海拔和环境因子对濒危植物青皮在海南岛中部丘陵区人工林下生长和光合特性的影响
2019-08-20王旭萍王亚陈刘强王瑾冯剑阮长林
王旭萍 王亚陈 刘强 王瑾 冯剑 阮长林


摘要:为了分析乡土树种青皮在海南岛中部丘陵地区人工林向自然林生态恢复中的应用情况,在270、380 m 2个不同海拔样地的人工林中种植青皮苗木,研究环境条件对青皮生长和光合作用特性的影响。结果表明,青皮的成活率经过初期的下降,在定植1年5个月之后基本稳定。海拔为380 m样地青皮的成活率在各个时间段均高于海拔为 270 m 样地的青皮,人为干扰的影响较大。2个海拔上的青皮苗木的株高和基径的生长没有明显差异。2个样地青皮苗木的株高、基径的净增长量随着时间的推移呈增大趋势,且海拔为270 m样地青皮的株高增长量与土壤含水量呈显著正相关,其株高增长量、基径增长量均与土壤全磷含量呈显著负相关;海拔为380 m样地青皮的株高增长量、基径增长量与土壤全钾含量呈显著或极显著负相关,2个样地的土壤全磷、土壤全钾含量逐年降低。海拔为270 m样地青皮的净光合速率明显大于海拔为380 m的样地。2个样地青皮的净光合速率与光合有效辐射、CO2浓度、空气相对湿度等呈极显著正相关,与温度呈极显著负相关,水分利用效率与光合有效辐射、温度、空气相对湿度等呈极显著正相关。2个样地青皮的净光合速率和水分利用效率随时间的推移而明显提高。由研究结果可知,青皮可在丘陵区不同的海拔地区存活并良好地生长。研究结果可为海南岛中部丘陵区青皮种群的人工恢复种植提供参考。
关键词:濒危物种;青皮;生长;光合作用;海拔
中图分类号: X171.4文献标志码: A
文章编号:1002-1302(2019)08-0148-06
青皮(Vatica mangachapoi)属东亚热带雨林特征科龙脑香科(Dipterocarpaceae)植物,是海南岛自然分布的3种龙脑香科植物之一,在海南岛曾经广泛分布[1]。但是目前青皮已经成为国家二级保护植物[2],在海南省主要分布在由中部山区分隔的自然保护区中,在沿海个别地区如万宁石梅湾有分布,在山区与沿海台地之间的丘陵区分布不多。人为砍伐破坏、生境破碎化以及青皮幼苗受光照环境和种间竞争的抑制是导致青皮濒危的三大主要原因[3-5]。目前,关于青皮林资源、种群和群落结构、分布规律及其生长过程的报道较多[2,6],海南岛大部分青皮分布在海拔300~650 m之间[2,6-7]。郝清玉等对不同海拔的青皮林进行抽样调查,结果发现青皮林的群落结构、多样性及天然更新程度与海拔有一定的相关性[8-9]。目前,针对青皮生长特性的研究大多仅集中于在不同人为控制的环境条件下(如不同土壤条件、不同光照条件、不同浓度的木麻黄浸提液等)对青皮光合特性的影响方面[10-13]。为了扩大青皮种群,本研究将青皮苗木混交种植在海南中部丘陵区不同海拔的人工林中,研究其生长规律,以期使用青皮促进人工林向自然植被演替,从而實现热带雨林的生态恢复。
1材料与方法
1.1研究地概况
