木麻黄海防林种子雨的时空动态
2019-08-08杨彬王玉郝清玉
杨彬,王玉,郝清玉
(热带岛屿生态学教育部重点实验室,海南师范大学生命科学学院,海口 571158)
森林更新是指以自然力或人为力重新形成森林的过程,而天然更新是森林更新的重要方式,也是森林资源再生产的一个重要手段。相比于传统的人工育林方式,天然更新具有育林成本低、成林周期短、对空间和地力的利用率高、环境稳定,生态平衡、幼树抗性、保存性高、保持水土,提高地力等优势[1-2],是实现森林恢复的重要途径[3]。为保证森林更新的进行,通常需要以下条件[4-5]:充足优质的种源、种子萌发能力强、适宜幼苗成长的环境,这些条件有任何一个出现问题都会造成更新的障碍[6]。种子是天然更新的保证,不同树种其种子往往存在巨大差异[7],其数量和质量都会显著影响更新。质量高、活性持续时间长的种子可以为土壤种子库提供充足的种源有利于更新的发生[8]。对于人工林而言,由于树林多是同一时间栽种,往往会有整片幼龄林或老龄林导致结子率不高种源数目成为更新障碍[9]。
种子雨是指成熟的种子在特定时间与空间内依靠自身重力及外界力量从母体上脱离到达地表的过程,植物天然更新的重要一环[10],直接影响了种子的扩散,进而对种子萌发、幼苗定居起着重要影响,最终影响着天然更新的质量[11]。种子雨往往存在着时空差异[12-13]。时间差异分为年际差异[14]和季节差异[11,15],前者指种子年际产量表现出显著差异,而不是在平均产量水平的波动[16],同时高纬度的差异往往大于低纬度地区[17];而降水、温度、光照等因子也会造成种子雨的季节差异,几乎所有对热带森林种子雨物候的研究也表明结实存在季节性[12]。种子扩散的空间过程会造成种子雨分布的空间差异,会影响种群乃至整个群落的空间分布[18]。
木麻黄(Casuarina equisetifolia)是原产于澳大利亚及太平洋诸岛的木麻黄科(Casuarinaceae)木麻黄属常绿乔木,高可达30 m。我国最早于1919年将其引种到福建泉州,20世纪50年代后,在东南沿海特别是雷州半岛和海南岛大面积栽种[19]。因其具有生长迅速、抗风能力强、不怕沙埋、耐盐碱等优良特性,成为我国南方沿海各省防风固沙的主要先锋树种[20]。海南省沿岸具有1 450 km宜林海岸线,自上世纪50年代我省开始有计划地引进木麻黄营造沿海防护林起至90年代,全岛海岸线除部分红树林(Mangrove)、椰子(Cocos nucifera)林、青皮(Vatica mangachapoi)林及马占相思(Acacia mangium)、大叶相思(A.auriculiformis)与木麻黄混交外,皆为木麻黄海防林纯林,成林面积达5.6×104hm2[21]。但以木麻黄为主的海防林也存在着诸多问题,如树种单一、结构简单、林分质量差、破坏严重等[22],此外木麻黄海防林的经营上还存在重要问题,即天然更新困难[23]。
