百脉根PIN基因家族的鉴定与分析
2019-08-23丘日光杨仕梅吴佳海
吕 赟,丘日光,杨仕梅,吴佳海,宋 莉
(1.贵州大学 生命科学学院/农业生物工程研究院/山地植物资源保护与种质创新教育部重点实验室/贵州省农业生物工程重点实验室,贵州 贵阳 550025; 2.贵州省草业研究所,贵州 贵阳 550006)
植物对外界环境变化的适应性生长发育与激素调控密切相关[1-2],生长素(Auxin,IAA)是调控该过程的关键激素之一[3-5],在植物生长发育过程中呈浓度梯度的时空不对称分布[6],这与其极性运输方式有关[7]。植物生长素极性运输由生长素输出载体蛋白PIN-formed (PIN)家族、AUXIN RESISTANT1/LIKE AUX1家族(AUX1/LAX)和 ATP-Binding Cassette subfamily B/P-glycoprotein家族(PGP/ABCB)3类蛋白质介导[8],其中PIN和PGP/ABCB属于外向型运输蛋白质,AUX1/LAX属于内向型运输蛋白质。PIN基因结构及其编码的蛋白质特性是其实现生物学功能的基本前提,PIN基因家族的跨膜蛋白是植物特有的生长素输出载体蛋白,几乎存在于所有的植物中,在生长素极性运输中发挥关键作用[9-10]。
百脉根(Lotuscorniculatus)为豆科百脉根属多年生草本植物,其营养丰富、皂素含量低、适口性好、采食率高,是一种优质牧草[11-12]。同时,因其匍匐生长、覆盖度好,根系较为发达,自繁能力强,在改造小流域、保持水土和提高经济效益中也有着重要价值[13-14]。此外,百脉根还具有再生力强、自交结实、遗传转化效率高、易得到稳定遗传的后代等优点,常用作豆科模式植物进行生物学及基因组学研究[15-17]。PIN基因在拟南芥和水稻等模式植物中研究得较为清楚,但在百脉根中鲜见报道,鉴于此,利用拟南芥PIN蛋白家族信息,在百脉根全基因组数据中对其PIN基因家族成员(LjPIN)进行鉴定,并分析该基因家族成员的进化关系、编码的蛋白质理化性质、顺式作用元件组成及组织差异表达等,旨在为百脉根LjPIN的功能研究奠定基础。
1 材料和方法
1.1 目的序列来源
百脉根PIN基因及编码蛋白质序列来自百脉根全基因组数据库(http://www.kazusa.or.jp/lotus/),拟南芥PIN蛋白家族序列来自KEGG数据库(http://www.kegg.jp/),水稻PIN蛋白家族序列来自Pfam数据库(http://pfam.xfam.org/family/PF03547/),大豆PIN蛋白家族序列参考WANG等[18]的报道。
1.2 PIN基因筛选、鉴定及定位分析
通过构建隐马尔科夫模型,用Pfam数据库(http://pfam.xfam.org/)含Mem_trans domain(PF03547)的模型与百脉根全基因组序列构建比对库,筛选鉴定百脉根PIN基因;通过在线软件MG2C(http://mg2c.iask.in/mg2c_v2.0/)对获得的PIN基因进行染色体定位,基因命名根据所在染色体位置进行。
1.3 PIN基因结构和进化分析
