外源草酸缓解马尾松根系铝毒
2019-08-12刘亚敏周文颖刘玉民张盛楠
郭 妮,刘亚敏,周文颖,刘玉民,张盛楠
(西南大学 资源环境学院,三峡库区生态环境教育部重点实验室,重庆 400715)
铝是地壳中含量最丰富的金属元素,约占地壳总质量的7.1%,通常以无毒的硅酸盐等形式存在[1]。当土壤pH低于5.5时,土壤中的铝就会转变成对植物具有毒害作用的活性铝[2],铝毒害是制约酸性土壤作物生产力的主要因子之一。酸性土壤广泛分布于我国南方14个省(市、自治区),总面积约203万km2,占全国耕地面积的21%[3]。缓解植物铝毒害已成为土壤酸化区目前亟待解决的问题。
研究认为,植物铝毒缓解机制主要包括外部排斥和内部耐受2种:外部排斥机制是指阻止活性铝通过质膜进入共质体,从而防止其到达细胞内的敏感位点,使植物免遭铝毒害;内部耐受机制是指对于进入共质体的活性铝,植物通过螯合的方式将其转化为无毒性或者毒性很小的结合形态铝,从而缓解植物体内的铝毒害[4]。研究发现,荞麦(FagopyrumesculentumMoench.)受到Al3+刺激时根尖会迅速分泌草酸,与Al3+形成无毒草酸铝复合物,稳定贮存于液泡中,以维持植物细胞质中游离铝处于低水平[5-6];当荞麦根尖有机酸的分泌受抑制时,其根系受铝毒害较严重,也反过来证明有机酸分泌是植物耐铝胁迫的主要机制[7]。到目前为止,植物可通过分泌不同种类的有机酸来缓解铝毒已在大豆[Glycinemax(Linn.) Merr.][8]、小麦(TriticumaestivumL.)[9]、水稻(OryzasativaL.)[10]等植物上得到广泛证实,添加柠檬酸[11]、苹果酸[12]等外源有机酸可有效缓解植物铝毒害也被广泛报道。
马尾松(PinusmassonianaLamb.)是我国亚热带地区的主要用材树种之一,但近年来马尾松林出现大面积生长衰退甚至死亡的现象。有研究认为,南方地区酸雨加剧引起土壤铝活化,是影响马尾松生长从而导致马尾松大面积死亡的主要原因[13-14]。王水良等[15]研究发现,铝胁迫下的马尾松幼苗通过大量分泌草酸和苹果酸螯合根部有毒的Al3+形成稳定无毒的复合物来实现对铝胁迫的拮抗;另有研究发现,草酸主要通过分子上与碳原子相邻的两个羧基与铝结合形成五环结构来达到解毒的效果[16-17]。添加外源草酸可缓解铝毒已经在大豆[18]、小麦[19]、油菜(BrassicanapusL.)[20]等植物上得到证实,但这些研究主要是针对草本植物进行的,目前关于草酸对木本植物铝毒害是否具有缓解作用及其可能的机制等还不明确。为此,本试验以耐铝型和铝敏感型马尾松为材料,采用水培法研究不同浓度外源草酸对铝胁迫下马尾松幼苗根系形态及生理特性的影响,探讨草酸对马尾松根系铝毒的缓解作用和最适缓解浓度,揭示草酸对马尾松铝毒的缓解作用机制,旨在为酸性土壤中马尾松林的铝毒治理提供一定的理论依据。
1 材料与方法
1.1 试验材料
经前期筛选,本试验分别选取耐铝型福建5号(FJ5)和铝敏感型广东20号(GD20)马尾松作为试验材料,试验材料的种子由福建省潭平五一国有林场提供。
选取大小一致、圆润饱满的FJ5和GD20马尾松种子,经2% H2O2消毒10 min后,流水冲洗20 min,再用去离子水清洗3次,25 ℃水浴锅避光浸种24 h,之后将种子均匀播于铺有一层珍珠岩的培养皿中,恒温培养箱25 ℃催芽至露白。露白后,将种子播于装有珍珠岩的育苗盘中,于培养室(恒温25 ℃,空气湿度60%,光照强度100 μmol·m-2·s-1)继续培养,每日光照时间6:00—18:00,定时浇入适量的蒸馏水。待马尾松第一轮针叶完全打开后,选取长势一致的幼苗,用纱布固定于避光的玻璃杯(200 mL)盖上,每瓶3株,用1/5 Hoagland完全营养液(pH值调至4.0)培养,每7 d更换一次营养液,培养6个月即得到试验所需幼苗[21]。