目前国内外对木麻黄的研究多集中在化感作用、抗逆性、土壤养分及微生物方面,关于木麻黄种子雨分布特征及其萌发性能均未见报道,因此本文通过对木麻黄海防林种子雨进行调查研究,揭示海南岛海防林中木麻黄种子雨的时空特征及种子萌发的动态特性,了解木麻黄在海南岛的更新潜力,为进一步优化海南岛木麻黄海防林的建设提供理论基础。
1 材料和方法
1.1 研究区概况
研究区位于海南岛海口市灵山镇后尾村附近的木麻黄(Casuarina equisetifolia)海防林,该区为热带季风气候,年均气温24.8℃,年均降水量1 646 mm,年均相对湿度为82%,年日照时数1 878 h[24]。土壤为砂质土,养分较少。在研究区内共设置3块30 m×20 m的样地,样地之间相距约100 m,调查样地均属于中龄林或成熟林,其中3号样地因遭台风破坏,小老树较多。木麻黄平均密度为1 350.01 ind.hm-2,雌雄株为1∶1.5,平均胸径为7.64 cm,并伴有几株人工混交的大叶相思成树和椰子幼苗(表1)。另外,调查发现木麻黄主要是雌株有结实球果,但也有极个别雌雄同株有结实球果。
1.2 种子雨的收集
2017年7月中旬在样地中布置10个1 m×1 m的种子雨收集网,共30个。收集网行间距为5 m×10 m(图1)。种子雨收集网为尼龙网,直接铺设在木麻黄凋落物上,四角用一次性竹筷子固定。从2017年8月初至2018年4月中旬收集种子雨,持续9个月。由于种子雨持续时间较长,且考虑到种子雨萌发的持续时间,种子雨散布期内,每隔半个月收集1次,一般是月初和中旬,若有意外情况酌情提前或延后1~3 d,共收集18次。以种子雨收集网为单位,每次收集30份种子,将收集回的种子净种,并去除种胚不完整的种子,清点每个收集网的种子数量,并记录为有效种子数量。
1.3 种子萌发
每次收集种子雨后进行种子萌发试验,检测种子质量。将有效种子混匀并均分为4份,每份取25粒,共100粒为1个萌发样本。种子萌发采用纸间法,在室温下种子不经任何其他处理完成萌发试验。每日按时定量补水并统计萌发数量,一般持续20 d。每次收集回的种子做3个重复,每个重复100粒种子,以平均萌发率代表每批次种子的质量。
1.4 木麻黄球果数量调查
木麻黄球果调查是以每个种子雨收集网为中心,调查周围5 m内的木麻黄雌株或雌雄同株的株数,及每株的球果数量(图1)。由于木麻黄树木不高,球果明显,因此,球果数量调查采用地面观测计数法,对于结实量较多的株数,采用4分法,即调查树冠1/4部分的球果数量,然后乘以4。
1.5 数据处理和分析
种子雨密度(grain m-2)=收集有效种子数量/收集面积;种子雨年际密度(grain m-2a-1)为各次种子雨密度之和。选用变异系数表示数据离散程度,CV=式中,CV为变异系数,为平均数,S为标准差。收集网的种子雨相对密度=各网各次种子雨密度/各网年际总密度×100%。
种子雨空间分析采用方差/均值(Cx)、负二项指数(K)、聚集度指标(I)及 Cassie.R.M.指标(CA)判定。为均匀分布,Cx>1为聚集分布,Cx=1为随机分布;K<0为非聚集分布,K>0为聚集分布;为均匀分布,I>0为聚集分布,I=0为随机分布;CA=为均匀分布,CA>0为聚集分布,CA=0为随机分布。式中,为均值;S2为方差。
