粤东亚热带森林群落树种组成对苔藓植物分布的影响
2019-08-08杨学成周庆唐启明徐明锋苏志尧
杨学成,周庆,唐启明,徐明锋,苏志尧
(华南农业大学林学与风景园林学院,广州 510642)
森林中苔藓植物是生物多样性的重要组成部分,也是增加森林结构复杂性的重要生物因素[1-4]。因此,对森林中苔藓植物开展研究有助于完整认识森林生态系统,从而为提高森林生态系统服务功能提供指导。
有研究表明不同树种组成的不同类型森林中苔藓植物的群落组成特征存在差异[5-10]。在诸多影响因子中,树种组成特征是引起苔藓植物物种组成差异的重要因素之一。Spitale[10]的研究揭示了苔藓植物分布与森林类型的生境关联,并分析了导致这种生境关联的内在环境因素。他认为基质类型的复杂性是决定苔藓多样性和物种组成的重要因素,这是由于研究区域大多数物种对基质选择存在专一性;针、阔叶森林的一些内在特征通过影响基质的理化性质(如树皮特征、林地表层凋落物特征等)会对苔藓多样性和组成产生影响;林内的小气候条件则通过影响基质的水分条件而影响苔藓植物的多样性和组成。Gosselin等[9]在3种不同优势树种组成的森林中开展的研究表明,松树林苔藓植物多样性显著低于橡树林和混交林,但后两者不存在明显差别,并且认为与基质类型相关的水化学效应是引起苔藓植物群落物种组成变化的主要因素之一。
在林分尺度上认识苔藓植物组成特征与树种组成特征的关系,不仅有助于我们明确苔藓植物与林分尺度因子的相互关系,而且从森林经营来说,林分作为林业经营的基本单元,明确林分尺度苔藓植物的生境关联无疑有助于我们加深对森林生物多样性的认识,从而在林分尺度上制定合理的经营保育策略。因此,本研究以粤东亚热带地区森林中的苔藓植物为研究对象,首先利用多元统计方法根据树种组成区分不同的林分类型,然后在此基础上探究苔藓植物种类组成对不同优势树种组成的林分的响应,以期明确苔藓植物在种类组成方面如何响应林分类型的变化。目前在本地区已开展了苔藓植物的分类学、特定地点的区系成分和群落生态学特征的相关研究[11-16],但森林中苔藓植物组成的研究还相对匮乏。因此本研究在完善对本地区森林群落的整体认识方面也具有重要的现实意义。
1 材料和方法
1.1 研究区概况
样地位于广东省东源康禾省级自然保护区(23°44′~23°53′N,115°04′~115°09′E),总面积为7 263.9 hm2。保护区地形属于低山陵地带,总体呈东北-西南走向,地貌复杂,地势起伏较大,最高点海拔为839.7 m。其气候属于亚热带季风气候,光热充足,雨量充沛,年平均气温为20.3℃~21.1℃,无霜期达350 d,年平均降水量为1 889.9 mm。区内土壤有机质积累较多,主要类型为赤红壤、红壤、山地黄壤和草甸土,且大部分土壤深厚疏松,湿润肥沃。自然保护区内植物资源丰富,据保护区统计,维管植物共有131科333属454种,其中双子叶植物、单子叶植物、蕨类、裸子植物各占总种数的70.3%、15.5%、8.4%和0.9%。区内植被类型丰富,地带型植被以亚热带常绿阔叶林为主,为苔藓植物提供了复杂多样的生境。
1.2 数据收集
参照热带森林大型固定样地调查规范[17],在保护区内设置了6 hm2的固定样地,大小为300 m×200 m的矩形,横轴(x轴)为西南-东北走向。使用Nikon DTM-310全站仪将6 hm2固定样地按水平距离划分为150个20 m×20 m的大样方,在每个样方的四角及中心顶点采用PVC管作为角桩标记固定。样方顶点角桩的编号从面向样地基准方向左下角的第一个单元起,从下往上顺着坐标轴方向依次为P001,P002,…… P150。对每个大样方进行每木检尺,记录胸径≥1 cm植株的种名、胸径、树高和冠幅。在每个大样方中设置5个2 m×2 m中样方,分别位于网格样方中心及对角线四分处。再将每个中样方等分为4个1 m×1 m的小样方。记录小样方内所有地面苔藓植物的种名、盖度和生境。苔藓物种名称及科属归类及分布主要根据《广东苔藓志》的资料进行整理[13,15]。
1.3 数据分析
林分的分类与排序以各样方乔木层树种(胸径≥10 cm)个体的相对基盖度建立数据集。基盖度以胸高断面积为指标。相对基盖度是样方某一树种胸高断面积与该样方总胸高断面积的比值。然后利用双向聚类对样地中的样方进行分类。在分类的基础上再进行多响应置换程序(MRPP)和排序分析,进一步确定林分分类的合理性。排序分析采用主坐标(PCoA)分析方法;MRPP和PCoA分析均在PC-ORD(6.0)软件中进行。
林分树种组成特征分析在对林分进行分类的基础上,通过计算各类型林分中物种的重要值分析各林分树种组成特征。物种的重要值(IV)=相对频度(RF)+相对优势度(RD)+相对多度(RA),优势度以盖度为测度单位。