1.2 试验设计
两个马尾松家系分别设置7个处理(表1),每个处理设置3组重复,每组6杯,每杯定植3株。供试有机酸选用草酸(OA),Al3+以AlCl3(分析纯)形式加入。处理60 d后测定各指标。
1.3 测定方法
马尾松总根长、根表面积、根系体积和根直径采用根系分析仪测定,根系活力采用氯化三苯酚四氮唑(TTC)法[22]测定,超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)光化还原法[23]测定,过氧化物酶(POD)活性采用愈创木酚法[24]测定,过氧化氢酶(CAT)活性、脯氨酸(Pro)含量参照张蜀秋[24]的方法测定,丙二醛(MDA)和可溶性糖含量参照张志良等[25]的方法测定,可溶性蛋白含量采用考马斯亮蓝(G-250)法[26]测定。根尖和根系中铝含量参照陈荣府[27]的方法进行前处理后采用电感耦合等离子体发射光谱仪(ICP-OES)测定,其中,根系铝含量以鲜重计,根尖铝含量以单个根尖含铝的物质的量计。
1.4 主成分分析
表1 试验设计与处理
Table 1 Experiment design
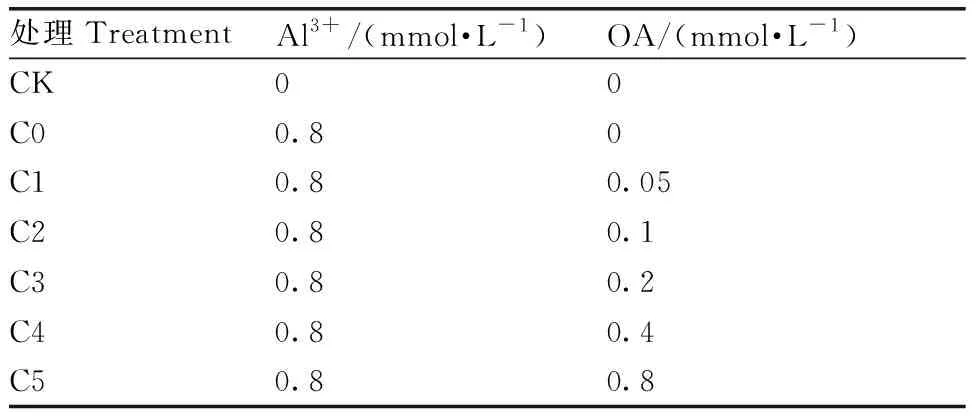
处理TreatmentAl3+/(mmol·L-1)OA/(mmol·L-1)CK00C00.80C10.80.05C20.80.1C30.80.2C40.80.4C50.80.8
选择铝敏感型GD20马尾松的生理指标进行主成分分析。分析前,先将数据进行标准化处理,转为无量纲矩阵后再进行统计分析,计算主成分特征根、方差贡献率。以特征根>1、方差累积贡献率>80%为提取标准,确定主成分数量[28]。根据各生理指标在主成分中的贡献率,分析草酸缓解马尾松铝毒害的关键指标。
标准化处理公式为
(1)
1.5 数据处理
利用Excel 2016软件对数据进行整理。运用SPSS 19.0软件进行单因素方差分析和主成分分析,对有显著差异(P<0.05)的处理采用LSD法进行多重比较。采用Origin 8.5软件制图。
2 结果与分析
2.1 草酸对马尾松根系铝毒的缓解效果
2.1.1 对根系形态的影响
Al3+对马尾松的根系生长有抑制作用,且对铝敏感型马尾松根系生长的抑制作用要强于耐铝型,FJ5在C0处理下的总根长、根表面积、根系体积、根直径较CK分别降低20.38%、24.53%、31.25%、10.26%,GD20在C0处理下的总根长、根表面积、根系体积、根直径较CK分别降低29.11%、33.49%、50.00%、21.28%。施加外源草酸后,马尾松受到的铝毒害均得到一定程度的缓解,随着草酸施入量的增大,缓解效果总体呈现出先上升后下降的趋势(表2)。当外源添加的草酸浓度为0.2 mmol·L-1时对耐铝型FJ5的缓解效果最好,植株总根长、根表面积、根体积及平均直径分别比C0处理显著(P<0.05)增加22.12%、33.20%、54.54%和14.28%,且与CK相比已无显著差异。铝敏感型马尾松GD20的总根长和根直径在C3处理下获得加铝条件下的最大值,分别比C0显著(P<0.05)增加39.12%和16.22%,但根表面积和根系体积在各浓度草酸处理下与C0相比均无显著差异。
2.1.2 对根系活力的影响
表2 不同处理对马尾松根系形态的影响
Table 2 Effects of different treatments on root morphology ofP.massonianaLamb.