从数据库(http://www.kazusa.or.jp/lotus/)获得百脉根PIN基因的外显子和内含子数据并进行分析,用GSDS 2.0基因结构显示系统(http//gsds.cbi.pku.edu.cn/)绘制基因结构图。利用MEGA 7.0邻接法(Neighbor-joining,NJ)构建百脉根、拟南芥、水稻和大豆的PIN蛋白进化树,Bootstrap值设置为1 000,缺口设置“pairwise deletion”,采用“poisson model”验证可信度。
1.4 PIN蛋白基本特征分析
采用ExPASy ProtParam(http://cn.expasy.org)程序对百脉根PIN基因家族编码的蛋白质分子质量、理论等电点、不稳定系数和亲水性指数进行分析;利用SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测蛋白质二级结构;通过MEME工具(http://meme.nbcr.net/meme/)分析蛋白质保守基序(Motif),设置基序数量15个,其余参数为默认条件。
1.5 PIN基因家族顺式作用元件分析
参考文献[19],利用在线预测工具PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对获得的百脉根PIN基因上游2 000 bp序列和主要的顺式作用元件进行预测和分析。
1.6 PIN基因家族组织差异表达分析
从基因芯片数据库(https://ljgea.noble.org/v2/)获得百脉根PIN基因在根、茎、叶、花等器官及不同时期种子的表达量数据,并进行分析、作图。
2 结果与分析
2.1 百脉根PIN基因家族的结构特征和染色体分布情况
经比对共获得35条LjPIN序列(表1),属于27个候选基因,其中,LjPIN2、LjPIN6、LjPIN7、LjPIN19、LjPIN25和LjPIN27基因各2条LjPIN序列,LjPIN26基因有3条LjPIN序列,其他LjPIN基因各1条LjPIN序列。Lj0g3v0172269、Lj0g3v0320849、Lj1g3v0264160、Lj3g3v3735560、Lj5g3v0323470、Lj5g3v1174510和Lj5g3v2166080共7条序列存在可变剪切。通过Pfam数据库对所有候选基因保守结构域进行分析发现,27个候选基因均含有PIN基因家族保守结构域Mem_trans(PF03547)。
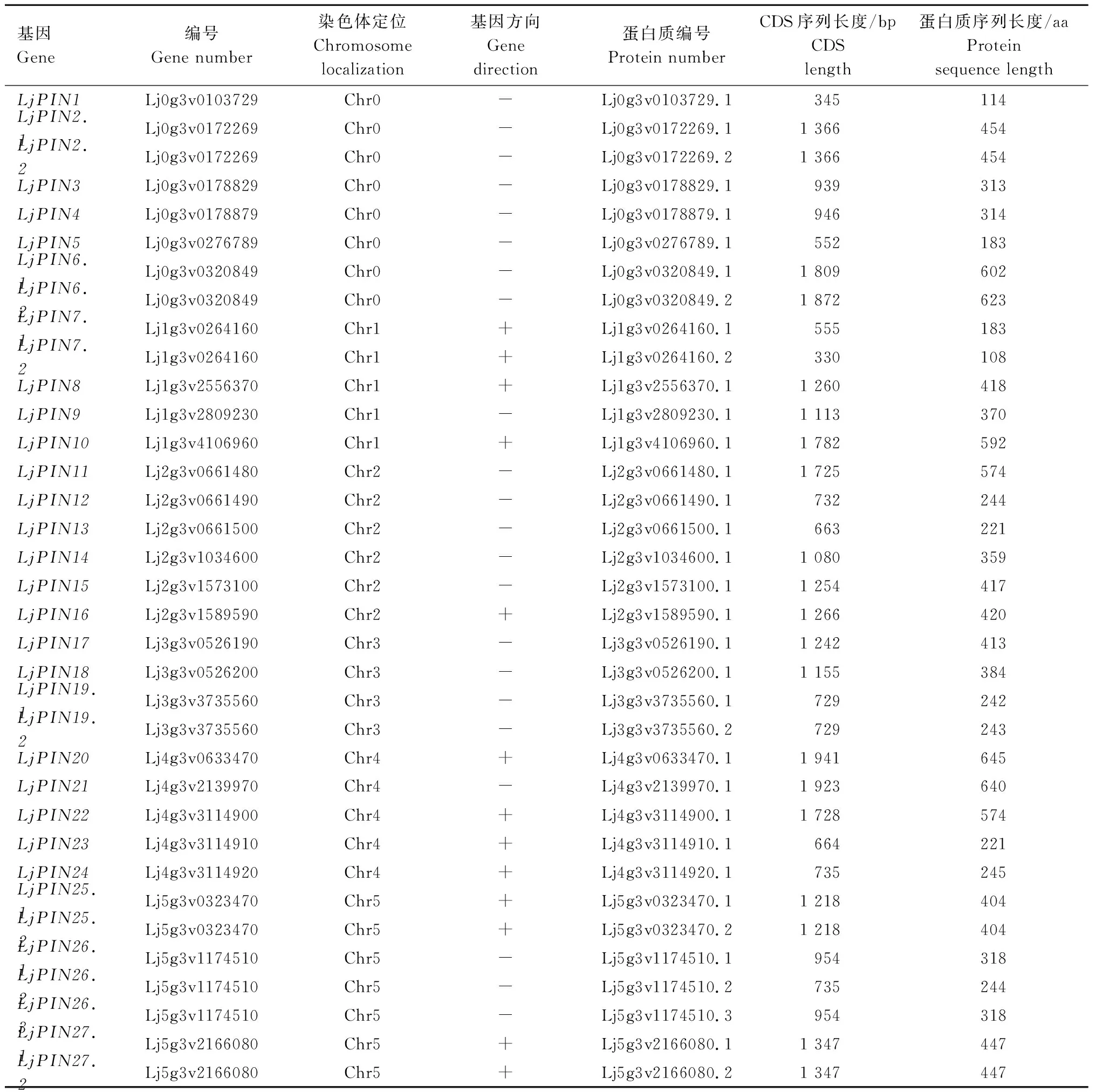
表1 LjPIN基因家族基本信息Tab.1 Basic information of LjPIN gene family members
注:+表示与染色体测序方向一致;-表示与染色体测序方向不一致。
Note: +means consistent with chromosome sequencing direction; - means inconsistent with the direction of chromosome sequencing.
