不同基因型烤烟对镉、铅富集特征评价
2019-08-20悦飞雪李继伟王艳芳刘杉王小东陈明灿刘领
悦飞雪 李继伟 王艳芳 刘杉 王小东 陈明灿 刘领


摘要:重金属镉、铅易在烟叶中富集,影响烟株生长,并通過抽吸过程进入人体,危害人类健康。为探讨豫西地区不同基因型烟草对重金属镉、铅的富集特征,筛选出烟叶原料安全的优良品种,以豫西烟区15个基因型烤烟为材料,采用水培试验研究不同水平镉(0、5、10、20 mg/L)、铅(0、20、50、100 mg/L)胁迫下不同基因型烤烟生物量、重金属含量、耐性指数、重金属积累量及转运系数的差异。结果表明:(1)重金属镉、铅胁迫对不同基因型烟株的生长均表现为低浓度促进,高浓度抑制的现象。(2)不同基因型烤烟对重金属镉、铅污染的响应不同。不同基因型烤烟对镉的积累分配主要在地上部,而对铅的积累分配主要在根部,且对重金属镉的富集和转运能力高于重金属铅。(3)不同烤烟品种对重金属镉、铅的吸收积累能力存在基因型差异。在高镉、铅水平下G15对重金属镉的积累量和转运系数较高,G5对重金属铅的积累量、耐性指数较高,适合作为土壤重金属镉、铅污染的植物修复材料。G2、G10在低镉水平下,镉的积累量较低,适宜在土壤轻度镉污染条件下进行烟叶安全生产。G4、G12在低铅水平下,铅积累量和地上部分铅含量较低,属于铅低积累基因型,可用于轻度铅污染条件下的烟叶安全生产。
关键词:烤烟基因型;镉和铅;耐性指数;吸收积累;土壤污染修复;植物材料
中图分类号: S572.01;X53文献标志码: A
文章编号:1002-1302(2019)08-0105-07
土壤重金属污染会导致农作物生长缓慢,产量和品质下降[1-2]。镉、铅是土壤重金属污染的重要元素,也是对动植物毒性较大的2种元素[3-4]。土壤中的重金属镉、铅含量一旦过量,就很难将其从土壤中去除,导致不能“自主移动”的植物不可避免地会受到镉、铅污染的毒害,进而通过食物链富集给人类健康带来巨大的危害[5]。因此,如何修复和利用不同程度重金属污染的土壤是人们面临的重要课题,也是十分迫切需要解决的重要环境问题。
植物修复技术由于具有运作成本低、操作简便、非扰动土壤结构、不造成二次污染等特点,而被视为一项绿色、环境友好和廉价的污染治理技术,主要包括植物提取和植物稳定2种修复途径[6]。植物提取技术是指利用重金属超富集植物把土壤中的重金属移至植物体内,减少土壤中的重金属积累,从根本上修复污染土壤[7]。然而,超富集植物往往因其生长慢、生物量低,在短期内的修复效果不佳,应用前景受限[8]。生物量高、重金属富集能力中等的植物具有较强的生态适应能力,在修复重金属污染土壤方面具有很大的潜力。因此,生物量高、重金属富集能力较强的植物材料具有潜在的应用前景[9]。植物稳定技术是指通过植物根系分泌物降低土壤中重金属的活性,减少作物对重金属的吸收,或通过种植重金属低积累的植物实现清洁生产和食物安全[10]。在重金属污染的土壤上低积累植物作为产品的安全生产模式日益受到重视,通过挖掘作物自身的遗传潜力,选择对污染物低吸收、低积累的作物品种是低污染条件下建立作物产品安全生产体系的重要途径[11-12]。
