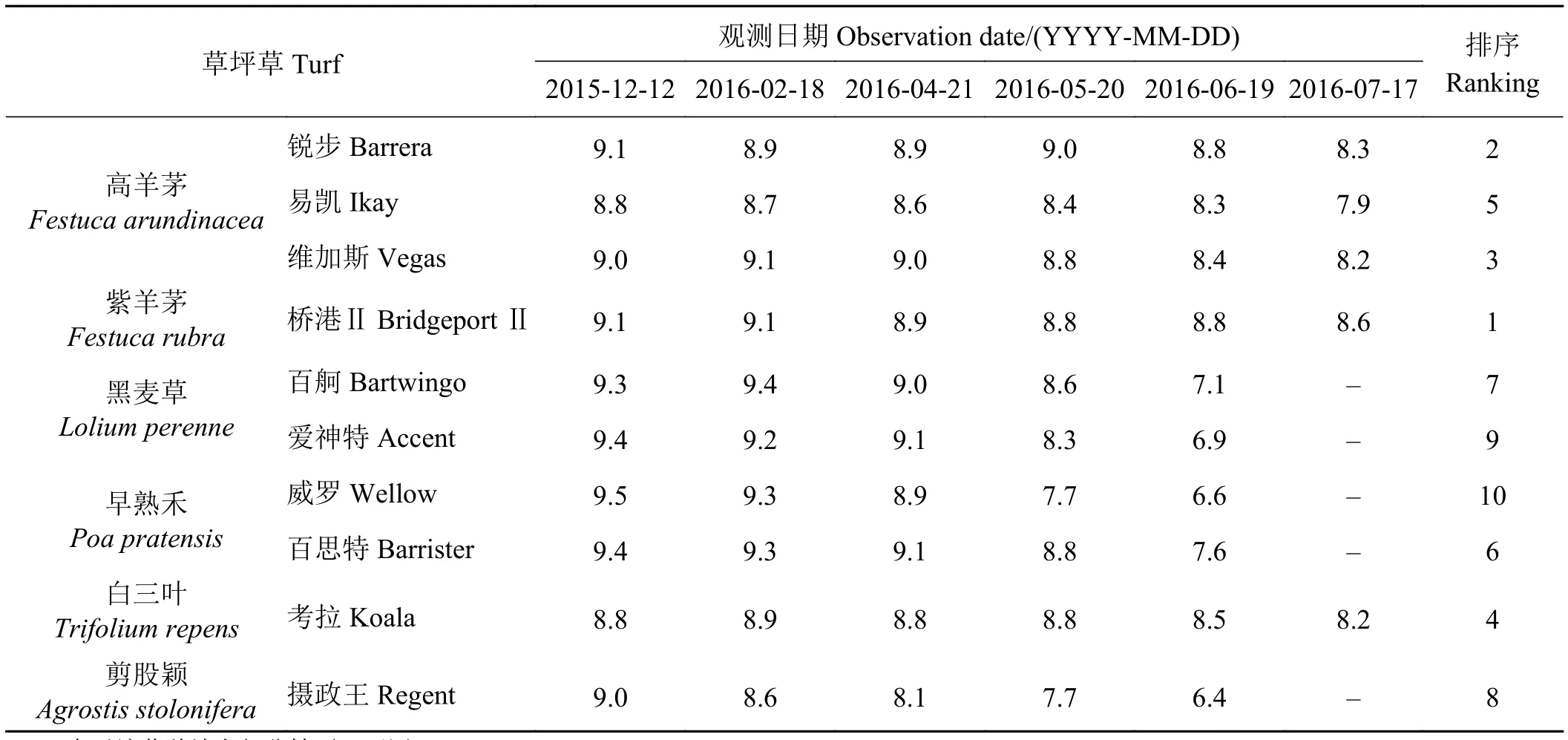
表1 田间生长条件下不同冷季型草叶片绿度的变化Table 1 Changes in leaf greenness of different cold-season grass species during field growth

表2 不同冷季型草种部分坪用指标变化Table 2 Changes in turf indicators of different cold-season grass species during field growth
2.2 室内高温胁迫培养下不同冷季型草抗性
2.2.1 高温对不同冷季型草种叶绿素含量的影响
随处理温度的增加,全部冷季型草种的叶片叶绿素含量均为降低的趋势,但降低幅度有3个梯度,高羊茅、白三叶和紫羊茅叶绿素含量随温度升高下降幅度较小,黑麦草和早熟禾百思特的叶绿素含量随温度升高下降幅度明显,而剪股颖和早熟禾威罗的叶绿素含量降低幅度最大(表3)。
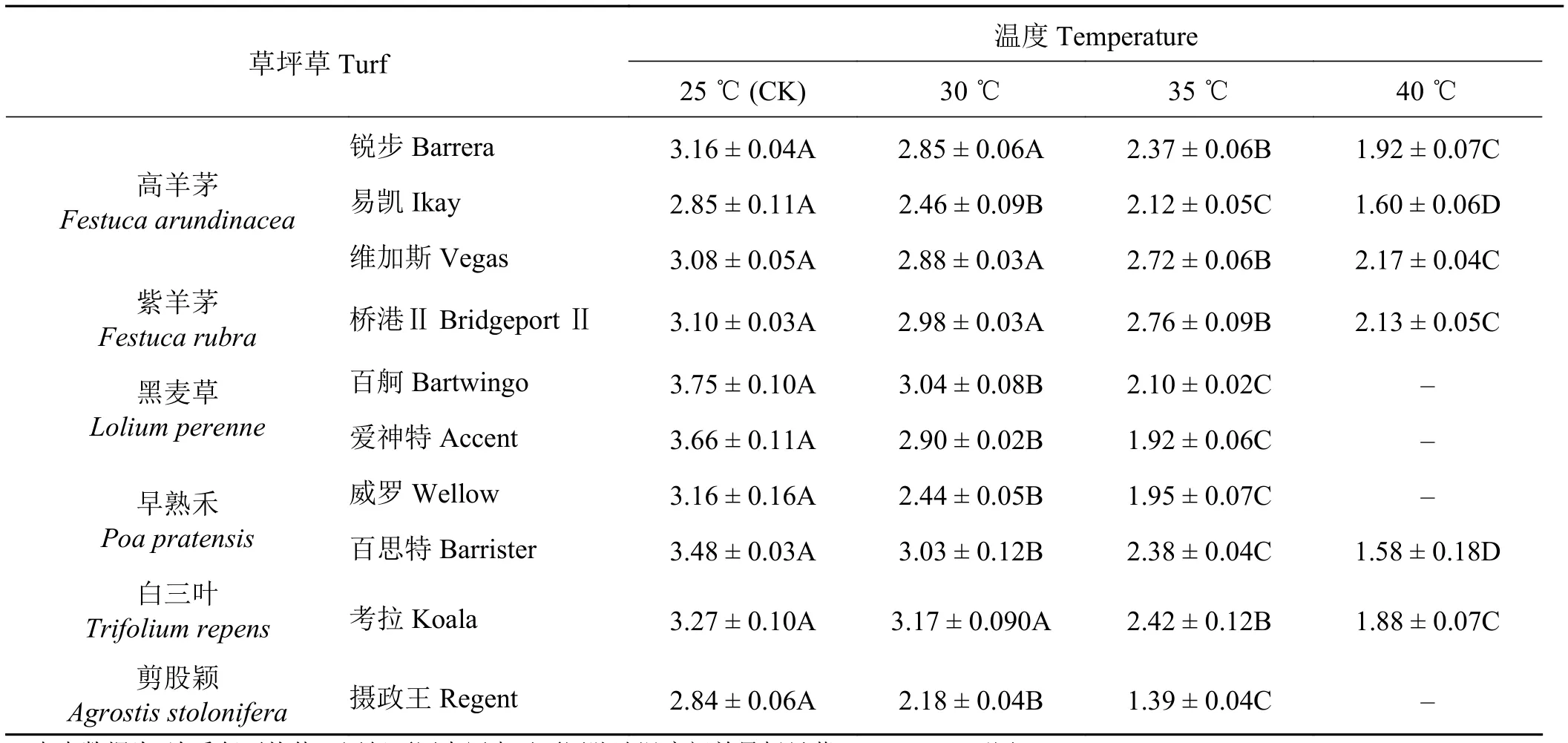
表3 不同温度处理下各草种叶绿素含量的变化Table 3 Changes in chlorophyll content of different grass species under different temperature treatments mg·g-1
锐步、易凯、维加斯和桥港Ⅱ在温度升高至40 ℃时,叶绿素含量分别比对照下降了39.2%、43.9%、29.5%和31.3%,且均有极显著差异(P< 0.01)。百舸、爱神特、威罗和摄政王在温度升高至35 ℃时,同其在对照温度下的叶绿素含量相比,4个品种中叶绿素含量分别下降了44.0%、47.5%、38.3%和51.2%,也均有极显著差异(P< 0.01);在温度升高至40 ℃时,5 d内这4种草均死亡,说明这4个草种高温承受的阈值低于40 ℃。