曼陀罗对土壤镉污染的响应及镉累积特征
2019-08-20周振张彪杨海涛师振亚包环宇苗利娟杨素勤
周振 张彪 杨海涛 师振亚 包环宇 苗利娟 杨素勤

摘要:为了探究镉污染土壤上曼陀罗生长及镉吸收和累积特征,设置对照和污染土壤组,采用盆栽方式对曼陀罗的生长状况、镉含量、生物累积量等指标进行分析。结果表明,相比于对照组,污染土壤组曼陀罗的株高和根长受到一定的抑制,但根部和茎部生物量并无明显减少,其生长未出现受害症状。移栽污染土壤60 d内,曼陀罗叶绿素a和叶绿素b并无显著降低,类胡萝卜素也出现相同规律。本研究中曼陀罗根、茎和叶镉含量最高可达38、43、47 mg/kg,曼陀罗的根部富集系数最高达2.69,地上部转移系数最高达1.40,曼陀罗地上部的镉累积量占总量的70%以上。因此,曼陀罗虽未满足镉超富集植物的标准,但可以作为修复植物用于镉污染土壤的治理。
关键词:植物修复;镉;曼陀罗;富集系数;叶绿素含量
中图分类号: X53;X173文献标志码: A
文章编号:1002-1302(2019)08-0269-04
近年来,土壤重金属污染问题引起人们广泛的关注。据《全国土壤污染状况调查公报》显示,我国土壤点位超标率为16.1%,污染类型以无机污染为主,其中,镉(Cd)的点位超标率达7.0%[1]。目前,对重金属污染土壤治理方法的研究已得到各界重视[2-3]。就耕地而言,重金属污染土壤的生物修复成为具有巨大发展前途的治理技术之一。植物修复技术是一种利用植物从环境中吸收重金属、放射性核素等有害物质,并转化为环境友好的代谢产物的治理方式[4]。随研究深入,科研工作者已发现多种重金属超富集植物,如李氏禾[5]、龙葵[6]和蜈蚣草[7]等。但是,这些植物因受地域和气候等因素限制,暂时无法广泛种植,难以大面积修复重金属污染土壤。因此,寻找符合当地生态条件、生物量足够大、重金属累积量可观的植物成为土壤修复成功的关键因素之一。
曼陀罗(Datura stramonium L.)别称醉心花,生长迅速、生物量大、适应性强,有一定的经济效益,可作为锰、铬、铜和砷污染土壤的修复植物[8-9]。但是,目前还未见曼陀罗用于镉污染土壤修复的报道。有研究表明,曼陀罗叶片镉含量[LL]为其他野生植物镉含量的数十倍[10]。董林林等研究表明,曼陀罗有较强的镉富集能力,其地上部镉含量最高可达 115 mg/kg[11]。但是关于曼陀罗对镉富集累积的研究还主要集中在某一时期,难以找出最佳的时间点,进行收割或者调控,用于土壤修复。因此,本试验侧重研究土壤镉污染对曼陀罗的生长和镉吸收的影响,重点探讨不同时间曼陀罗的生物量累积和富镉特性,以期为曼陀罗修复重金属镉污染土壤提供参考和依据。
1材料与方法
1.1供試植物及土壤
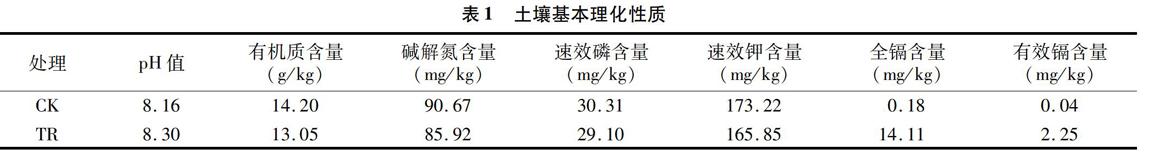
供试野生曼陀罗种子采自豫西某铅锌冶炼厂周边。供试清洁土壤和镉污染土壤分别采自豫西某铅锌冶炼厂西南 25 km 和西北1 km处,分别用CK和TR表示,均采集0~20 cm 表层土壤,风干、碾碎、过4 mm尼龙筛。土壤基本理化性质见表1。
1.2试验设计
试验于2015年5—8月在河南农业大学科教园区进行。选择饱满均一的种子,用5% NaClO溶液消毒30 min,再用10% NaOH浸泡90 min破除休眠[12],经去离子水冲洗、25 ℃遮光催芽后,移至基质育苗盘中,置于河南农业大学培养室。待幼苗长出7~8片真叶(约40 d)后,选择健壮、长势一致的幼苗移栽到花盆中,每盆1株,盆内装土10 kg。每天浇水,使土壤含水量保持在田间持水量的80%左右。试验期间,每 10 d 调整花盆位置。试验共2个处理(CK和TR),每个处理设置12盆,共24盆。采集样品时,先用自来水冲洗去除泥土,再用去离子水冲洗后,85 ℃杀青30 min,于70 ℃烘干备用。