试验材料Material处理Treatment总根长Root length/cm根表面积Total surface area/cm2根系体积Root volume/cm3根直径Root diameter/cmFJ5CK139.86 a17.00 a0.16 a0.39 aC0111.35 c12.83 c0.11 c0.35 bC1116.72 bc13.77 bc0.13 bc0.38 aC2123.91 abc15.03 abc0.15 ab0.39 aC3135.98 ab17.09 a0.17 a0.40 aC4133.45 abc15.86 ab0.15 ab0.38 aC5118.81 abc14.85 abc0.15 ab0.40 aGD20CK131.02 a19.32 a0.24 a0.47 aC092.88 c12.85 b0.12 b0.37 cC1123.39 ab15.15 ab0.15 b0.39 bcC2125.64 ab15.86 ab0.16 b0.40 bcC3129.21 ab14.57 ab0.15 b0.43 abC4128.10 ab15.70 ab0.15 b0.39 bcC5100.40 bc12.97 b0.13 b0.42 bc
同一试验材料同列数据后无相同字母的表示处理间差异显著(P<0.05)。下同。
Data marked without the same letters in the same column indicated significant difference atP<0.05 within treatments for the same material. The same as below.
植物根系活力是体现植物根系对矿物质营养吸收能力和呼吸作用强度的重要指标。铝胁迫降低了马尾松的根系活力值,与CK相比,C0处理下铝敏感型马尾松GD20和耐铝型马尾松FJ5的根系活力分别显著(P<0.05)降低64.07%、59.26%,加入外源草酸后,马尾松的根系活力有所提高,随外源草酸添加浓度增加总体呈现先上升后下降的趋势(图1)。当外源添加浓度为0.2 mmol·L-1时,草酸对马尾松铝毒害的缓解效果最好,铝敏感型马尾松GD20和耐铝型马尾松FJ5的根系活力分别为0.23、0.30 mg·g-1·h-1。外源草酸对耐铝型马尾松的作用效果更佳,C3处理下,其根系活力与CK相比已无显著差异。
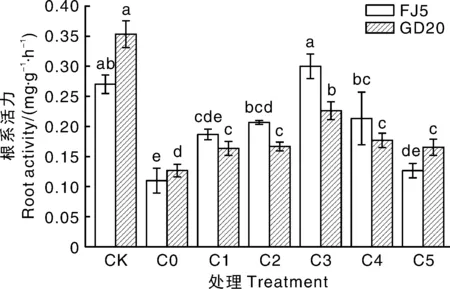
同一家系柱上无相同字母的表示处理间差异显著(P<0.05)。下同。Bars marked without the same letters in the same family indicated significant difference at P<0.05. The same as below.图1 不同处理下马尾松的根系活力Fig.1 Root activity of P. Massoniana Lamb. under different treatments
2.1.3 对根系抗氧化酶活性的影响
逆境下,植物通过抗氧化酶协同清除体内各种活性氧自由基,以保护膜系统,减轻逆境危害。与CK相比,C0处理下马尾松的根系SOD、POD、CAT活性显著(P<0.05)上升。随着外源添加草酸浓度增加,马尾松根系SOD活性整体呈现先增加后下降的趋势。当外源添加草酸浓度为0.2 mmol·L·1时,铝敏感型马尾松GD20和耐铝型马尾松FJ5的根系SOD活性均达到最大值,显著(P<0.05)高于C0处理。但高浓度的外源草酸(0.8 mmol·L-1)导致耐铝型马尾松FJ5的根系SOD活性显著低于C0处理(图2-A)。

图2 不同处理对马尾松根系抗氧化酶活性的影响Fig.2 Effects of different treatments on antioxidant enzymes in roots of P. massoniana Lamb.