苔藓物种组成特征分析以林分分类与排序为基础,分别对各类型林分中苔藓群落物种组成进行分析,计算每一种苔藓的重要值。不同类型林分中苔藓植物群落物种组成差异的显著性分析采用MRPP方法。
2 结果和分析
2.1 林分的分类和排序
在样地150个大样方乔木层中共出现69种胸径≥10 cm的树种。根据各树种在样方内的相对基盖度,利用双向聚类方法可首先将150个样方基本区分为3种林分类型(图1)。从树种组成的角度看,属于林分类型Ⅰ的样方为43个,占样方总数的28.7%,表现出以翻白叶树(Pterospermum heterophyllum)为优势种,南酸枣(Choerospondias axillaris)为次优种的树种组成特征;属于林分类型Ⅱ的样方为20个,占样方总数的13.3%,表现出以木荷(Schima superba)和米锥(Castanopsis carlesii)为共优种,红背锥(C.fargesii)为次优种的树种组成特征;属于林分类型Ⅲ的样方数为87个,占样方总数的58.0%,表现出以米锥占绝对优势,木荷为次优种的树种组成特征(表1)。多响应置换程序的分析结果也进一步表明3种林分存在显著差异(表2)。
图2为以双向聚类分析结果对样地乔木层树种组成进行的主坐标分析排序图。从主坐标排序的结果看,3种林分类型的样方在排序图上的位置存在明显的差异,体现出以样方树种组成进行的排序结果能很好地反映环境因子梯度;也进一步说明双向聚类验证了林分类型分类结果的合理性。
2.2 苔类植物组成对不同林分类型的响应
3种林分中分别有13、11和13种苔类植物出现。表3为3种林分中的相对优势度>5%的苔类植物的重要值。从优势物种的组成来看,林分类型Ⅰ中以双齿裂萼苔(Chiloscyphus latifolius)为优势种,细指苔(Kurzia gonyotricha)为次优种;林分类型Ⅱ中以双齿裂萼苔和细指苔为共优势种,神山细鳞苔(Lejeunea eifrigii)和变异多褶苔(Spruceanthus polymorphus)为共次优种;林分类型Ⅲ中以细指苔为绝对优势种,双齿裂萼苔次优种的物种组成特征。多响应置换程序分析表明,3种林分的苔类植物群落组成存在显著差异,且林分类型Ⅲ和另两种林分类型的差异极显著(表4)。
2.3 藓类植物组成对不同林分类型的响应
3种林分中分别有藓类植物26、23和29种,分别占藓类植物总数的81.3%、71.9%和90.6%。表5为3种林分中相对优势度大于5%的藓类植物重要值。从优势物种来看,3种林分均以东亚拟鳞叶藓(Pseudotaxiphyllum pohliaecarpum)为优势种,粗肋凤尾藓(Fissidens laxus)为次优种。但相对而言,林分类型Ⅱ中还有2个次优种细叶小羽藓(Haplocladium microphyllum)和锦藓(Sematophyllum subpinnatum),使得优势种东亚拟鳞叶藓在群落中的优势地位相对下降。多响应置换程序分析表明,3种林分的藓类植物群落组成总体上存在显著差异,主要体现在林分类型Ⅱ和另两种林分间(表6)。
3 讨论
本研究结果表明,粤东亚热带地区森林群落物种组成对苔藓植物群落的物种组成产生显著的影响。在森林中,生长在多样化基质上的体型微小的苔藓植物是林下植被的重要组成部分,上层植物群落特征,如年龄结构、树种组成、结构和经营强度等都会影响林下的生境条件,从而影响到苔藓植物群落的相关特征[18]。本研究从森林群落树种组成的角度开展研究,按照树种组成可划分为以翻白叶树(I)、木荷+米锥(II)、米锥(III)为优势种的3种林分类型;3种类型森林中苔类和藓类植物群落均存在显著差异。这与前人的研究结论是一致的,均表明森林群落的树种组成是影响苔藓植物群落的重要生物因素[6,8-10],表现出本地区苔藓群落组成与森林群落树种组成的关联,实际上森林群落树种组成和林分所构成的环境就成为影响苔藓植物分布的生物生境(biotic habitat)。这种生境关联的存在对未来开展森林经营和苔藓植物保育具有重要的理论指导意义,即通过这种关联在明确生产林木树种组成的基础上就可以相对明确地把握苔藓植物群落的物种组成特征。

图1 基于林分树种组成的样方双向聚类分析。横轴为种名,纵轴为样方号;灰度的深浅代表相对基盖度的大小。Fig.1 Two-way cluster dendrogram for plots based on tree species composition of the forest stands.X-axis stands for tree species,while Y-axis for plot ID;the greyscale in the squares represent greater to smaller values of relative basal area.