植物对元素过量、缺素或者其他逆境胁迫的耐性存在广泛的物种间及基因型间差异[13]。已有研究表明,不同类型及基因型的植物对土壤中重金属的吸收积累能力有很大差别,如Chernobrovkina等研究发现,红三叶草、蒲公英和梯牧草地上部对土壤中Zn、Pb、Cu、Ni的积累具有较大差异,且对4种重金属的积累能力大小均表现为红三叶草>蒲公英>梯牧草[14]。李月芳等报道了不同基因型玉米对重金属铅富集能力的差异,结果表明,掖单13号具有较强的铅转运能力,申甜1号的生物富集能力最强,掖单13号和申甜1号均具有较强的铅吸收和积累能力,属于潜在的高积累基因型;金珠蜜超甜和申甜1号2个甜玉米基因型果穗内铅积累量较少,其含量符合国家规定的食品生产相应的安全标准[15]。另外,一些研究报道了其他作物对镉、铅积累的基因型差异,例如,水稻、小麦、大麦、大豆、花生、马铃薯、向日葵等[16-17]。
烟草是我国重要的经济作物之一,优质烟叶原料是烟草行业赖以生存和发展的物质基础,也是烟区可持续发展的重要保障[18-19]。重金属是卷烟烟气中的有害成分之一,在抽吸过程中卷烟中的微量重金属随主流烟气进入人体,并在人体中富集[20]。吸烟作为人体重金属富集的一个途径一直是卷烟降焦减害的研究热点之一。源头控制是减少烟草重金属危害的主要途径。豫西烟区是我国著名的优质烟叶生产基地之一,也是我国重要的重金属矿集区,近年来由于过度的矿产资源开发,该地区镉、铅污染问题较为严重,已经成为限制烟区发展的重要因子[21-22]。尽管近年来围绕不同基因型烟草对重金属的富集特征开展了一些研究,但多数研究中的参试基因型数量较少,且关于重金属镉元素的研究较多,而对重金属铅元素的研究较少,目前对豫西烟区重金属烟草品种的筛选研究鲜有报道。为此,本研究针对豫西烟区烟草的品种特色,分析不同基因型烤烟对重金属镉、铅的富集特征,目的是一方面筛选出低积累的烤烟品种,为当地烟叶安全生产提供服务;另一方面筛选出对重金属富集能力较强的烤烟品种,为植物修复土壤重金属污染提供材料。
1材料与方法
1.1供试材料
以河南省洛阳市烟草公司提供的15份不同基因型烤烟(表1)为试验材料,包括豫西烟区的主栽品种及部分引进品种,来源比较广泛且品种间存在较大遗传差异。供试的15个基因型烤烟由洛阳市烟草公司统一育苗,待烟苗长至5叶期时,备用。供试浮盘由聚苯乙烯泡沫塑料制成。供试重金属模拟污染试剂为硫酸镉(CdSO4)和醋酸铅[Pb(CH3COO)2]。
[BH]G15龙江981黑龙江引品种[BG)F][FK)]
1.2试验设计
试验设置0、5、10、20 mg/L等4个镉胁迫水平,分别记为Cd0、Cd5、Cd10、Cd20处理;设置0、20、50、100 mg/L等4个铅胁迫水平,分别记为Pb0、Pb20、Pb50、Pb100处理。镉、铅胁迫水平分别由纯品硫酸镉、醋酸铅配制而成,每个处理重复3次,完全随机排列。