本研究样地位于海南省琼中县湾岭镇罗马村,属于热带海洋气候区北缘,具有昼热夜凉的山区气候特征。年平均气温为22.8 ℃,年平均降水量为2 400 mm左右,有明显的干湿季。干季为12月至翌年5月,湿季为6—12月,9、10月为降雨高峰期。一年中的6、7月气温最高,1月最低。试验样地分别位于湾岭罗马村海拔为270、380 m的2个人工林中,海拔为270 m的样地(109°38′42″ E,19°16′29″N)植被主要为人工种植的菠萝蜜(Artocarpus heterophyllus)林,海拔为380 m的样地(109°38′78″ E,19°16′18″N)植被主要为人工种植的马占相思(Acacia mangium)林。
1.2试验材料
青皮苗木为一年生种子苗木,取自海南省尖峰岭苗圃,平均株高为11.7 cm,于2009年1月定植于上述2个人工林样地内。按行距80 cm、株距60 cm种植,每个样地种植的青皮数量为100株。从定植的2009年1月至2012年12月的4年间,每半年观测1次,2013—2015年间每年观测1次。
1.3试验方法
1.3.1青皮生长状况的测定(1)存活率。存活率=存活的青皮苗木数量/定植时的青皮苗木总数量(100株)×100%。(2)株高、基径。分别用钢卷尺与游标卡尺测定苗木的株高、基径。(3)光合作用特性等指标。选择晴好的天气,使用Li-6400便携式全自动光合测定仪测定,测定的光合因子有净光合速率[Pn,μmol/(m2·s)]、气孔导度[Gs,mol/(m2·s)]、蒸腾速率[Tr,mmol/(m2·s)]、胞间CO2浓度(Ci,μmol/mol),08:00—18:00每隔2 h测定1次;水分利用效率=净光合速率/蒸腾速率[14]。
1.3.2土壤因子的测定在样地内按“S”形线路选取5个点,分别测定各个点的土壤环境因子及土壤养分含量。现场测定的土壤因子指标有土壤pH值(用IQ150土壤pH计测定)、土壤水分含量、盐分含量、温度(用Hydra土壤水分-盐分-温度测定仪测定);测定的土壤养分指标有土壤全氮含量(用凯氏定氮法测定)、土壤全磷含量(用钼锑抗比色法测定)、土壤全钾含量(用NaOH熔融-火焰光度法测定)[14]。
1.3.3气候因子的测定测定的气候因子指标有光合有效辐射[PAR,μmol/(m2·s)]、空气CO2浓度(Ca,μmol/mol)、空气温度(Ta,℃)、空气相对湿度(RH,%)。测定使用的仪器为Li-6400便携式全自动光合测定仪。
2结果与分析
2.1青皮生长状况的比较
2.1.12个样地青皮存活率的比较如图1所示,在2009年6月至2010年6月的种植初期,青皮的成活率在定植后5个月至1年5个月的时间内大幅下降;2010年6—12月的超强降水引起的洪涝灾害可能是造成2个样地的存活率都急剧下降的原因,另外也与青皮苗木适应环境需要一定的时间有关;至2010年12月定植约2.0年后,青皮的存活率逐渐趋于平稳,但海拔为380 m样地的青皮成活率明显高于海拔为270 m的样地,可能与低海拔样地靠近村庄,人为活动频繁,干扰强度较大有关,因此笔者多次到样地做观测研究时发现,低海拔样地有附近村民的牛在其中活动,它们会踩踏、啃食植物。
2.1.22个样地青皮的株高和基径生长状况由图2-A可以看出,2个样地青皮的株高、基径都随着时间的推移而不断增长,但是2个样地之间在各个时间点上的差异均不明显。由图2-B可以看出,2个样地青皮株高、基径的净增长量有随时间的推移而不断加大的趋势。在定植初期的2009年6月至2010年6月期间,株高、基径的净增长量均较小,而2010年6月至2014年12月,株高的净增长量明显增大,呈现加速生长的趋势,但是2个样地之间在各个时间点上的差异均不明显。