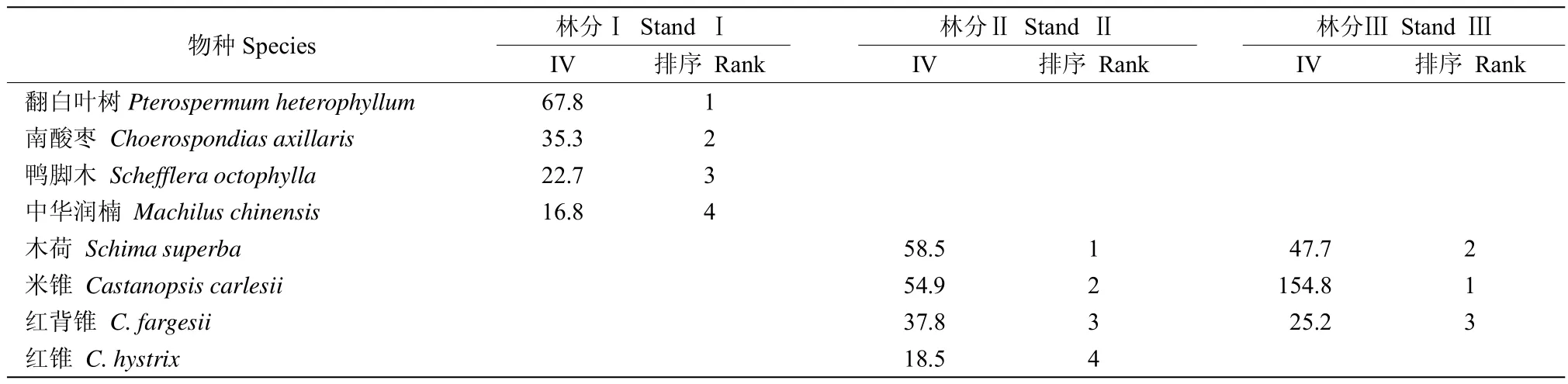
表1 相对优势度大于5%的乔木层树种的重要值(IV)Table1 Importance value(IV)of species in tree layer with relative dominance>5%
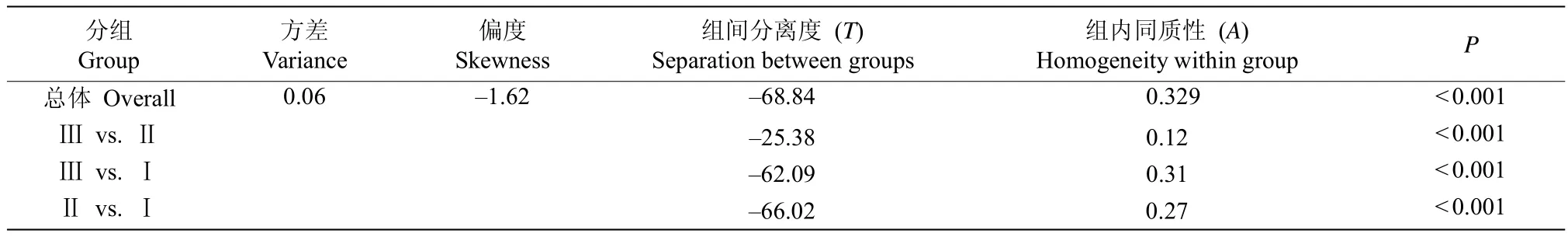
表2 3种林分乔木层树种群落组成的多响应置换程序分析Table2 Multi-response permutation procedures(MRPP)for the variations in species composition of canopy tree species across three stands
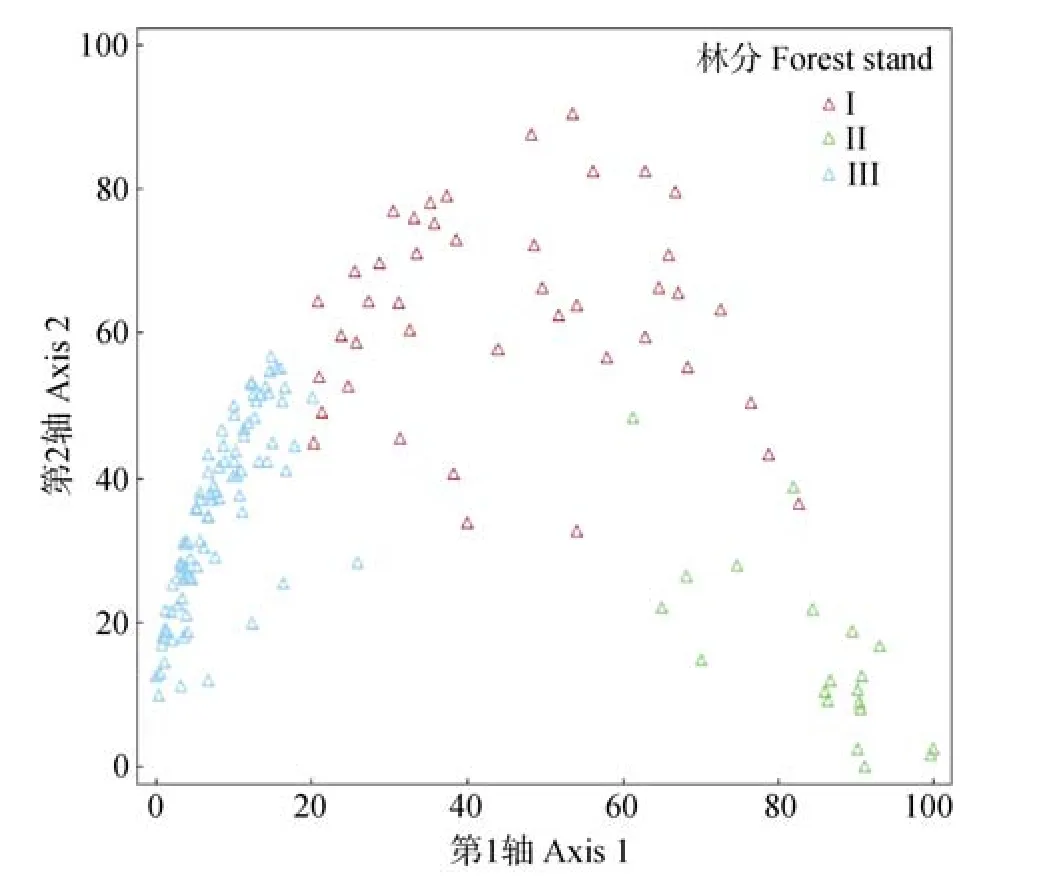
图2 基于双向聚类的样地乔木层树种组成主坐标分析排序Fig.2 Ordination by principal coordinate analysis(PCoA)of canopy tree species by the two-way cluster analysis from the surveys

表3 相对优势度大于5%的苔类植物重要值(IV)Table3 Importance value(IV)of liverwort species with relative dominance>5%