随着外源添加草酸浓度增加,马尾松根系POD活性同样呈现先增加后下降的趋势。当外源添加草酸浓度为0.1~0.2 mmol·L·1时,铝敏感型马尾松GD20和耐铝型马尾松FJ5根系的POD活性均显著(P<0.05)高于C0处理。耐铝型马尾松FJ5根系POD活性在不同浓度草酸处理间差异不显著,但铝敏感型马尾松GD20在C2、C3处理下的根系POD活性显著(P<0.05)高于其他外源添加草酸的处理,且高浓度的外源草酸(0.8 mmol·L-1)导致铝敏感型马尾松GD20的根系POD活性显著(P<0.05)低于C0处理(图2-B)。
外源添加0.05~0.2 mmol·L-1的草酸后,耐铝型马尾松FJ5的根系CAT活性与C0相比无显著变化;而当外源添加0.4~0.8 mmol·L-1的草酸后,耐铝型马尾松FJ5的根系CAT活性与C0相比则显著(P<0.05)降低,且显著(P<0.05)低于CK处理。与此不同,随着外源添加草酸浓度增加,铝敏感型马尾松GD20根系的POD活性则呈现先增加后下降的趋势。当外源添加草酸浓度为0.2 mmol·L·1时,铝敏感型马尾松GD20的根系CAT活性达到最大值,显著(P<0.05)高于其他处理;当外源添加0.4~0.8 mmol·L-1的草酸后,铝敏感型马尾松GD20的根系CAT活性与C0相比显著(P<0.05)降低,C4处理的根系CAT活性与CK无显著差别,而C5处理的根系CAT活性则显著(P<0.05)低于CK处理(图2-C)。
2.1.4 对根系渗透调节物质的影响
与CK相比,铝胁迫下(C0处理)的马尾松根系脯氨酸含量显著(P<0.05)增加,铝敏感型马尾松GD20和耐铝型马尾松FJ5的根系脯氨酸含量分别为CK的1.73和1.71倍(图3-A)。除外源添加0.2 mmol·L-1的草酸后,耐铝型马尾松FJ5的根系脯氨酸含量与C0相比显著(P<0.05)降低外,其他处理的根系脯氨酸含量与C0相比均无显著差异。与此不同,外源添加草酸后,铝敏感型马尾松GD20的根系脯氨酸含量与C0相比均显著(P<0.05)降低,但仍显著(P<0.05)高于CK处理,且C4、C5处理下根系脯氨酸含量显著(P<0.05)高于C1~C3处理。
C0处理下铝敏感型马尾松GD20和耐铝型马尾松FJ5的根系可溶性蛋白含量分别为CK的1.37倍和1.93倍,说明铝胁迫刺激马尾松根系可溶性蛋白含量显著(P<0.05)升高(图3-B)。外源添加草酸后,马尾松根系的可溶性蛋白含量与C0相比均显著(P<0.05)降低,与C0相比,耐铝型马尾松FJ5的根系可溶性蛋白含量降低39.4%~62.8%,铝敏感型马尾松GD20的根系蛋白含量降低27.1%~54.3%。C1~C3处理的耐铝型马尾松FJ5根系可溶性蛋白含量间无显著差异,均显著(P<0.05)低于C5处理,且C4处理的根系可溶性蛋白含量显著(P<0.05)低于C5处理。外源添加0.2 mmol·L-1草酸处理下,铝敏感型马尾松GD20的根系可溶性蛋白含量最低,显著(P<0.05)低于其他处理,C1、C2、C4处理的根系可溶性蛋白含量无显著差异,但均显著(P<0.05)低于C5处理。

图3 不同处理对马尾松根系渗透调节物质的影响Fig.3 Effects of different treatments on osmotic adjustment substances in roots of P. massoniana Lamb.