2.3不同基因型烤烟镉、铅耐性指数
耐性指数越大说明该品种对重金属的耐性越强。从图 3-a可以看出,不同基因型烤烟的耐性指数随着镉胁迫水平的升高整体呈现出降低趋势。不同基因型烟株对镉胁迫的响应不同,在5 mg/L镉浓度胁迫下,G12、G14的镉耐性指数显著高于其他基因型烟株(P<0.05);在10 mg/L镉浓度胁迫下,G3的镉耐性指数显著高于其他基因型烟株(P<0.05),G2、G9、G10、G13的镉耐性指数显著低于其他基因型烟株(P<0.05);在20 mg/L镉浓度胁迫下,G3、G5、G14的镉耐性指数较高,G2、G8的镉耐性指数显著低于其他基因型烟株(P<0.05)。
从图3-b可以看出,随着铅胁迫浓度的升高,不同基因型烟草的耐性指数整体呈现降低的趋势。不同基因型烟株对铅胁迫的响应不同,在3个铅胁迫水平下,G5的铅耐性指数均较高。在50 mg/L铅浓度下,G5的铅耐性指数显著高于其他基因型烟株(P<0.05),G1、G2的铅耐性指数显著低于其他基因型烟株(P<0.05);在100 mg/L铅浓度胁迫下,G5的铅耐性指数较高,G6的铅耐性指数最低,且显著低于其他基因型烟株(P<0.05)。
2.4不同基因型烤烟镉、铅积累量
从图4-a可以看出,在未添加镉的处理中,不同基因型烟草镉积累量很小。对不同基因型烤烟的镉积累量而言,在 5 mg/L 镉浓度胁迫下,G3、G13、G15的镉积累量较高,其中G15的镉积累量显著高于其他基因型烟株(P<0.05)。在 10 mg/L 镉浓度胁迫下,G3、G15的镉积累量较高,且显著高于其他基因型烟株(P<0.05);G2、G10的镉积累量较低,显著低于其他基因型烟株(P<0.05)。在20 mg/L镉浓度胁迫下,G15的镉积累量最高,显著高于其他基因型烟株(P<0.05),G1、G2、G4、G10的镉积累量较低,显著低于其他基因型烟株(P<0.05)。
由图4-b可知,随着铅胁迫浓度的增加,烟草对重金属铅的积累量整体呈现出升高趋势。在50 mg/L铅浓度胁迫下[CM(25],G5、G8的铅积累量较高,显著高于其他基因型烟株(P<0.05);G2的铅积累量最低,显著低于其他基因型烟株(P<0.05)。在100 mg/L铅浓度胁迫下,G5的铅积累量显著高于其他基因型烟株(P<0.05),G2的铅积累量最低,其次是G6、G9、G10。
2.5不同基因型烤烟的镉、铅转运系数
由图5-a可知,随着镉胁迫水平的升高,部分基因型烟株的镉转运系数呈现出先增大后减小的趋势,说明低浓度处理有利于烟株向地上部分转运镉,而高浓度时由于胁迫程度的加剧导致烟株的镉转运系数减小。G15的镉转运系数随着镉浓度的升高表现出逐渐增大趋势,说明G15对重金属镉具有较强的富集能力。从不同基因型烟株的镉转运系数来看,在10 mg/L镉浓度胁迫下,G6、G12的镉转运系数与G7、G15无显著差异,但与其他基因型烟株差异显著(P<0.05)。在20 mg/L镉浓度胁迫下,G15的镉转运系数最大,且显著大于其他基因型烟株(P<0.05)。G3、G7在低镉胁迫水平下的镉转运系数较小,G4、G9在高镉胁迫水平下的镉转运系数较小。