百思特和考拉在温度升高至40 ℃时,叶绿素含量比对照分别降低了54.6%和42.5%,尽管下降较多,然而百思特可以在40 ℃高温下仍然不死,说明百思特比另一个早熟禾品种威罗的高温抗性更好。叶绿素含量随温度增高而降低的幅度越小,说明耐高温胁迫能力越强。因此,单从叶绿素含量的增加来看,不同冷季型草品种的高温胁迫抗性依次表现为维加斯 >桥港Ⅱ > 锐步 > 考拉 > 易凯 > 百思特 > 百舸 > 爱神特 > 威罗 > 摄政王。
2.2.2 高温对不同冷季型草种丙二醛(MDA)含量的影响
随处理温度的升高,全部冷季型草种的叶片MDA含量均表现为增加的趋势(表4)。增加幅度有2个梯度,高羊茅、白三叶、紫羊茅和早熟禾的百思特MDA含量随温度升高增加幅度较小,黑麦草、剪股颖和早熟禾威罗的MDA含量增加幅度较大;但百思特和白三叶考拉在温度达到40 ℃时增幅较大,高于高羊茅和紫羊茅,说明虽然百思特和白三叶对持续的低强度高温有一定的耐受性,但不耐高强度高温。
在4个处理温度下,参试草种叶片中的MDA含量存在明显差异。25 ℃时各冷季型草种叶片的MDA含量差别较小,当温度升高至30 ℃时,除了紫羊茅桥港Ⅱ和早熟禾的百思特,其余草种叶片MDA含量均极显著上升(P< 0.01)。锐步、易凯、维加斯和桥港Ⅱ在温度增加至40 ℃时,与对照相比,叶片内MDA含量分别升高了34.1%、39.9%、32.5%和31.9%,差异极显著;百舸、爱神特、威罗和摄政王当温度升高至35 ℃时,与对照相比叶片MDA含量分别升高43.0%、49.6%、74.2%和67.3%,都有极显著性差异,而在温度升高至40 ℃时这4个草种全部死亡;百思特和考拉在温度达到40 ℃时,与对照相比,MDA含量各自升高了69.4%与63.4%,差异极显著。从高温胁迫下叶片MDA含量的变化看,不同冷季型草种在高温胁迫的抗性大小表现为锐步 > 维加斯 > 易凯 > 桥港Ⅱ > 考拉 > 百思特 >百舸 > 爱神特 > 摄政王 > 威罗。