根据百脉根基因组信息分析27个PIN基因在染色体上的分布情况。百脉根共有6条染色体(Chr1—Chr6),另定义Chr0为无法定位到染色体上的基因所虚构的一条染色体。分析表明,在Chr1、Chr2、Chr3、Chr4、Chr5上分别分布着4、6、3、5、3个PIN基因, Chr6上无PIN基因分布,其中Lj0g3v0103729、Lj0g3v0172269、Lj0g3v0178829、Lj0g3v0178879、Lj0g3v0276789和Lj0g3v0320849存在于Chr0上(图1)。
2.2 百脉根PIN基因家族结构组成和进化地位
对鉴定得到的27个LjPIN基因进行系统进化、内含子和外显子分析。结果(图2)表明,百脉根PIN基因家族除了LjPIN4、LjPIN13、LjPIN23这3个以外,其他均含1~10个内含子和1~10个外显子,其中LjPIN1和LjPIN2还存在非编码区(UTR)。根据AtPIN与LjPIN的进化关系,参考文献[20],可以将LjPIN分为两大类,其中LjPIN3、4、6、10、11、13、20、21、22、23为Long PIN,其余的LjPIN成员为Short PIN。
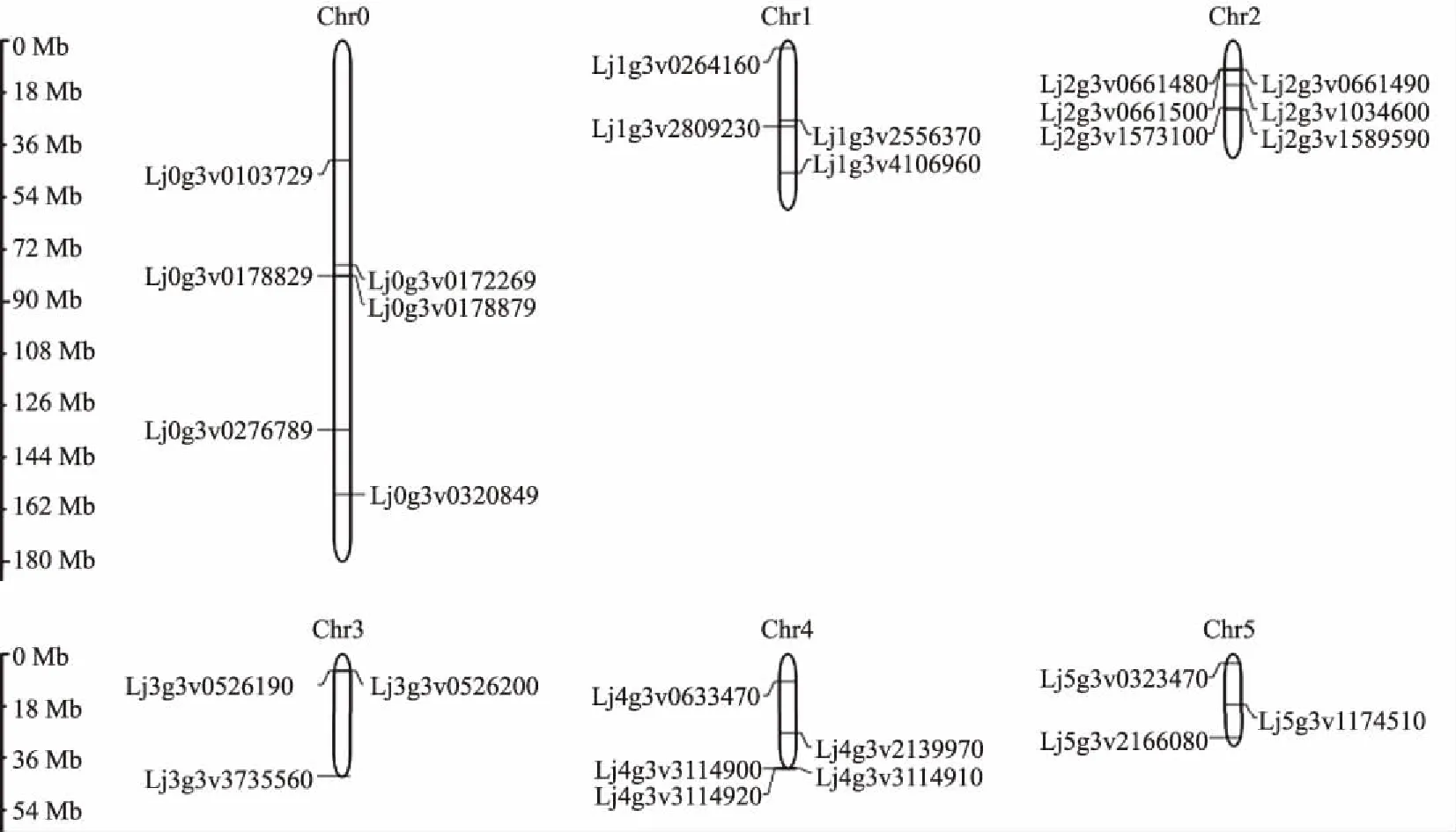
Chr0为虚拟的一条染色体Chr0 is a virtual chromosome图1 LjPIN基因的染色体定位分布Fig.1 Location map of LjPIN in chromosome

图2 LjPIN的基因结构和系统发育关系Fig.2 Gene structure and phylogenetic relationship of LjPIN
为了研究百脉根PIN基因与其他植物的亲缘关系,对植物PIN蛋白家族进化进行探讨。利用PIN基因研究较多的拟南芥、水稻和大豆3种植物与百脉根共78个PIN蛋白序列构建系统进化树(图3)。分析发现,百脉根LjPIN蛋白与拟南芥AtPIN、大豆GmPIN的进化关系较接近,而与水稻OsPIN的进化关系较远。其中16个LjPIN未能与其他物种的PIN聚类到一起,可能与物种分化后因不同选择压力导致的进化方向不同有关,可作为百脉根LjPIN基因鉴定的分子特征。

•表示未能与其他物种的PIN聚类到一起的16个LjPIN•means 16 LjPINs that failed to cluster with PINs from other species图3 拟南芥、大豆、水稻和百脉根PIN蛋白的系统进化树Fig.3 Phylogenetic tree of PIN proteins from Arabidopsis thaliana,soybean,rice and Lotus corniculatus