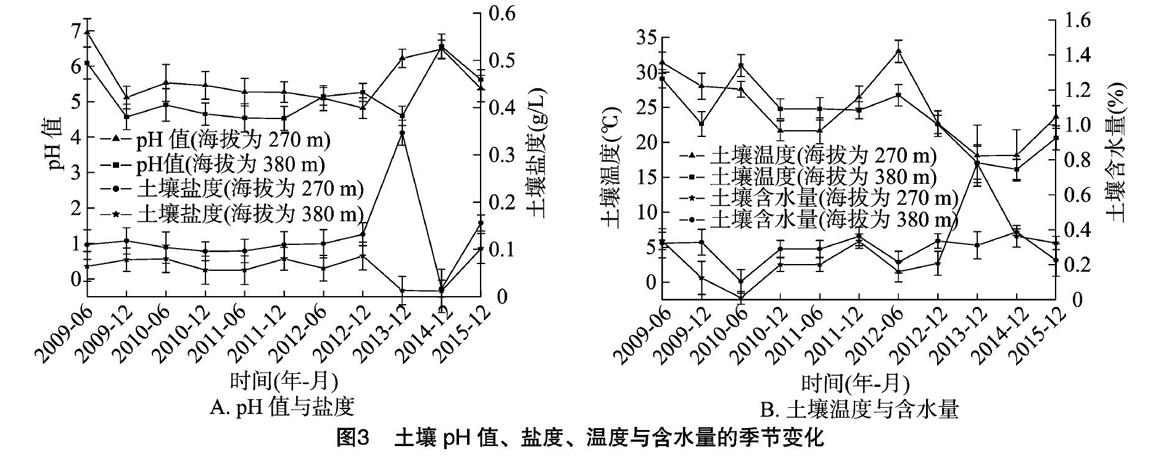
2.1.3土壤環境因子的季节变化动态由图3-A可知,海拔为270 m与海拔为380 m样地的土壤pH值均在4.47~6.94之间波动,且差异不明显。海拔为270 m样地的土壤盐度在2012年12月至2014年12月间出现剧烈的“上升—下降”波动,380 m 海拔样地土壤盐度的下降,可能与台风带来的巨量短时降雨的影响有关。因为高海拔样地位于一个丘坡的顶部,其土壤受到雨水的强烈冲洗后,会使盐度降低,而低海拔样地处于丘坡的下部,地势相对平缓,丘坡上部淋溶下来的盐分可能在这里积累,导致盐分含量上升,而2个样地的土壤盐度差异不明显。
由图3-B可以看出,2个样地的土壤温度、含水量变化趋势具有季节性。海拔为270 m样地的平均土壤温度大于海拔为380 m的样地,且土壤温度总体上有下降趋势,可能由于林地的进一步发育,盖度增大,林下光照度下降。海拔为 380 m 样地的土壤含水量平均值略大于海拔为270 m的样地,海拔为270 m的样地的土壤含水量在2012年12月至2014年12月间出现的剧烈“上升—下降”波动可能与台风带来的强降雨有关,使得海拔较低的地区雨水聚集,但是2个样地间的土壤温度、含水量差异均不明显。
2.1.4土壤养分的季节变化动态如图4所示,随着时间的推移,2个样地土壤全钾、全磷含量整体呈降低的趋势,但是2个样地土壤全氮含量都比较稳定。海拔为270 m样地的土壤全钾含量明显大于海拔为380 m的样地,全磷、全氮含量差异不明显。
在季节动态维度上对青皮生长状况与环境因子进行相关性分析,如表1所示,海拔为270 m样地青皮的株高增长量与土壤含水量呈显著正相关,与土壤全磷含量呈显著负相关;海拔为380 m样地青皮的株高增长量和基径增长量与土壤全钾含量呈显著或极显著负相关。水分对青皮的生长至关重要,土壤全磷、全钾含量呈下降趋势,一方面与树木生长消耗了土壤养分有关,另一方面与地形、降雨量有关。土壤全氮含量相对稳定,可能与高海拔样地的马占相思(为豆科植物)可以固氮有关。青皮等树木的生长是以消耗土壤养分为代价的,随着样地树木的生长,土壤养分被大量消耗却得不到足量的补给,就会导致如图4所示土壤全磷、全钾含量连年降低的现象,进而使得青皮的生长与土壤养分含量呈负相关。
2.2青皮光合作用的变化动态
2.2.1气候因子的日变化动态如图5-A所示,2个海拔样地的光合有效辐射、空气温度日变化均呈单峰型,在t1~t3这3个时间段内呈上升趋势,在t3时达到最大值,之后呈下降趋势;海拔为270 m样地的光合有效辐射、气温整体上均明显高于海拔为 380 m 的样地,这可能与人工林的植被状况有关,海拔为270 m的样地内主要是一些灌木和较大的菠萝蜜树,光照度日均值为68~129 μmol/(m2·s),郁闭度低于海拔为380 m的成片马占相思林,后者的光照度日均值为40~68 μmol/(m2·s)。
由图5-B可以看出,2个海拔样地空气CO2浓度、空气相对湿度的日变化趋势与光合有效辐射、 气温的日变化趋势相反,均在t3时间段内达到最小值,且海拔为380 m样地的空气CO2浓度与空气相对湿度均大于海拔为270 m的样地,但差异都不明显。表明在郁闭度较高的林地,其对水分的保持力较强。