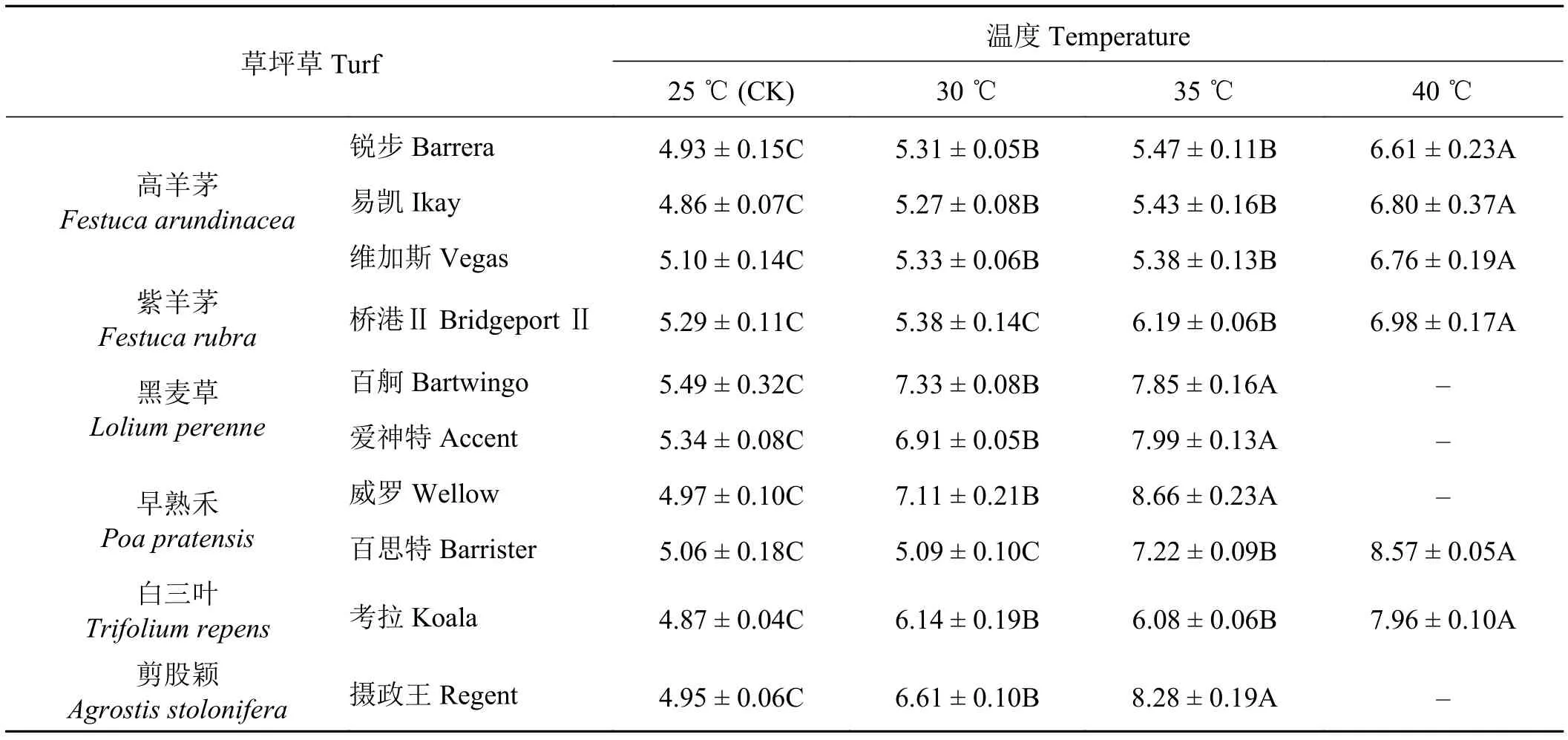
表4 不同温度处理下各草种丙二醛(MDA)含量的变化Table 4 Changes in malondialdehyde (MDA) content of different grass species under different temperature treatments μmol·g-1

表5 不同温度处理下各草种脯氨酸含量的变化Table 5 Changes in proline content of different grass species under different temperature treatments μg·g-1
2.2.3 高温对不同冷季型草种脯氨酸含量的影响
各冷季型草种叶片脯氨酸含量在处理温度从25 ℃升高至30 ℃时有所增加,但增加幅度较小(表5);当温度升高至35 ℃时,除了威罗和摄政王外其余叶片脯氨酸含量都极显著地升高(P< 0.01),升高幅度较大的是高羊茅、紫羊茅、早熟禾百思特和白三叶。温度升高至40 ℃时,黑麦草、早熟禾威罗以及剪股颖摄政王全都死亡,其他草种叶片脯氨酸含量仍然持续升高,其中高羊茅和紫羊茅的游离脯氨酸含量几乎成倍增加。
锐步、易凯、维加斯和桥港Ⅱ在处理温度升高至40 ℃时,与对照相比,叶片脯氨酸含量分别升高583.8%、365.0%、544.1%和513.2%,差异极显著(P< 0.01),百舸、爱神特、威罗和摄政王35 ℃处理下与25 ℃处理相比,叶片脯氨酸含量分别升高266.4%、205.4%、160.5%和50%,差异极显著(P< 0.01),且在温度升高至40 ℃时全部死亡;早熟禾百思特和白三叶考拉在40 ℃温度条件的处理下与对照温度处理相比,其脯氨酸含量分别升高266.2%和162.6%,均有极显著差异,可见,百思特和威罗虽然同属早熟禾,但两者的高温抗性差异非常大,威罗的高温胁迫抗性要远弱于百思特,这与田间观察基本一致;但在强高温胁迫下,两者的抗性比高羊茅和紫羊茅低很多。