2.3 百脉根PIN基因家族编码蛋白质序列特性
蛋白质序列特性分析(表2)表明,LjPIN蛋白序列长度在108~645个氨基酸,其中最长的是LjPIN20,长度为645个氨基酸,蛋白质分子质量为70.112 00 ku;最短的是LjPIN7.2,长度为108个氨基酸,分子质量为12.098 51 ku。35个PIN蛋白的理论等电点在5.33~9.94,除了LjPIN1、LjPIN2、LjPIN3、LjPIN5、LjPIN15、LjPIN16和LjPIN27为酸性蛋白质外,其他均为碱性蛋白质。不稳定系数比较发现,除了LjPIN8、LjPIN11、LjPIN12、LjPIN15、LjPIN16、LjPIN19.2、LjPIN21、LjPIN22和LjPIN26.2为不稳定蛋白质(不稳定系数>40)外,大部分PIN蛋白为稳定蛋白质。亲水性指数分析得到,LjPIN1、LjPIN2、LjPIN5、LjPIN7.1、LjPIN8、LjPIN9、LjPIN14、LjPIN15、LjPIN16、LjPIN17、LjPIN18、LjPIN25、LjPIN27蛋白均为疏水蛋白质,其余LjPIN蛋白为两性蛋白质,亲水性介于-0.5~0.5(GRAVY为负值表示亲水性,正值表示疏水性)。
蛋白质结构分析(表3)表明,LjPIN蛋白由α-螺旋、延伸链、β-转角、无规卷曲4种二级结构元件组成,并以α-螺旋和无规卷曲2种元件为主。LjPIN1、LjPIN2.1、LjPIN2.2、LjPIN5、LjPIN7.1、LjPIN7.2、LjPIN8、LjPIN9、LjPIN13、LjPIN14、LjPIN15、LjPIN16、LjPIN17、LjPIN19.1、LjPIN19.2、LjPIN23、LjPIN25.1、LjPIN25.2、LjPIN27.1和LjPIN27.2等20个蛋白质中的α-螺旋占比高于无规卷曲,其他LjPIN蛋白中则是α-螺旋占比低于无规卷曲。

表2 LjPIN蛋白的基本特性Tab.2 Basic characteristics of LjPIN proteins
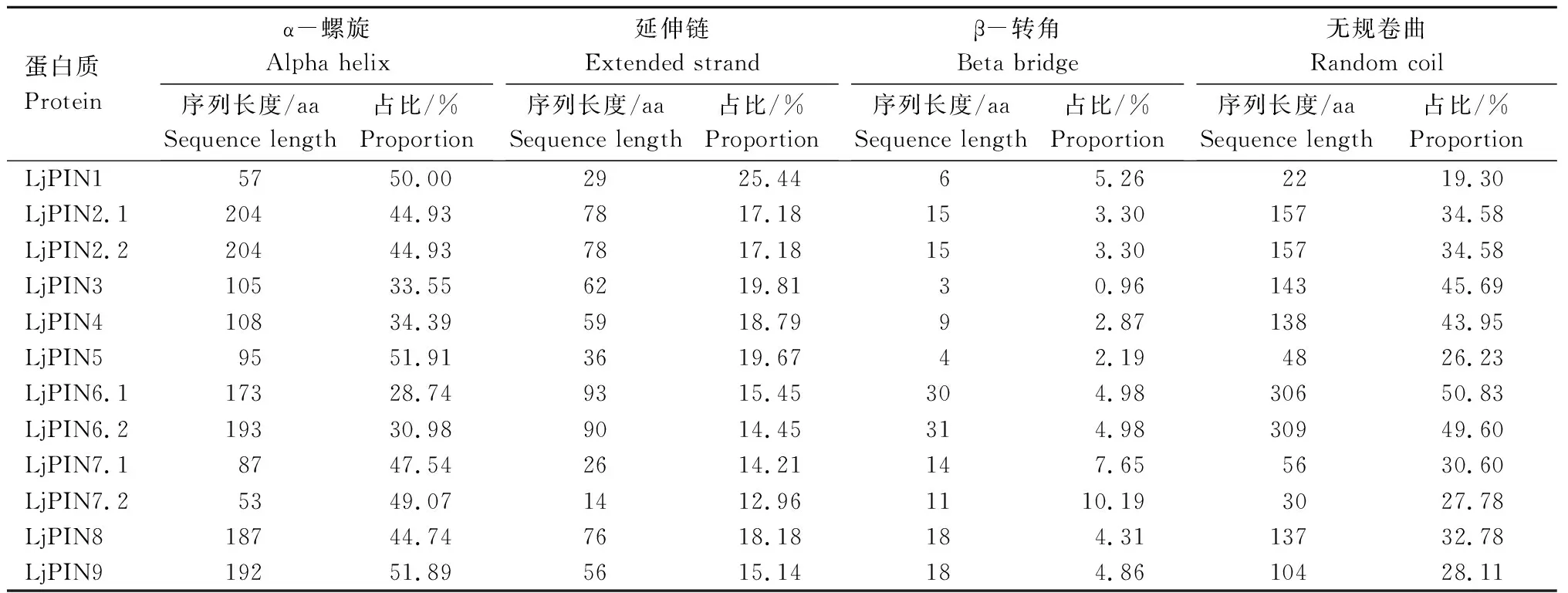
表3 LjPIN蛋白的二级结构 Tab.3 Secondary structure of LjPIN proteins