由图5-b可知,随着铅胁迫水平的升高,不同基因型烟株的铅转运系数整体呈现减小趋势,说明随着铅浓度的提高不同基因型烟株的铅转运能力整体降低。从不同基因型烟株的铅转运系数来看,在50 mg/L铅浓度下,G15的铅转运系数最大,显著大于其他基因型烟株(P<0.05);在100 mg/L铅浓度下,G6、G15的铅转运系数较大,且显著大于其他基因型烟株(P<0.05),G3、G11的铅转运系数在2浓度水平下均较小,显著小于同水平下的其他基因型烟株(P<0.05)。
3讨论与结论
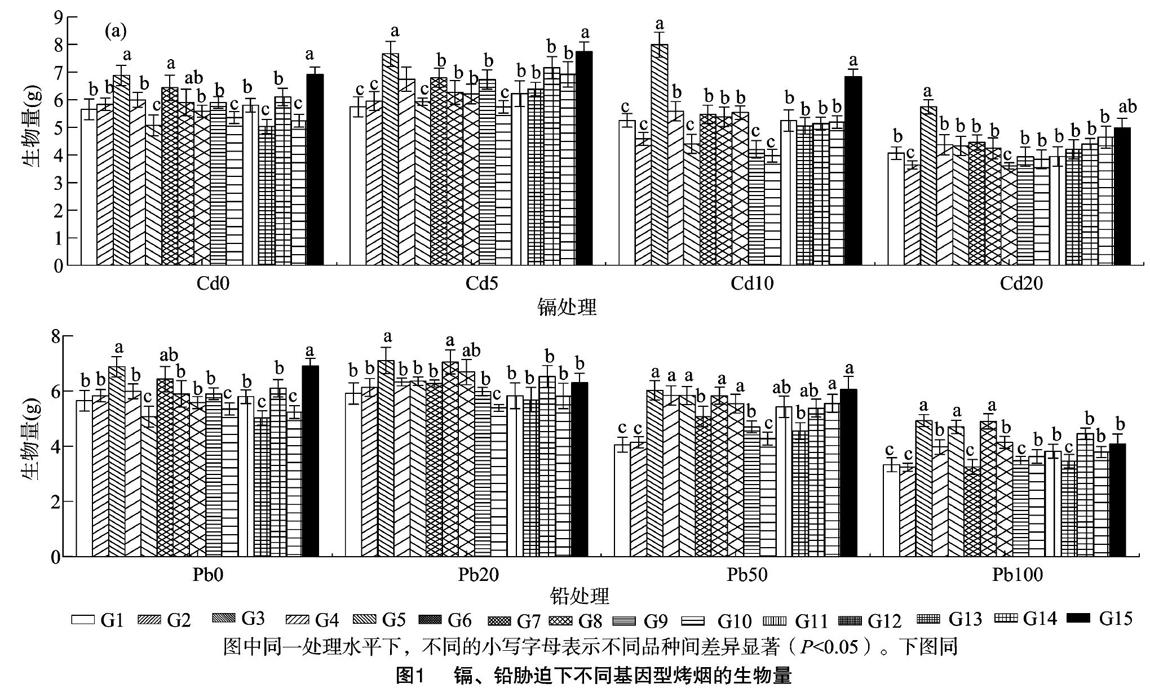
低积累作物的筛选和培育是土壤受重金属轻微、轻度污染修复的一种经济、有效途径[28],而高富集植物的筛选主要由植物地上部重金属富集量和适宜的生物量所决定[29]。本研究中的供试烟草材料主要为豫西烟区的主栽品种及部分国外引进品种,来源比较广泛且品种间存在较大的遗传差异,为镉、铅积累评价提供了必要条件。本研究结果表明,受镉、铅胁迫的15个基因型烤烟生物量、植物组织中镉含量、耐性指数、重金属积累量、转运系数存在不同的差异。同一镉、铅胁迫水平下不同基因型烟草生物量之间存在较大差异,不同镉、铅污染水平下同一基因型烟草生物量也会发生较大变化。15个基因型烤烟总体上对镉、铅污染具有较强的适应性,表现为低浓度的镉、铅促进不同基因型烟株生长,而高浓度镉、铅抑制烟株生长。轻度镉、铅污染能不同程度地促进烟草生物量的增加,可能與低浓度污染对烟草生长具有刺激作用有关,尤其是重金属镉,烟草对重金属镉具有较强偏好性。在轻度镉、铅胁迫刺激下,根部可能会分泌一些有机酸,从而促进植物吸收根际附近的矿质元素[10]。Liu等在盆栽条件下的研究表明,轻度镉污染有利于烟株生长,而高浓度镉胁迫显著抑制烟株生长[30]。烟株受轻、重度胁迫刺激的生长程度在不同基因型间也存在差异,表明不同基因型对环境的适应性存在不同的生态幅。随着污染胁迫的继续加剧,开始有部分基因型烟草生长受阻,导致生物量减少,其原因可能是这些镉、铅污染使相对敏感的烟草基因型在中、重度镉、铅污染下的叶绿体亚显微结构发生破坏,光合色素合成受到影响,阻碍电子传递,影[CM(25]响类囊体的蛋白质合成和参与卡尔文(Calvin)循环的酶活性,从而影响光合作用[31]。本研究发现,在镉污染条件下,G3、G15具有明显生长优势,G2、G8的生长较弱;而在20、50、100 mg/L铅污染下,G3、G7的生物量相对较高,中、重度铅污染下,G1、G2的生物量均较低,该结果可为烟草品种筛选提供首要依据。