从高温胁迫下叶片脯氨酸含量的变化看,不同冷季型草品种的高温胁迫抗性强弱表现为维加斯 > 锐步 > 易凯 > 桥港Ⅱ > 百思特 > 考拉 > 爱神特 > 百舸 > 威罗 > 摄政王。
2.2.4 高温对不同冷季型草种可溶性糖含量的影响
各冷季型草种叶片可溶性糖含量在处理温度从25 ℃升高至30 ℃时有所增加,但增加幅度较小(表6);当温度升高至35 ℃时,全部草种叶片可溶性糖含量较对照均极显著升高(P< 0.01),升高幅度较大的是高羊茅、紫羊茅、早熟禾百思特和白三叶。温度再往上升高到40 ℃时,黑麦草、早熟禾威罗和剪股颖摄政王全部死亡,其他草种叶片可溶性糖含量仍然持续,其中高羊茅和紫羊茅的游离可溶性糖含量几乎成倍增加。
在不同的温度处理下,所有草种叶片可溶性糖含量与对照相比均有明显升高。锐步、易凯、维加斯和桥港Ⅱ在40 ℃处理下与对照相比,可溶性糖含量分别升高了277.7%、242.6%、199.0%和252.0%,达到极显著水平(P< 0.01),百舸、爱神特、威罗和摄政王35 ℃与对照相比,可溶性糖含量分别升高了39.4%、32.0%、28.8%和48.7%,差异极显著(P< 0.01),而在温度上升到40 ℃处理后分别死亡;百思特和考拉在25 ℃到40 ℃等不同的温度梯度之间均存在极显著差异,40 ℃与对照相比可溶性糖含量分别升高107.1%和187.7%。从高温胁迫下叶片可溶性糖含量的变化看,不同冷季型草品种的高温胁迫抗性强弱依次表现为维加斯 > 锐步 > 桥港Ⅱ > 易凯 > 考拉 > 百思特 > 百舸 >摄政王 > 威罗 > 爱神特。