与CK相比,C0处理下马尾松根系可溶性糖含量显著(P<0.05)增加,且对铝敏感型马尾松的影响更大,GD20的根系可溶性糖含量较CK组增加94.8%(图3-C)。外源添加草酸各处理下,耐铝型马尾松FJ5的根系可溶性糖含量与C0相比无显著变化。但外源添加草酸后,铝敏感型马尾松GD20的根系可溶性糖含量均较C0处理显著(P<0.05)降低,并以C2、C3处理的根系可溶性糖含量最低,且显著(P<0.05)低于其他外源添加草酸的处理。
2.1.5 对根系膜脂过氧化的影响

图4 不同处理对马尾松根系MDA含量的影响Fig.4 Effects of different treatments on MDA content in root of P. massoniana Lamb.
MDA含量是反映植物膜脂过氧化程度的指标,可反映植物受胁迫或损伤的程度。与CK相比,C0处理下马尾松根系的MDA含量均显著(P<0.05)上升,说明铝胁迫下马尾松根系的膜脂发生过氧化(图4)。外源添加0.05~0.2 mmol·L-1的草酸后,耐铝型马尾松FJ5的根系MDA含量较C0显著(P<0.05)下降,与CK无显著差别;外源添加0.4~0.8 mmol·L-1的草酸后,耐铝型马尾松FJ5的根系MDA含量与C0、CK均无显著差别。外源添加0.05~0.4 mmol·L-1的草酸后,铝敏感型马尾松GD20的根系MDA含量较C0显著(P<0.05)下降,但仍显著(P<0.05)高于CK处理,其中,C3处理的根系MDA含量最低,且显著(P<0.05)低于其他外源添加草酸的处理;外源添加0.8 mmol·L-1的草酸后,铝敏感型马尾松GD20的根系MDA含量较C0显著(P<0.05)上升。
2.1.6 对根系和根尖铝含量的影响
如图5所示,各处理下铝敏感型马尾松GD20根系及根尖中的Al含量均高于耐铝型马尾松FJ5。外源添加草酸显著(P<0.05)降低了马尾松根尖的铝含量。随着外源添加草酸浓度的增加,根尖铝含量呈现出先降低后增加的趋势,当外源添加草酸浓度为0.2 mmol·L-1时,铝敏感型马尾松GD20和耐铝型马尾松FJ5根尖铝含量较C0处理分别降低76.5%和75.1%。外源添加0.1~0.2 mmol·L-1草酸后,铝敏感型马尾松GD20根系铝含量较C0处理显著(P<0.05)下降;但当外源添加草酸的浓度增大至0.4~0.8 mmol·L-1后,铝敏感型马尾松GD20的根系铝含量较C0处理反而显著(P<0.05)增加。对耐铝型马尾松FJ5而言,仅当外源添加草酸浓度为0.2 mmol·L-1时根系铝含量较C0处理显著(P<0.05)下降,当外源添加草酸浓度为0.8 mmol·L-1时根系铝含量较C0处理显著增加(P<0.05),其他处理下根系铝含量与C0处理并无显著差异。
2.2 草酸缓解马尾松根系铝毒的主导生理因子

图5 不同处理对马尾松根系(A)和根尖(B)铝含量的影响Fig.5 Effects of different treatments on Al content in roots (A) and root tips (B) of P. massoniana Lamb.