2.2.2青皮光合作用的日变化动态植物的光合特性受气候、立地条件和本身生理生态特性等多个因素的影响,随着这些因素的变化而呈现出一定的日变化规律[15]。由图6-A、图6-B 可知,2个海拔样地青皮苗的净光合速率、气孔导度、蒸腾速率的变化曲线均呈单峰型。分析表明,海拔为 270 m 样地的青皮苗胞间CO2浓度、气孔导度、蒸腾速率整体上均大于海拔为380 m的样地,但差异都不明显。青皮胞间CO2浓度的日变化趋势与净光合速率、气孔导度、蒸腾速率的日变化趋势相反,在t3时间段达到最小值;分析表明,海拔为 270 m 样地青皮的净光合速率明显大于海拔为380 m的样地。由图6-C可知,2个海拔样地青皮的水分利用效率的变化曲线均呈单峰型,海拔为380 m样地青皮苗的水分利用效率大于海拔为270 m的样地,但差异不明显。
由表2可以看出,2个样地青皮的净光合速率与光合有效辐射、空气CO2浓度、空气相对湿度呈极显著正相关,与温度呈极显著负相关;气孔导度与空气相对湿度呈极显著正相关,与温度呈极显著负相关;胞间CO2浓度与空气CO2浓度、空气相对湿度呈极显著正相关,与光合有效辐射呈极显著负相关;蒸腾速率与光合有效辐射、温度、CO2浓度、空气相对湿度呈极显著正相关;水分利用效率与光合有效辐射、温度、空气相对湿度呈极显著正相关。海拔为270 m样地的青皮胞间CO2浓度与温度呈极显著正相关;在温度升高至导致植物进行午休前,其净光合速率随着温度的升高而升高,此时,叶片的气孔打开,吸收大量CO2来进行光合作用,即出现正相关的现象;海拔为270 m样地青皮的水分利用率与空气CO2浓度呈极显著正相关,水分利用效率反映了植物的光合能力,CO2是植物光合作用的原料,海拔为270 m样地的CO2浓度小于海拔为380 m的样地,由于植物的光合作用更需要CO2的供给,出现这种现象,可能是由海拔为270 m的样地CO2供不应求造成的。
2.2.3青皮光合作用的季节变化动态如图7-A、图7-B所示,2个样地青皮苗的净光合速率季节变化趋势基本相同,总体上均呈增长趋势,反映了随着青皮苗木的生长,其净光合速率也随之增长;青皮气孔导度、蒸腾速率的季节变化趋势与净光合速率的变化趋势基本相同;蒸腾速率整体呈波动上升趋势,2个样地青皮的胞间CO2浓度总体上保持平稳状态。从图7-C可以看出,2个海拔样地青皮苗木的水分利用效率总体上随着青皮苗的生长呈增大的趋势,说明随着青皮苗的生长,其对水分的利用效率在逐渐增大。
3结论与讨论
3.1青皮的生长状况
在本研究中,青皮苗的存活率在初期下降得较快,翁文霞等将青皮移植到滨海木麻黄林中进行生长研究,发现青皮在生长初期的苗木死亡数量较多,在将近2年的时间里,存活率下降了50%左右[10],与本研究中青皮移植到林地中后生长初期的情况相似,这可能与青皮苗木对新的生境需要一定的适应期有关。此外,海拔为270 m样地青皮的存活率在不同阶段均低于海拔为380 m的样地,这可能与低海拔地区人类频繁活动有关。在进行观测研究时,笔者发现样地时常有人、畜等进入,对青皮生长等造成了严重干扰,这也是青皮存活率下降的原因之一。2个样地青皮的株高、基径与土壤全磷、全钾含量呈负相关,且2个样地的土壤全磷、全钾含量出现连年降低的现象,表明随着2个样地中青皮及人工林植物的生长,土壤养分被大量消耗却得不到足量的补给。青皮和周围其他植物的生长是以消耗土壤养分为代价的, 土壤养分的供不应求会导致土壤养分的含量下降,在数年的较长观察期中,呈现出植物生长和土壤养分呈负相关的情形。