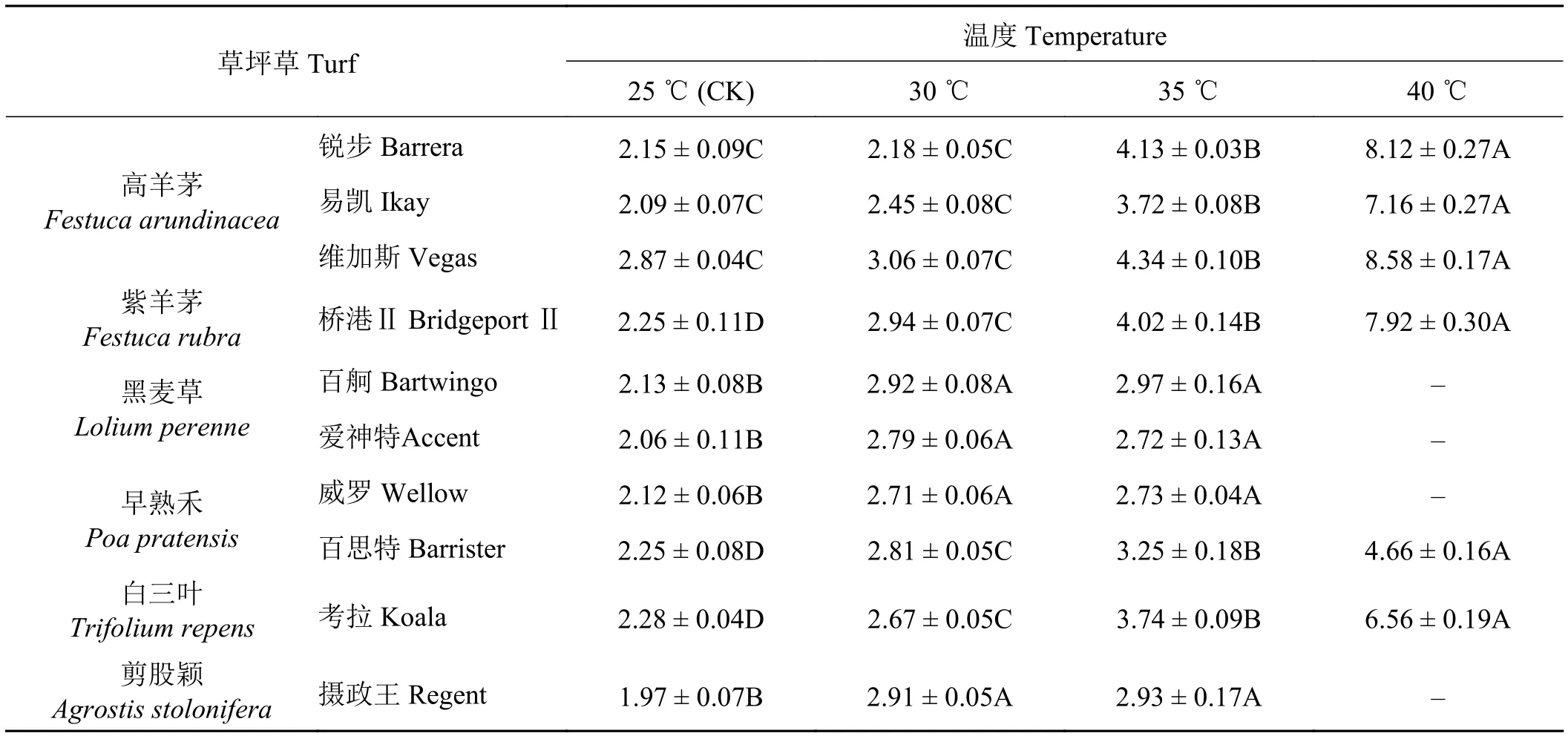
表6 不同温度处理下各草种可溶性糖含量的变化Table 6 Changes in soluble sugar content of different grass species under different temperature treatments mg·g-1
2.2.5 高温对不同冷季型草种相对电导率的影响
不同冷季型草种从25 ℃到30 ℃之间的相对电导率都有缓慢增加,但增幅不大(表7);当处理温度从30 ℃上升到35 ℃,黑麦草百舸和爱神特、早熟禾威罗、剪股颖的相对电导率都有大幅度增加;而高羊茅和紫羊茅的相对电导率则继续平稳增加;至40 ℃时黑麦草、早熟禾威罗、剪股颖摄政王4种冷季型草品种全部死亡。百思特作为早熟禾品种尽管当温度升高至40 ℃时相对电导率升高相当大,受害严重,但是仍然在40 ℃高温下维持7 d,说明在早熟禾品种中,百思特的抗热性较好。
在不同温度处理下,与对照相比,不同草种叶片相对电导率都有极显著增加(P< 0.01)。锐步、易凯、维加斯和桥港Ⅱ相对电导率40 ℃与对照相比分别增加了125.6%、170.0%、107.4%和142.7%,差异极显著;百舸、爱神特、威罗、百思特和摄政王当温度升高至35 ℃时,与对照相比,电导率分别增加了147.4%、216.3%、220.9%和167.0%,差异极显著,且都在温度上升到40 ℃时死亡;百思特和考拉在25 ℃到40 ℃之间的几个温度梯度间相对电导率都有极显著差异,但与百舸、爱神特、威罗、百思特和摄政王相比,受到的伤害要小,因此从相对电导率的角度看,抗性比这几个草种要好,但比高羊茅和紫羊茅抗性差。从高温胁迫下叶片相对电导率的变化看,不同冷季型草品种的高温胁迫抗性大小依次表现为, 维加斯 >锐 步 > 桥港Ⅱ > 易凯 > 考拉 > 百思特 > 百 舸 >摄政王 > 威罗 > 爱神特。