采用Microsoft Excel 2019及SPSS 19.0等软件对数据进行统计分析。采用单因素方差分析(One-Way ANOVA)对种子雨密度、萌发率等进行差异显著性分析,采用Duncan’s多重比较对不同散落时间种子雨的萌发率进行比较以P<0.05表示差异显著。采用Pearson相关系数对种子雨密度和附近球果数进行相关性分析。
2 结果和分析
2.1 种子雨的大小
木麻黄种子雨的年际总密度为1764.63grainm-2a-1;其中1号样地的最大(2 659.00 grain m-2a-1);3号样地最小(862.90 grain m-2a-1),其中,单网年际密度最小值为78 grain m-2a-1,发生在3号样地;单网最大年际密度为10 100 grain m-2a-1,发生在2号样地(表2)。方差分析表明,不同样地和同一样地不同收集网种子雨的差异极显著(P=0.002和0.000<0.010)。
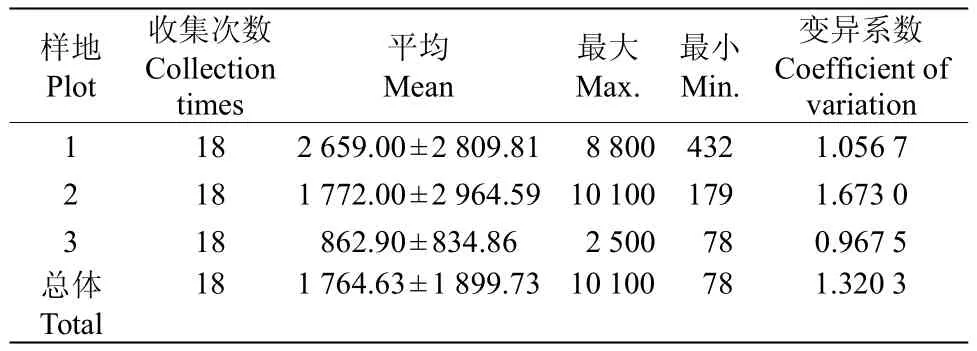
表2 不同样地的种子雨年际总密度(grain m-2a-1)Table2 Total interannual seed rain density(grain m-2a-1)in different plots
2.2 种子雨的时间分布特征
从8月开始,3个样地平均种子雨密度大体随时间缓慢增加,高峰期集中在12月上中旬,落种量占整个散落量的43.38%,此后,种子雨密度呈波动状缓慢减少,到次年4月中旬基本结束(图2)。在种子雨初期,1号样地的种子雨密度高达133.6 grain m-2,然后逐渐减少至9月初的51.3 grain m-2,此后逐步升高至峰值596.7 grain m-2;2号样地种子雨密度呈波动状缓慢增加,但大体均小于1号样地;3号样地与2号样地一样,其密度最小,均低于2号样地。
研究期间共计收集540网次,93%的收集网种子雨密度峰值出现在12月上中旬,其中21个收集网种子雨最大密度出现在12月初,占70%;7个出现在12月中旬,占23%;另有2个出现在次年2月中旬,仅占7%(图3)。另外,不同收集网种子雨密度峰值不同,其中最大峰值为3 762 grain m-2,最小为22 grain m-2,相差171倍。从图4可见,种子雨相对密度较大(>32.7%)的6个网次均在12月上中旬;37个网次的种子雨相对密度中等(16.4%~32.7%),其中12月份两次收集的约占81%;相对密度较小(<16.4%)的有497个网次,12月仅占4.8%。
在整个种子雨散播过程中,种子雨平均萌发率很低,仅为11.31%,其中萌发率较高的有4次,也仅为22.3%~30.0%;萌发率较低的有8次,为0~6.0%;萌发率中等的有6次,为9.0%~17.3%。不同时期种子雨的萌发率差异显著(P<0.05)(图5)。