利用不同指标评价植株对重金属的富集能力可能得到不同的结果[32]。一般来说,烟草体内镉、铅含量随着污染程度的增强而相应增加,但不同基因型烟株对重金属污染程度增强的响应不尽相同,同一基因型烟株不同器官之间的重金属积累也因胁迫强度的不同而有所差异。本研究中,G15在3个镉污染条件下地上部的镉含量较高,在20 mg/L镉浓度下,G15地上部分镉含量达到183.75 mg/kg,而G1、G2、G4、G10地上部和根部镉含量较低;在中、高度铅污染条件下,G5的地上部铅含量均最高,G2、G3的地上部铅含量较低。在中度铅污染水平下,G8根的铅含量较高,在重度铅污染水平下,G11根的铅含量较高;轻度铅污染条件下G4的根部铅含量最低。在100 mg/L铅浓度下,G2、G15根部的铅含量较低,分别为53.37、52.74 mg/kg。不同器官之间的镉含量存在差异,一般以叶部最高,根部次之;不同器官之间的铅含量存在差异,一般以根部最高,叶部次之。同一烟草材料在不同的重金属浓度下可能会表现出相似或相反的重金属耐性和积累能力[33]。Iannone等研究表明,在100、500 μmol/L镉处理下,野生型(SR1)和缺过氧化氢酶(CAT1AS)烟草植株镉含量表现出相反趋势[34]。本研究中,G1在20 mg/L铅浓度胁迫下植株铅含量较低,而在100 mg/L铅浓度下含量较高,这与上述研究结果相同。不同基因型烤烟材料重金属含量和积累量也存在显著差异。刘登璐等通过水培试验对不同烟草品种镉积累进行评价发现,在2、10 mg/L镉处理下地上部镉含量和积累量最大值分别为最小值的2.12、5.51倍和2.08、5.10倍[35]。本研究发现,在20 mg/L镉浓度胁迫下,高富集基因型烤烟地上部、根部的镉含量最大值分别是低富集基因型烤烟最小值的3.11、2.05倍;在100 mg/L铅浓度胁迫下,高富集基因型烤烟地上部、根部的铅含量最大值分别是低富集基因型烤烟最小值的1.83、2.00倍。耐性指数越大说明该品种对重金属的耐性越强[26]。在10 mg/L镉污染条件下,G3的镉耐性指数较高,G2、G9、G10、G13的镉耐性指数较低。在铅污染条件下,G5在3个污染水平下的铅耐性指数较高,G6的铅耐性指数较低。进一步分析表明,镉污染条件下镉积累量较高的是G3、G15,较低的是G2、G10;在100 mg/L铅污染条件下,G5的铅积累量显著高于其他基因型烟株,而G2、G6、G9、G10的铅积累量较低。张艳玲等分析发现,K326、云烟87、云烟85与中烟100等4个品种对重金属镉和铅的吸收积累不一致,但是无显著差异[36]。但本研究结果表明,部分基因型烤烟之间对重金属镉、铅的吸收积累存在显著差异,可能是因为供试烤烟基因型来源比较广泛且品种间存在较大的遗传差异。另外,雷丽萍等的研究表明,烟株对不同重金属的积累与品种有关[37]。