1.3测定指标及方法
1.3.1生长指标测定
分别于移栽后20、40、60、80 d时采集植株样品,测量株高和根长,分根、茎和叶3个部分,称干质量。叶绿素和类胡萝卜素采用95%乙醇浸提-比色法[13]测定。
1.3.2镉的测定
土壤总镉采用HF-HClO4-HNO3法消解,原子吸收分光光度法测定;土壤有效镉采用二乙基三胺五乙酸(DTPA)浸提剂提取,原子吸收分光光度法测定;植株镉采用HNO3-HClO4(体积比为3 ∶1)消化,原子吸收分光光度法[14]测定。
1.4数据分析
富集系数=根部镉含量/土壤中镉含量;转移系数=地上部镉含量/根部镉含量;镉累积量=器官镉含量×器官质量。采用SPSS 20.0进行数据统计分析;采用Excel 2007软件绘图。
2结果与分析
2.1镉对曼陀罗生长状况的影响
2.1.1株高和根长
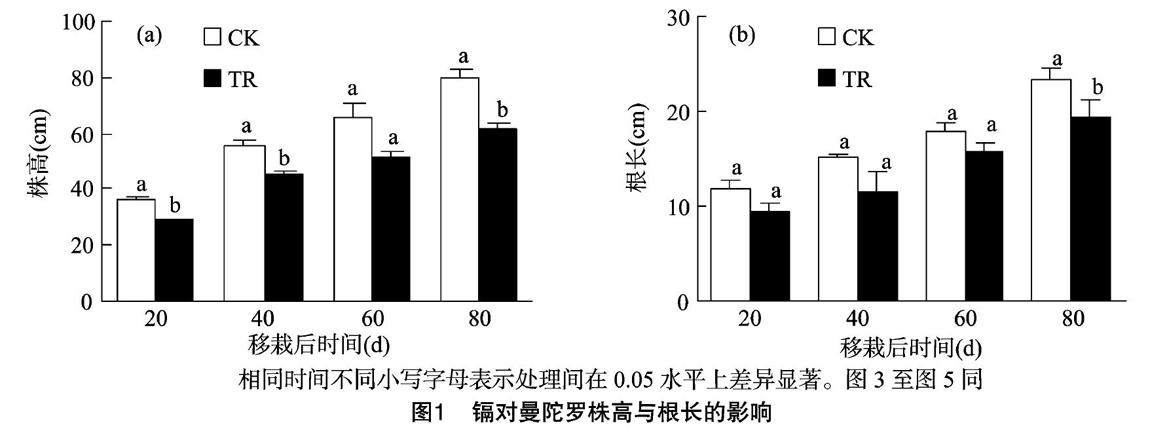
随移栽时间的增加,污染土壤组和对照(清洁土壤)组曼陀罗的株高和根长均有所增加,其中污染土壤组株高和根长最高可达61.9 cm和19.3 cm,相比对照减少22.6%和17.2%(图1)。曼陀罗移栽至污染土壤后,其株高明显低于对照组。就根长而言,移栽60 d之后,污染土壤组较对照组才开始受到抑制。
2.1.2生长指数
叶质量比和茎质量比分别是指叶和茎质量占植株总质量的比值,能够反映植株生物量的分配情况[15]。植株地上部生物量越大,其提取的重金属越多。由图2-a 可知,随时间增加,污染土壤组叶质量比呈现先减小后增加的趋势,在移栽60 d后,污染土壤组与对照组无明显差异。由图2-b可知,移栽20~60 d时污染土壤组曼陀罗的茎质量比高于对照组,随后趋于稳定。
2.1.3各器官干质量
随移栽时间的增加,污染土壤组与对照组的曼陀罗根、茎和叶干质量均增大(图3),但三者又有所不同。与对照相比,各采样时间点污染土壤组的根部干质量无明显减少(图3-a)。就地上部而言,整个生长期内,污染土壤组曼陀罗茎部干质量与对照无明显差异(图3-b);当移栽40 d以后,污染土壤组曼陀罗叶片干质量较对照显著减少(图3-c)。
2.2叶绿素和类胡萝卜素
由图4可知,当移栽60 d内,与对照组相比,污染土壤组的曼陀罗叶绿素a、b和类胡萝卜素含量无明显降低。只有当移栽时间增至80 d时,污染土壤组曼陀罗叶绿素a、b和类胡萝卜素较对照组显著降低,分别为1.10、0.38、0.21 mg/g。
2.3镉含量及富集、转移系数
由图5可知,随移栽时间增加,污染土壤組的曼陀罗根、茎、叶中镉含量均呈现减少的趋势。根、茎和叶镉含量分别达到26.09~37.96、26.67~42.93、24.54~47.02 mg/kg。在移栽后20 d时,曼陀罗根、茎和叶的镉含量最大,并且随时间增加,曼陀罗根部、叶片镉含量趋于稳定。相比于移栽20 d时,曼陀罗根部、茎部和叶片中镉含量最大降幅分别为31.3%、37.9%、47.8%。
富集系数反映植物吸收重金属能力[16],转移系数反映植物将重金属从地下部转移到地上部能力[17]。由表2可知,曼陀罗的根部富集系数均大于1,当移栽后20 d时,曼陀罗根部富[CM(25]集系数显著高于其他时间。随移栽时间增加,曼陀罗叶片转移系数变化不大,茎部转移系数先升高后降低,在移栽时间为40 d时,达到最大值1.54,显著高于其他时间。随移栽时间增加,地上部转移系数先升高后降低,最小值为0.94。
2.4各器官对镉的累积量
曼陀罗对镉的累积量反映其从土壤中吸取的镉总量。由图6可知,随时间增加,污染土壤组曼陀罗根部的镉累积量逐渐增加。曼陀罗根、茎和叶的镉累积量在移栽后最大可达 0.12、0.18、0.15 mg/株。在整个生长周期内,曼陀罗的地上部镉累积量所占百分比均在70%以上。
3讨论