表7 不同温度处理下草种相对电导率的变化Table 7 Changes in electric conductivity of different grass species under different temperature treatments%
3 讨论
3.1 高温胁迫对冷季型草坪用质量的影响
草坪坪用质量是草坪草在成坪后叶片的绿度、质地、均一性和绿期长短等一系列评价指标的综合体现[13]。暖季型草坪草在夏季高温条件下具备较好的自我调节能力和适应能力,与暖季型草坪草不同,冷季型草坪草对高温更加敏感,更容易受高温影响而枯黄、死亡,从而降低草坪的观赏性和使用功能[14-15]。另一方面,冷季型草坪草不同草种之间的坪用性状在高温胁迫下的表现也有较大的差异[6,10]。本研究所使用的6个草种在田间受高温影响后的坪用性状表现可以分为两大类,分别是坪用质量较好的高羊茅、紫羊茅和白三叶,以及坪用质量在高温胁迫下较差的黑麦草、早熟禾和剪股颖。
3.2 冷季型草坪草对高温胁迫的响应
如果植物持续地暴露在高温之下,其细胞中的许多代谢活动将会被剧烈影响,甚至失调。这些持续的或者高强度的高温将损伤细胞膜,致使细胞膜的半透性失效,最终致使细胞膜中物质渗漏到细胞外[16-17]。本研究所检测的5个生理指标中,冷季型草坪草受高温胁迫后代谢活动失调的直接表现是丙二醛的积累和相对电导率的升高。丙二醛是植物细胞膜脂过氧化的重要产物,会引发细胞内相关酶反应,并且是造成细胞膜损伤最直接的因素[18]。相对电导率的高低直接表征着植物细胞渗透势的高低,是衡量细胞膜的功能是否正常发挥的重要指标[19]。而丙二醛含量增加和相对电导率的变大间接指示出叶片膜透性的增大,是衡量植物细胞受损害程度的指标。本研究中,高羊茅、紫羊茅、早熟禾百思特和白三叶的丙二醛含量和相对电导率随温度的增高而增加的幅度比其他草种小很多,这表明这几个草种在高温胁迫下受到的损害程度最低。