转运系数可用来评价植物将重金属从根部向地上部转运的能力[38]。本研究结果显示,G15在高镉水平下的镉转运系数较大,G1、G10、G12在低镉水平下的镉转运系数较大,而G3、G7在低镉水平下的镉转运系数较小,G3、G4、G9在高镉水平下的镉转运系数较小;在中、高度铅污染水平下,G3、G11的铅转运系数较小,且均显著小于其他基因型烟株(P<0.05),说明这些基因型烟株对铅具有较强的根际阻隔能力。与镉胁迫相比,铅胁迫条件下转运系数较小,说明烟株对镉元素的转运能力强于铅元素。镉、铅元素在不同器官中的分布将决定不同基因型的实际利用价值,植物生物量大且体内镉、铅积累量高的基因型,可作为植物修复的潜在选材;而烟株镉、铅积累量比较低的基因型,可满足烤烟安全生产的目的。本研究结果表明,在低镉水平下,G1、G2、G10的镉积累量是低镉污染条件下的低积累基因型,可作为在轻度镉污染条件下进行烟叶安全生产的良好材料。在高鎘、铅水平下,G5对铅的积累量、生物量、转运系数和耐性指数均较高,G15的镉积累量、转运系数较高,属于高积累品种,这些品种不利于在重金属镉、铅污染的土壤上安全生产烟叶,但可用于植物对土壤重金属的提取修复。在低铅水平下,G4、G12的铅积累量和地上部铅含量较低,属于铅低积累基因型,可用于重金属铅污染条件下安全生产烟叶。李晓婷等的研究也表明,不同烤烟品种对镉、铅的积累效应不同[39];鲁黎明等研究发现,不同品种烟草对铅富集的差异不明显,但对镉的积累却显著不同[40]。不同基因型烟株之间镉、铅富集能力和转运能力的差异,可能与不同基因型烟株对重金属镉、铅的偏好性及吸收积累重金属的内在机制不同有关,重金属敏感的基因型根际对镉、铅具有较好的阻控效应,可通过根细胞壁沉淀和分泌的代谢物质(有机酸和金属络合物)减少重金属向地上部分的运输;而对重金属富集能力较强的基因型可能在叶片细胞内具有较强的重金属解毒机制,若能够把重金属镉、铅存储在液泡中,将不会对烟株产生毒害[41-42]。
综上所述,15个基因型烤烟品种对镉、铅污染具有较强的适应性。低浓度的镉、铅胁迫可促进不同基因型烟株的生长,而高浓度镉、铅胁迫抑制烟株生长;但15个基因型烤烟品种对重金属镉、铅污染的响应不同,不同基因型烤烟品种对镉的积累分配主要在地上部分,而对铅的积累分配主要在根部,烟草对重金属镉的富集和转运能力高于重金属铅;烟草对重金属镉、铅的吸收积累能力存在基因型差异,是植物资源利用的最重要依据。在高镉、铅水平下,G15对镉的积累量、转运系数较高,G5的生物量以及对铅的积累量和耐性指数均较高,属于高积累、高耐性品种,这些品种不利于在重金属镉、铅污染的土壤上安全生产烟叶,但可用于重金属植物提取修复。G2、G10镉的积累量较低,是镉污染条件下的低积累基因型;在低铅水平下,G4、G12的铅积累量和地上部铅含量较低,属于铅低积累基因型,可用于轻度重金属铅污染条件下安全生产烟叶。科学认识与评价不同植物及基因型的耐逆性在生理生态上的差异并充分利用这些植物资源特质是植物修复与污染地再利用的根本原则。筛选资源、科学评价、杂交培育适合污染土壤安全生产的品种,是过去、现在及未来生态生理和生态育种学者不断努力的方向。植物对重金属富集和转运能力的差异,以及植物对重金属的阻遏、解毒机制仍需要广大学者深入研究。
参考文献:
[1]冯子龙,卢信,张娜,等. 农艺强化措施用于植物修复重金属污染土壤的研究进展[J]. 江苏农业科学,2017,45(2):14-20.