土壤养分的补给能力可以从“生”和“储”2个方面进行评价:2个样地的磷、钾均主要来自植物凋落物降解以及土壤母质的分解释放,海拔为270 m的样地主要为灌木和几株菠萝蜜树,海拔为380 m的样地主要为成熟的马占相思林,钾的补给能力低可能是由于雨水的淋溶,使得380 m海拔样地钾的流失量较大造成的。2个样地中土壤全氮含量相对稳定,这与高海拔样地种植的可以固氮的豆科植物马占相思有关,通过雨水等的淋溶,可以对低海拔样地的土壤氮含量进行补充。
3.2青皮的光合特性
2个样地青皮苗的水分利用效率与空气温度、空气相对湿度呈正相关,其中海拔为270 m的样地与空气相对湿度呈极显著正相关,表明空气相对湿度是青皮对水分利用的显著影响因子,在较高的空气相对湿度下,青皮苗的光合速率更高,对水分的利用效率也就更高,且在2个样地中,随着青皮的生长,对水分的利用效率整体呈现出逐渐增大的趋势。水分利用效率通常用来作为评价植物适应性的指标,随着青皮林龄的增长,青皮的净光合速率呈逐渐增长的趋势,导致其对水分利用效率的提高。气孔导度直接影响光合速率,而随着青皮林龄的增长,其气孔导度呈现缓慢增长的趋势,与净光合速率的变化一致。袁颖红等在研究不同林龄人工林尾巨桉的光合特性及水分利用效率时发现,尾巨桉随林龄的增大,其净光合作用减弱,导致其水分利用效率降低[16],本研究结论与之相似。
2个不同海拔样地青皮的光合特征日变化趋势基本相似,是环境因子和青皮苗生理生态因子综合作用的结果,与王亚陈等对木麻黄林下青皮苗光合特性对光照度的响应的研究结果[12]相似。青皮胞间CO2浓度日变化趋势呈“V”形,这与杨雨华等对不同生长势马尾松各无性系胞间CO2浓度日变化规律研究结果一致[13]。海拔为270 m样地青皮的胞间CO2浓度、气孔导度、蒸腾速率大于海拔为380 m的样地,净光合速率明显大于海拔为380 m的样地,海拔为270 m样地青皮水分利用效率的平均值小于海拔为380 m的样地,2个海拔样地光合特性存在明显差异与2个样地的环境条件不同[14-15,17-18]有关。相关性研究表明,青皮净光合速率、蒸騰速率与光合有效辐射呈显著正相关,说明较强的光照有利于青皮的生长,马兴宇等探究了环境因子对鼠尾藻生殖托生长及光合特性的影响,结果表明,光照度、温度对鼠尾藻生殖托的生长及光合特性均有显著影响[19],与本研究结果相似,也与Shu等研究得出的光照度对水葫芦生长及生理特性影响的结果一致[20]。
海南岛全岛的地形为中高周低,在中部的山区与沿海台地之间有大面积的丘陵区,丘陵区的林地多为人工林。可见在丘陵区将人工林恢复为青皮林,也对改善海南岛森林生态环境具有积极意义。本研究中,在不同海拔人工林下的青皮苗木的株高和基茎的生长没有显著差异,2个样地的青皮长势较好,但是青皮苗木的存活率受人为干扰较严重。此外,不同海拔的环境因子差别对青皮的净光合速率造成了较大的影响。如果延长观测时间,净光合速率的差异可能最终导致青皮植株在生长上的差异。因此,在海南岛中部丘陵区将青皮苗移栽到不同海拔的林地中进行混交种植,通过这种方式扩大和恢复青皮的种群是完全可能的,但需要注意排除人为干扰,使其自然生长恢复,可以取得更好的效果。
参考文献:
[1]陈铭枢. 海南岛志[M]. 海口:海南出版社,2004:347-355.