另一个体现植物抗损害能力的指标是叶绿素含量的变化。叶绿体色素含量影响光合作用,其含量的减少会导致光合产物减少,进而影响植物正常生长。本研究中,高羊茅、紫羊茅和白三叶的叶绿素含量随温度的增高而降低的幅度比其他草种小很多,也表明这几个草种在高温胁迫下受到的损害程度最低。因此,丙二醛含量、相对电导率和叶绿素含量的变化可以作为衡量冷季型草坪草高温抗性的抗损害能力筛选指标;丙二醛含量和相对电导率在高温胁迫下增加的幅度越小、叶绿素含量在高温胁迫下下降的幅度越小,速度越慢,表明草种的抗损害能力越强。
细胞膜功能完整是确保植物正常进行光合作用和呼吸作用的关键[20]。一旦植物细胞膜受到损伤,相关功能和结构不完整,植物的逆境适应与恢复机制就会启动。当植物遭受逆境胁迫时,细胞内的脯氨酸合成酶中存在的抑制脯氨酸合成的机制会减弱,造成体内游离脯氨酸的合成增多[21],从而增加细胞内的渗透压,弥补细胞膜透性失效造成的细胞或组织渗透压降低,减少伤害;同时,在环境刺激下,细胞还会合成各种糖类和醇类等低分子量的有机化合物,它们也具有渗透调节、细胞功能保护、细胞组分完整性的维持等功能[22]。因此,高温逆境下冷季型草坪草脯氨酸和可溶性糖含量的升高是其进行自我恢复的生理途径,属于逆境调节能力。本研究中,随着处理温度的上升,各草种脯氨酸含量和可溶性糖含量均升高,但以高羊茅、紫羊茅、早熟禾百思特和白三叶的脯氨酸含量和可溶性糖含量随温度的增高而增加的幅度比其他草种要大,且当温度提升到40 ℃时,这几种草种叶片脯氨酸含量和可溶性糖含量有极大幅度的增加,其他草种死亡。可见,高羊茅、紫羊茅、早熟禾百思特和白三叶的逆境调节能力大于其他草种,具有更好的高温抗性。本研究结果与已有研究结果一致[5,24-25]。