表4 3种林分苔类植物群落组成的多响应置换程序分析Table4 Multi-response permutation procedures(MRPP)for the variations in species composition of liverwort community across three stands

表5 相对优势度大于5%的藓类植物重要值(IV)Table5 Importance value(IV)of moss species with relative dominance≥5%across three stands

表6 3种林分藓类植物群落组成的多响应置换程序分析Table6 Multi-response permutation procedures(MRPP)for the variations in species composition of moss community across three stands
另外,从本研究中苔藓植物群落的优势种和次优种来看,3种森林群落的差异相对较少,主要表现为物种重要值排序的相对变化。在苔类植物群落中,双齿裂萼苔和细指苔重要值的排序变化体现出3种森林类型中苔类植物群落的差异;而3种森林类型中藓类植物群落均以东亚拟鳞叶藓为优势种,其差异主要表现在次优种上,这可能体现出森林所处位置的气候条件对苔藓植物群落的主导作用[19-21];但还需要进一步在更大尺度的研究来证实。另一方面,森林苔藓植物的分布受到各种生物因子和非生物因子的影响,非生物因子中的海拔、地形、土壤等因素均对森林苔藓植物的组成和分布产生显著的影响,而本文的研究仅是揭示了森林群落树种组成及林分类型作为生物因子与苔藓植物分布的关联。