目前已发现的许多超富集植物生长缓慢且适应能力差,这成为植物修复技术走向应用的障碍因素之一。曼陀罗作为一种野生植物,具有生长快、抗逆性强、分布广且生长周期短等特点[18],这些都有助于污染土壤的修复。本试验中,即使在重度污染土壤中,曼陀罗亦未发现明显的受害症状,表明曼陀罗对污染土壤的适应性强。虽然曼陀罗株高和根长受到一定抑制(图1),但就生物量而言,污染土壤组曼陀罗根部和茎部的干质量与对照组差异并不明显(图3),且地上部生物量(茎和叶的生物量之和)占总生物量比例也无明显差异,表明利用曼陀罗修复重度污染土壤具有一定的可行性。在移至污染土壤40 d时,茎质量比明显高于对照,进一步的生长调节和镉吸收调控若能针对这一时刻进行,则会提升修复效果。曼陀罗增加分配至茎部生物量的原因和机制目前尚不清楚,可能和污染条件下曼陀罗叶片的物质合成(叶绿素、类胡萝卜素等)及运输有关。为此,本研究分析了叶绿素、类胡萝卜素以初步判断曼陀罗叶片的生理状况。
随时间增加,污染土壤组与对照组的曼陀罗叶绿素a、b和类胡萝卜素均逐渐增多,表明这些物质在生物体内的合成并未受到阻止。叶绿素影响植物光合作用的强度及物质合成速率[19],一定程度反映了植物的适应能力;而类胡萝卜素影响植物细胞内活性氧的清除,与光合作用关系密切[20]。在移栽后的60 d内,污染土壤组与对照组曼陀罗的叶绿素a、b和类胡萝卜素无明显差异,由此分析,镉对曼陀罗生长并未产生显著影响。只有当胁迫时间增至80 d后,污染土壤组曼陀罗叶绿素a、b和类胡萝卜素含量才开始低于对照,这也侧面验证了曼陀罗对镉具有较强的耐受能力。
本试验中,曼陀罗地上部镉含量达到24.54~47.02 mg/kg,且在前3次采样时间点,地上部转移系数均大于1(表2),这表明曼陀罗具备相当的镉富集能力。就镉含量而言,曼陀罗相比于超富集植物有一定差距,但由于其具有较大的生物量,曼陀罗镉累积量与镉超富集植物东南景天相差不大。姚桂华等研究表明,镉浓度为13.15 mg/kg的酸性土壤中,东南景天单株镉累积量为0.53 mg[21]。魏树和等研究表明,当土壤中镉浓度为10 mg/kg时,镉超富集植物龙葵幼苗根、茎和叶镉含量分别达到29、62、76 mg/kg;高积累镉植物水蒿、蒲公英和刺儿菜地上部镉含量分别达到21、33、16 mg/kg。相比于龙葵,曼陀罗根部镉含量与之相差不大,但茎部和叶片的镉含量较低,并未达到镉超富集植物的标准[22-23]。李念等采用大田栽培模式,发现曼陀罗单株镉提取量远小于本试验结果[24]。这可能是由于本试验采用的土壤镉浓度较大,以及栽培管理模式不同造成的。但是,鉴于曼陀罗具有较高的镉含量、较强的根部富集及地上部转移等优点,初步认为曼陀罗具有镉高累积的特性。
本研究设置了移栽后20、40、60、80 d 4个采样时间,以全面了解曼陀罗对镉的吸收、转移和累积特征。曼陀罗的镉累积量在整个生长周期中并不是持续增加,表现出独特的镉累积特性,这一特性主要受地上部镉浓度、生物量累积、转移系数等影响。累积量在移栽后60 d后达到最大,此时地上部镉累积量占植株累积量的70%以上,当胁迫时间继续增加至 80 d 后,累积量不再增加。
4结论
在选取的重度污染土壤上,曼陀罗对镉具有较强的富集性,镉含量最高可达47 mg/kg。从修复角度而言,地上部具有较高的镉累积能力,其累积量占植株累积量的比例超过70%。镉累积量随时间有所增加,达到峰值(本试验为移栽60 d)后趋于稳定,曼陀罗植物提取修复应选择最佳的时间点收割地上部,尤其是茎部。
参考文献:
[1]环境保护部,国土资源部. 全国土壤污染状况调查公报[R/OL]. (2014-04-17)[2018-11-11]. http://www.gov.cn/foot/site1/20140417/782bcb88840814ba158d01.pdf.