续表3 LjPIN蛋白的二级结构 Tab.3(Continued) Secondary structure of LjPIN proteins
保守基序分析(图4—5)发现,35个LjPIN蛋白的保守基序数量介于2~9个,其中,LjPIN1的保守基序数量最少(2个),LjPIN20的保守基序数量最多(9个)。基序Motif6、Motif7、Motif9、Motif10和Motif13在LjPIN基因编码的蛋白质中分布最广,分别存在于15个LjPIN基因中,表明这些基序在LjPIN蛋白的进化上具有相对保守性。

氨基酸位置 Amino acid position图4 LjPIN蛋白的Motif分布Fig.4 Motif distribution of LjPIN proteins

图5 LjPIN蛋白的关键MotifFig.5 Key motif of LjPIN proteins
2.4 百脉根PIN基因家族顺式作用元件组成
通过对PIN基因上游2 000 bp序列进行分析发现(图6),LjPIN基因家族除了存在大量的顺式作用基本元件CAAT-box和TATA-box外,还存在光合反应、激素调控和生长发育相关的3类顺式作用元件,具体包括:(1)光响应调节元件G-Box;(2)生长素响应元件TGA-elemnt、茉莉酸甲酯(MeJA)反应元件TGACG-motif、水杨酸反应元件TCA-element、脱落酸反应元件ABRE和赤霉素反应相关元件(TATC-box、P-box和GARE-motif);(3)参与胚乳表达的调节元件GCN4-motif、参与分生组织表达的调控元件CAT-box、参与栅栏叶肉细胞分化的HD-Zip1元件。以上研究表明,LjPIN基因家族在百脉根中以激素调控为主,与植物生长发育过程密切相关,其可能通过对生长素与其他激素的共同调控作用实现对植物生长发育的调控。
2.5 百脉根PIN基因家族组织表达差异情况
对百脉根的根、茎、叶、花及不同形成时期种子(按授粉后天数计算)的芯片数据进行分析,结果如图7所示。由图7可以看出,27个PIN基因中仅有11个在9个样本中出现组织差异表达(上调),其中,PIN2.1、PIN6.1、PIN8、PIN9、PIN15和PIN25.1在叶中高表达;PIN2.1、PIN6.1、PIN8、PIN9和PIN14在茎中高表达;PIN2.1和PIN8在花中高表达;PIN2.1、PIN6.1、PIN8、PIN10和PIN21在根中高表达;PIN2.1、PIN8和PIN18在授粉后10、12、14 d的种子中高表达,PIN2.1、PIN8、PIN18和PIN25.1在授粉后16、20 d的种子中均高表达。