采用主成分分析法,对铝敏感型马尾松GD2的SOD、POD、CAT活性,根系活力,以及脯氨酸、可溶性蛋白、可溶性糖、MDA含量等8个生理指标进行分析。结果表明,前2个成分的贡献率分别为51.835%、34.867%(表3),累计贡献率达86.701%,表明前2个成分代表了原有8个指标86.701%的信息,可以用这2个成分概括分析外源草酸对铝胁迫下马尾松根系生理特性的影响。在主成分载荷矩阵中,特征向量的绝对值反映了其对主成分的贡献率,绝对值越大,则贡献率越大。由表4可知,脯氨酸、可溶性蛋白、可溶性糖含量在第一主成分上有较高的载荷,说明第一主成分主要反映马尾松根系渗透调节物质的信息;SOD、POD、CAT活性在第二主成分上有较高载荷,说明第二主成分主要反映抗氧化酶活性的信息。综上,脯氨酸和可溶性蛋白含量可能是上述8个生理指标中具有较高代表性的指标,可以作为外源草酸缓解马尾松铝毒害的关键指标。
表3 主成分分析结果
Table 3 Principle component analysis result
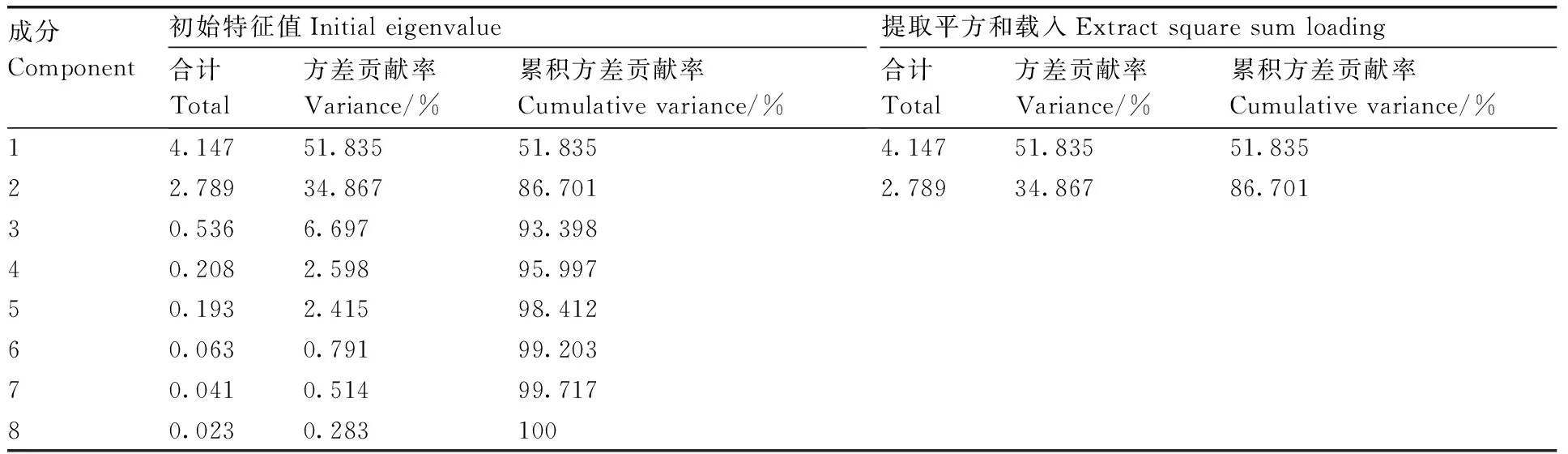
成分Component初始特征值Initial eigenvalue合计Total方差贡献率Variance/%累积方差贡献率Cumulative variance/%提取平方和载入Extract square sum loading合计Total方差贡献率Variance/%累积方差贡献率Cumulative variance/%14.14751.83551.8354.14751.83551.83522.78934.86786.7012.78934.86786.70130.5366.69793.39840.2082.59895.99750.1932.41598.41260.0630.79199.20370.0410.51499.71780.0230.283100
表4 主成分载荷矩阵
Table 4 Component matrix of principal component analysis
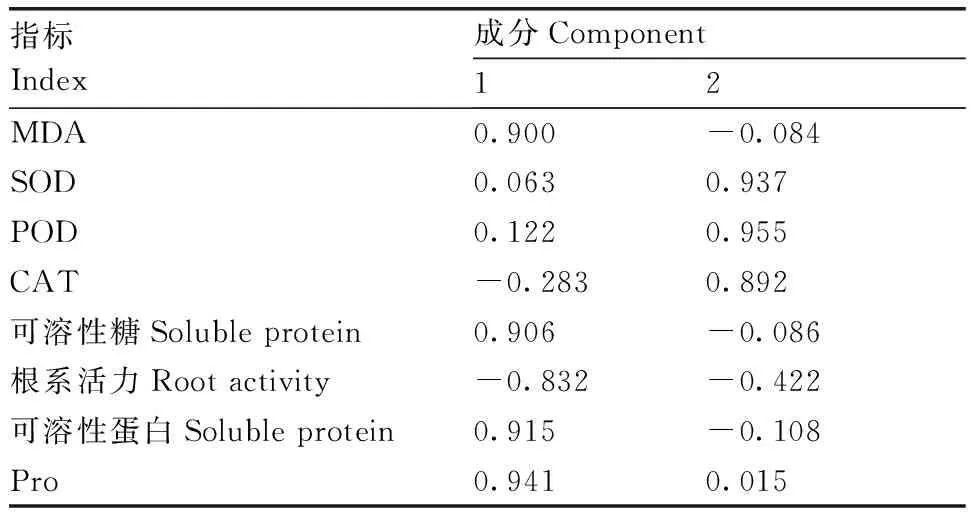
指标Index成分Component12MDA0.900-0.084SOD0.0630.937POD0.1220.955CAT-0.2830.892可溶性糖Soluble protein0.906-0.086根系活力Root activity-0.832-0.422可溶性蛋白Soluble protein0.915-0.108Pro0.9410.015