[2]Zhou H,Zeng M,Zhou X,et al. Heavy metal translocation and accumulation in Iron plaques and plant tissues for 32 hybrid rice (Oryza sativa L.) cultivars[J]. Plant and Soil,2015,386(1/2):317-329.
[3]Liu L,Zhang Q,Hu L L,et al. Legumes can increase cadmium contamination in neighboring crops[J]. PLoS One,2012,7(8):e42944.
[4]Fu F L,Wang Q. Removal of heavy metal ions from wastewaters:a review[J]. Journal of Environmental Management,2011,92(3):407-418.
[5]Liu L,Hu L L,Tang J J,et al. Food safety assessment of planting patterns of four vegetable-type crops grown in soil contaminated by electronic waste activities[J]. Journal of Environmental Management,2012,93(1):22-30.
[6]Cheraghi M,Lorestani B,Khorasani N,et al. Findings on the phytoextraction and phytostabilization of soils contaminated with heavy metals[J]. Biological Trace Element Research,2011,144(1/2/3):1133-1141.
[7]Lasat M M. Phytoextraction of toxic metals:a review of biological mechanisms[J]. Journal of Environmental Quality,2002,31(1):109-120.
[8]Ali H,Khan E,Sajad M A. Phytoremediation of heavy metals-concepts and applications[J]. Chemosphere,2013,91(7):869-881.
[9]Shen Z G,Li X D,Wang C C,et al. Lead phytoextraction from contaminated soil with high-biomass plant species[J]. Journal of Environmental Quality,2002,31(6):1893-1900.
[10]Ghosh U D,Saha C,Maiti M,et al. Root associated iron oxidizing bacteria increase phosphate nutrition and influence root to shoot partitioning of iron in tolerant plant Typha angustifolia[J]. Plant and Soil,2014,381(1/2):279-295.
[11]劉维涛,周启星. 重金属污染预防品种的筛选与培育[J]. 生态环境学报,2010,26(6):1452-1458.
[12]杨惟薇,刘敏,曹美珠,等. 不同玉米品种对重金属铅镉的富集和转运能力[J]. 生态与农村环境学报,2014,30(6):774-779.
[13]Meharg A A. Mechanisms of plant resistance to metal and metalloid ions and potential biotechnological applications[J]. Plant and Soil,2005,274(1/2):163-174.
[14]Chernobrovkina P N,Titov A F,Robonen E V,et al. Effect of boric acid on the ability of plants to accumulate heavy metals[J]. Russian Journal of Ecology,2012,43(1):29-32.
[15]李月芳,刘领,陈欣,等. 模拟铅胁迫下玉米不同基因型生长与铅积累及各器官间分配规律[J]. 农业环境科学学报,2010,29(12):2260-2267.
[16]Grant C A,Clarke J M,Duguid S,et al. Selection and breeding of plant cultivars to minimize cadmium accumulation[J]. Science of the Total Environment,2008,390(2/3):301-310.
[17]郭晓方,卫泽斌,丘锦荣,等. 玉米对重金属累积与转运的品种间差异[J]. 生态与农村环境学报,2010,26(4):367-371.
[18]凌天孝,李志鹏,张晓帆,等. 巫山烟区海拔高度对烤烟常规化学成分含量及其协调性的影响[J]. 江苏农业科学,2017,45(5):73-76.
[19]周冀衡,张建平. 构建中式卷烟优质特色烟叶原料保障体系是新形势下中国烟草的战略选择[J]. 中国烟草学报,2008,14(1):42-46,57.
[20]庞永强,李雪,罗彦波,等. 接装纸和成型纸透气度对卷烟中元素迁移行为的影响[J]. 烟草科技,2015,49(6):64-67,72.
[21]李继伟,周俊学,张喜文,等. 烤烟田休闲期5种绿肥的生物量及硫苷含量比较[J]. 湖北农业科学,2016,55(19):5056-5058,5132.