图6 LjPIN基因的顺式作用元件位点组成Fig.6 The cis-acting element site of LjPIN genes

图7 LjPIN基因的时空差异表达变化Fig.7 Expression of LjPIN genes in different tissues and seed maturation stages
3 结论与讨论
PIN蛋白是调控植物生长素极性运输的重要载体元件,参与调控植物生长发育。拟南芥和水稻等模式植物中的PIN基因研究得较为清楚,但在百脉根中尚未见报道,研究百脉根LjPIN基因对其功能鉴定和利用具有重要意义。本研究从百脉根基因组中共鉴定出了27个PIN基因,编码35个PIN蛋白,高于拟南芥(8个)、水稻(12)和大豆(23个)中的PIN成员数量[10,18,21]。系统进化分析发现,LjPIN与拟南芥和大豆PIN基因家族的进化关系最为密切,LjPIN蛋白大多为碱性稳定蛋白质,这与其他植物中的相关报道一致[22-23]。百脉根LjPIN基因的顺式作用元件中含有光响应、激素调控相关及生长发育响应元件,表明LjPIN的生物学活性较为广泛;百脉根PIN基因中的激素调控元件除了生长素类之外,还包括茉莉酸甲酯、水杨酸、脱落酸、赤霉素等其他激素的响应元件,这表明LjPIN基因家族的功能涉及到多种激素调控,推测其在调控生长素的分布后,进而在局部与其他激素协作或拮抗,共同调控植物的生长发育。本研究发现,百脉根LjPIN基因由多种顺式作用元件组成,这为植物体中生长素与其他植物激素通过交互响应调控植物生长发育[5]提供了重要依据。HE等[24]发现,棉花PIN基因家族中仅有2个PIN基因对棉花的主根、侧根发育起关键作用。WANG等[18]发现,大豆PIN基因存在的表达类型、时间和空间差异性对每个组织的发育过程均至关重要,而来自不同组织的PIN基因协同表达,有助于大豆进化过程的灵活性及多变性。本研究中,在百脉根的根、茎、叶、花和不同发育阶段的种子中,仅有11个LjPIN基因出现上调差异表达,表明LjPIN基因存在表达类型差异和时空表达特异性,这种差异可能与不同PIN基因对不同部位和不同时期的调控有关,百脉根LjPIN基因家族与上述棉花[24]和大豆[18]中PIN基因家族的表达类型差异和空间表达差异一致,而这种一致性是各种植物中该基因家族功能丰富和满足特定发育部位和阶段所必需的。综上,百脉根中存在PIN基因家族,该家族基因具有典型的结构特征和特定的表达特性,对进一步研究百脉根LjPIN奠定了基础,也为其他植物PIN基因家族的研究提供了参考信息。