3 讨论

逆境胁迫对植物的次生伤害为渗透胁迫。植物遭受逆境胁迫时,会积累一些可溶性的无机和有机渗透调节物质,如脯氨酸、可溶性蛋白质和可溶性糖等,降低渗透势,使其在低渗透势生境下仍能吸收水分,维持细胞水分平衡,保护植株正常生长[36]。本研究结果表明,铝胁迫下马尾松根系脯氨酸、可溶性蛋白、可溶性糖含量显著升高,这与吴成龙等[37]研究结果一致,表明植物根系对外界胁迫敏感,胁迫下会将大量渗透调节物质运送到根系,导致根系渗透调节物质积累。可溶性糖含量的增加主要是通过不溶性碳水化合物的降解及其转化实现,可溶性蛋白含量升高则主要是由于铝胁迫诱导了植物抗性蛋白的产生。外源添加草酸后,马尾松根系脯氨酸、可溶性蛋白、可溶性糖含量均显著降低。可溶性糖和可溶性蛋白含量降低可能是因为:一方面,有机酸缓解了铝毒对马尾松膜脂过氧化的抑制作用,使根系细胞渗透物质含量恢复正常;另一方面,因为草酸缓解了植物铝毒,植物得以正常生长,此时需要大量能源,可溶性糖作为能源物质被马尾松大量吸收利用,而可溶性蛋白多是与膜系统特异性结合的酶,主要用来进行生理生化等代谢活动[38]。草酸对马尾松根系渗透调节物质的影响呈现浓度差异,随着草酸浓度的增加,3种渗透调节物质均呈现先降低后增加的趋势,在草酸浓度为0.2 mmol·L-1时有最小值,高浓度草酸处理反而会使渗透调节物质含量增加,这与姚虹宇等[39]在柠檬酸缓解马尾松铝毒研究中的结果一致,说明高浓度有机酸会抑制马尾松铝毒害的缓解。
总的来看,活性铝且对马尾松幼苗产生一定的毒害作用,并使马尾松根系生理代谢受到影响。铝胁迫下,马尾松幼苗总根长、根表面积、根系体积、根系活力等均有所降低,根系抗氧化酶(POD、SOD和CAT)活性、渗透调节物质(可溶性糖、可溶性蛋白和脯氨酸)含量均升高,膜质过氧化程度加剧。添加适宜浓度的外源草酸能有效缓解马尾松幼苗的铝毒害作用,使马尾松根系代谢系统得到恢复。添加外源草酸后马尾松根系活力升高,根系中丙二醛和渗透调节物质含量降低,抗氧化物酶活性维持较高水平,根系铝和根尖铝的含量减低(草酸浓度≤0.2 mmol·L-1),且对铝敏感型马尾松的缓解效果好于耐铝型。草酸缓解马尾松根系铝毒害存在浓度效应,以外源添加草酸浓度为0.2 mmol·L-1时缓解效果最好。脯氨酸和可溶性蛋白等渗透调节物质可能是调控草酸缓解马尾松铝毒害的主导生理因子。