[22]廉勇. 豫西典型地段土壤重金属污染分析与评价[J]. 黄金,2015,36(6):65-68.
[23]李合生. 现代植物生理学[M]. 北京:高等教育出版社,2002.
[24]贺远,王树声,刘海伟,等. 镉浓度对烤烟幼苗镉含量及生长和生理指标的影响[J]. 中国烟草科学,2014(2):37-42.
[25]Zhang H J,Zhang X Z,Li T X,et al. Variation of cadmium uptake,translocation among rice lines and detecting for potential cadmium-safe cultivars[J]. Environmental Earth Sciences,2014,71(1):277-286.
[26]Ahmad I,Akhtar M J,Zahir Z A,et al. Organic amendments:effects on cereals growth and cadmium remediation[J]. International Journal of Environmental Science and Technology,2015,12(9):2919-2928.
[27]籍贵苏,严永路,吕芃,等. 不同高粱种质对污染土壤中重金属吸收的研究[J]. 中国生态农业学报,2014,22(2):185-192.
[28]雷恩,刘艳红. 个旧矿区周边农田高产、重金属低积累玉米品种的筛选[J]. 江苏农业科学,2015,43(9):124-127.
[29]熊国焕,何艳明,栾景丽,等. 龙葵、大叶井口边草和短萼灰叶对Pb、Cd和As污染农田的修复研究[J]. 生态与农村环境学报,2013,29(4):512-518.
[30]Liu L,Li Y,Tang J,et al. Plant coexistence can enhance phytoextraction of cadmium by tobacco (Nicotiana tabacum L.) in contaminated soil[J]. Journal of Environmental Sciences,2011,23(3):453-460.
[31]Bhaduri A M,Fulekar M H. Antioxidant enzyme responses of plants to heavy metal stress[J]. Reviews in Environmental Science and Bio/Technology,2012,11(1):55-69.
[32]王宝媛,濮阳雪华,宋桂龙,等. 20个高羊茅品种镉耐性评价及富集特征[J]. 草地学报,2017,25(1):107-114.
[33]魏益华,何俊海,冯小虎,等. 土壤重金属处理对烟草中As、Cd、Hg和Pb的累积与分布的影响[J]. 中国烟草学报,2016,22(1):47-54.
[34]Iannone M F,Groppa M D,Benavides M P. Cadmium induces different biochemical responses in wild type and catalase-deficient tobacco plants[J]. Environmental and Experimental Botany,2015,109:201-211.
[35]劉登璐,李廷轩,余海英,等. 不同烟草材料镉积累差异评价[J]. 农业环境科学学报,2016,35(11):2067-2076.
[36]张艳玲,尹启生,周汉平,等. 中国烟叶铅、镉、砷的含量及分布特征[J]. 烟草科技,2006(11):49-52,57.
[37]雷丽萍,夏振远,方敦煌,等. 玉溪烟叶几种重金属含量状况研究[J]. 西南农业学报,2011,24(4):1612-1614.
[38]曲荣辉,张曦,李合莲,等. 不同锌水平对低剂量镉在水稻中迁移能力的影响[J]. 中国生态农业学报,2016,24(4):517-523.
[39]李晓婷,常寿荣,徐洁,等. 不同烤烟品种吸收积累镉铅的差异性研究[J]. 西南大学学报(自然科学版),2014,36(6):16-21.
[40]鲁黎明,顾会战,彭毅,等. 不同品种烟草对重金属积累的动态差异[J]. 贵州农业科学,2013,41(4):40-43.
[41]罗洁文,黄玫英,殷丹阳,等. 类芦对铅镉的吸收动力特性及亚细胞分布规律研究[J]. 农业环境科学学报,2016,35(8):1451-1457.
[42]付铄岚,王昌全,李冰,等. 外源Cd在不同品种水稻组织中的细胞分布和化学形态特征研究[J]. 中国生态农业学报,2017,25(6):903-910.