[2]许涵,李意德,骆土寿,等. 海南岛国家重点保护植物青皮(Vatica mangachapoi)研究综述[J]. 热带林业,2007,35(2):8-11.
[3]杨永康,吴家坤,龚祝南. 中国龙脑香科植物分类研究(三)[J]. 中国野生植物资源,2002,21(5):1-7.
[4]杨清,肖来云,普正和,等. 渐危树种青梅的迁地保护研究[J]. 广西植物,1996,16(1):64-68.
[5]孟令曾,许再富. 中国龙脑香科植物受胁状况及迁地群落保护探讨[J]. 广西植物,2005,25(1):8-13.
[6]李希娟,宋启道,陈秋波. 海南霸王岭林区青皮天然林资源与乔木层群落学特征[J]. 林业资源管理,2008(2):85-89,94.
[7]方洪,李意德,罗文,等. 尖峰岭国家级自然保护区青皮林资源及其垂直分布特征[J]. 热带林业,2004,32(4):43-46,38.
[8]郝清玉,刘淑菊,钟琼芯,等. 海南岛不同海拔高度青皮林群落结构与多样性[J]. 林业资源管理,2012(4):48-54.
[9]郝清玉,王贵,吕冰,等. 海南岛不同海拔高度青皮林天然更新特征[J]. 林业资源管理,2012(5):39-45,58.
[10]翁文霞,许素云,刘强,等. 青皮苗木在不同土壤中光合作用日变化的初步研究[J]. 海南师范大学学报(自然科学版),2009,22(4):441-445.
[11]伍恩华,李燕华,王敏英,等. 青皮苗木在不同光照处理下的光合作用日变化特征[J]. 海南师范大学学报(自然科学版),2009,22(2):185-190,203.
[12]王亚陈,翁文霞,刘强. 木麻黄林下青皮苗光合作用特性对光照度的响应[J]. 湖北农业科学,2012,51(8):1615-1621.
[13]杨雨华,宗建伟,杨风岭. 不同生长势马尾松光合日变化研究[J]. 中南林业科技大学学报,2014,34(8):25-29.[HJ1.72mm]
[14]Cousins A B,Johnson M,Leakey A B. Photosynthesis and the environment[J]. Photosynthesis Research,2014,119(1/2):1-2.
[15]Zhu J J,Wang K,Sun Y R,et al. Response of Pinus koraiensis seedling growth to different light conditions based on the assessment of photosynthesis in current and one-year-old needles[J]. Journal of Forestry Research,2014,25(1):53-62.
[16]袁颖红,樊后保,吴建平,等. 不同年龄人工林尾巨桉(Eucalyptus urophylla×E. grandis)叶片光合特性及水分利用效率[J]. 应用与环境生物学报,2016,22(1):58-63.
[17]Maberly S C. The fitness of the environments of air and water for photosynthesis,growth,reproduction and dispersal of photoautotrophs:an evolutionary and biogeochemical perspective[J]. Aquatic Botany,2014,118(SI):4-13.
[18]Yamori W,Hikosaka K,Way D A. Temperature response of photosynthesis in C3,C4,and CAM plants:temperature acclimation and temperature adaptation[J]. Photosynthesis Research,2014,119(1/2):101-117.
[19]馬兴宇,梁洲瑞,刘福利,等. 环境因子对鼠尾藻生殖托生长及光合特性的影响[J]. 中国水产科学,2013,20(4):851-858.
[20]Shu X,Zhang Q F,Wang W B. Effects of temperature and light intensity on growth and physiology in purple root water hyacinth and common water hyacinth (Eichhornia crassipes)[J]. Environmental Science and Pollution Research,2014,21(22):12979-12988.