植物的抗性能力强弱是植物应对逆境时综合能力的体现,不可能由单一指标进行准确判断[23]。冷季型草坪草坪用性状作为草坪建植质量,可以直观展示冷季型草高温抗性强弱。湖南地区从5月下旬起,气温显著升高,夏季日平均气温在30 ℃以上有85 d,气温高于35 ℃的炎热日,年平均大概30 d。这期间草坪草均受到高温胁迫,其中高羊茅的3个品种、白三叶考拉和紫羊茅桥港Ⅱ的综合坪用性状,比黑麦草、剪股颖和早熟禾好。这一结果与室内培养试验设定的高温梯度胁迫结果一致。本研究综合检测了冷季型草坪草高温胁迫下可溶性糖含量、游离脯氨酸含量、相对电导率、叶绿素含量、丙二醛含量5个生理生化指标的变化,发现这5个指标均能较好地区分不同草种耐高温胁迫能力的强弱,且这5个指标分属冷季型草高温抗性的两大类型,分别为抗高温损害能力、逆境调节能力。因此,这5个指标可以用来综合鉴定不同冷季型草坪草高温抗性能力的高低,从而筛选出具有较强耐热性的草种用于草坪建植。
4 结论
本研究选取叶片可溶性糖含量、游离脯氨酸含量、相对电导率、叶绿素含量、丙二醛含量5个指标,对10个冷季型草种在室内持续高温胁迫下的生理变化进行检测,并结合田间试验,以此来判断10个草种的高温抗性能力的强弱,筛选出适合湖南地区栽培的冷季型草种。
1)经分析,冷季型草坪草抵御高温胁迫的生理机制可以分为2个类型,一为抗高温损害能力,二是逆境调节能力;选取的5个判断指标中,丙二醛含量、叶绿素含量和相对电导率属于抗高温损害能力的评价指标;可溶性糖含量和游离脯氨酸含量属于植物的逆境调节能力评价指标。
2)室内人工模拟高温胁迫试验与田间试验结果比较吻合,无论是根据田间的高温胁迫下各草种表型差异,还是根据室内人工模拟实验检测的各草种生理指标差异均表明,这10个草种的高温抗性 强弱 表现为 维加 斯 > 锐步 > 桥 港Ⅱ > 易凯 >考拉 > 百思特 > 百舸 > 爱神特 > 威罗 > 摄政王。且大致分为3个等级:维加斯、锐步、桥港Ⅱ和易凯为一个等级,高温抗性最高;考拉和百思特为一个等级,高温抗性较好;百舸、爱神特、威罗和摄政王,高温抗性最差。