[2]Hua M,Zhang S J,Pan B C,et al. Heavy metal removal from water/wastewater by nanosized metal oxides:a review[J]. Journal of Hazardous Materials,2012,211/212:317-331.
[3]van der Ent A,Baker A J M,Reeves R D,et al. Hyperaccumulators of metal and metalloid trace elements:facts and fiction[J]. Plant and Soil,2013,362(1/2):319-334.
[4]Mahar A,Wang P,Ali A,et al. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils:a review[J]. Ecotoxicology and Environmental Safety,2016,126:111-121.[HJ1.72mm]
[5]Zhang X H,Liu J,Huang H T,et al. Chromium accumulation by the hyperaccumulator plant Leersia hexandra Swartz[J]. Chemosphere,2007,67(6):1138-1143.
[6]魏樹和,周启星,王新,等. 一种新发现的镉超积累植物龙葵(Solanum nigrum L.)[J]. 科学通报,2004,49(24):2568-2573.
[7]Ma L Q,Komar K M,Tu C,et al. A fern that hyperaccumulates arsenic[J]. Nature,2001,409(6820):579.
[8]中国科学院中国植物志编辑委员会. 中国植物志:第67卷[M]. 北京:科学出版社,1978.
[9]Varun M,DSouza R,Pratas J,et al. Metal contamination of soils and plants associated with the glass industry in North Central India:prospects of phytoremediation[J]. Environmental Science and Pollution Research,2012,19(1):269-281.
[10]杨海涛,卢炳坤,苗利娟,等. 野生曼陀罗生态学特性及镉铅携带量的研究[J]. 西南农业学报,2016,29(2):413-419.
[11]董林林,赵先贵,韦良焕. 曼陀罗和苍耳对污染土壤中镉的吸收与富集[J]. 生物技术,2009,19(2):29-32.
[12]史雷,慕小倩. 曼陀罗种子破眠方法研究[J]. 种子,2010,29(9):40-43.
[13]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[14]鲍士旦. 土壤农化分析[M]. 3版. 北京:中国农业出版社,2008.
[15]林波,刘庆. 四种亚高山针叶林树种的表型可塑性对不同光照度的响应[J]. 生态学报,2007,28(10):4665-4675.
[16]张红振,骆永明,章海波,等. 水稻、小麦籽粒砷、镉、铅富集系数分布特征及规律[J]. 环境科学,2010,31(2):488-495.
[17]李霞,张银龙,魏猷刚,等. 芦蒿对Cd积累与转运的基因型差异研究[J]. 农业环境科学学报,2013,32(11):2132-2139.
[18]杨珍平,郝教敏,卜玉山,等. Cd胁迫对5种植物体内Cd积累及根际土壤特性的影响[J]. 水土保持学报,2011,25(6):188-190.
[19]宇克莉,孟庆敏,邹金华. 镉对玉米幼苗生长、叶绿素含量及细胞超微结构的影响[J]. 华北农学报,2010,25(3):118-123.
[20]陈良,隆小华,郑晓涛,等. 镉胁迫下2种菊芋幼苗的光合作用特征及镉吸收转运差异的研究[J]. 草业学报,2011,20(6):60-67.
[21]姚桂华,徐海舟,朱林刚,等. 不同有机物料对东南景天修复重金属污染土壤效率的影响[J]. 环境科学,2015,36(11):4268-4276.
[22]魏树和,周启星,王新. 超积累植物龙葵及其对镉的富集特征[J]. 环境科学,2005,26(3):167-171.
[23]魏树和,周启星,王新,等. 杂草中具重金属超积累特征植物的筛选[J]. 自然科学进展,2003,13(12):1259-1263.
[24]李念,李荣华,冯静,等. 粉煤灰改良重金属污染农田的修复效果植物甄别[J]. 农业工程学报,2015,31(16):213-219.