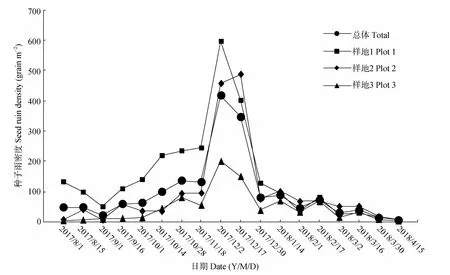
图2 种子雨密度的时间变化Fig.2 Temporal variation of seed rain density
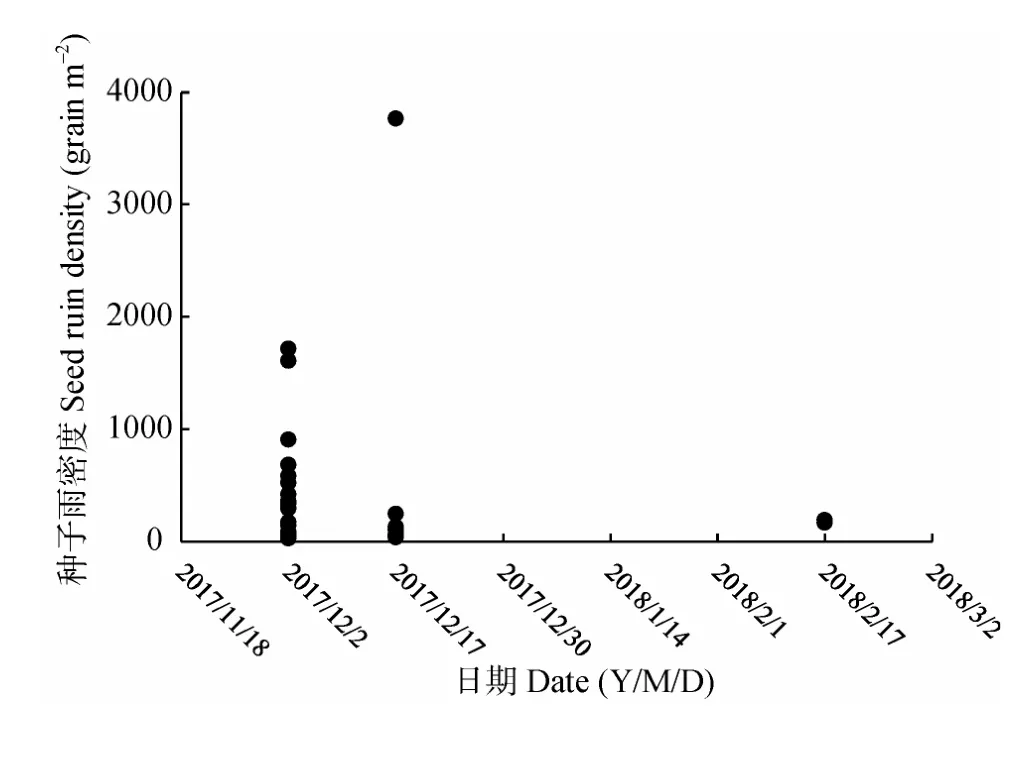
图3 各收集网种子雨密度峰值分布图Fig.3 Distribution of the peak density of each seed rain collection nets
2.3 种子雨的空间分布特征
种子雨密度只能提供种子雨分布的数量信息,种子雨的分布类型则能说明种子是如何在空间配置的。种子雨的分布类型主要分为均匀分布、聚集分布和随机分布(表1)。
从表3可见,3个样地和总体的方差/均值比CX>1,负二项指数K>0,聚集度指标I>0,Cassie.R.M.指标CA>0,4种方法均表明木麻黄种子雨呈聚集分布,且聚集程度很高(CX和I远远大于1)。
相关分析表明(图6),种子雨密度和附近球果数呈极显著正相关关系(r=0.530;P=0.003<0.01),表明木麻黄的种子雨主要散播在母树周围,同时也说明木麻黄种子虽然具有种翅,种子也较轻,但在木麻黄林分内扩散距离十分有限。相关分析结果与分布格局结论相一致,充分说明木麻黄种子雨呈聚集分布。
3 讨论和结论
3.1 种子雨的密度
植物种子雨密度大小是衡量其结实量和更新潜力的重要指标。不同植物因繁殖策略不同,结实量往往存在巨大差异,因此不同物种的种子雨密度也不尽相同[20]。调查样地木麻黄雄株比例高达60%,种子雨密度高达1 764.63 grain m-2a-1,远高于大部分物种种群,如川西南山地高山栲(Castanopsis delavayi,2.3~2.8 grain m-2)[25]、瑞典北方云杉(Picea asperata,86 grain m-2)[26]、准噶尔盆地梭梭(Haloxylon ammodendron,189 grain m-2)[13]、福建南方红豆杉(Taxus chinensis,252 grain m-2)[27],但小于福建杉木(Cunninghamia lanceolata)纯林(3 300 grain m-2)[11]。当然,海南岛不同分布区的木麻黄海防林,因龄级结构、密度、雌雄株比例不同,种子雨密度会有显著变化,但考虑到调查样地并非属于较好的林分结构,因此木麻黄成熟海防林的种子雨密度较大,种源数量充足,表明木麻黄天然更新的障碍因子并非是种源数量问题。

图4 收集网种子雨相对密度的热点图Fig.4 Hotspot map of seed rain relative density

图5 种子萌发率随时间的变化。柱上不同字母表示差异显著(P<0.05)。Fig.5 Temporal variation of seed germination rate.Different letters upon column indicate significant difference at 0.05 level.
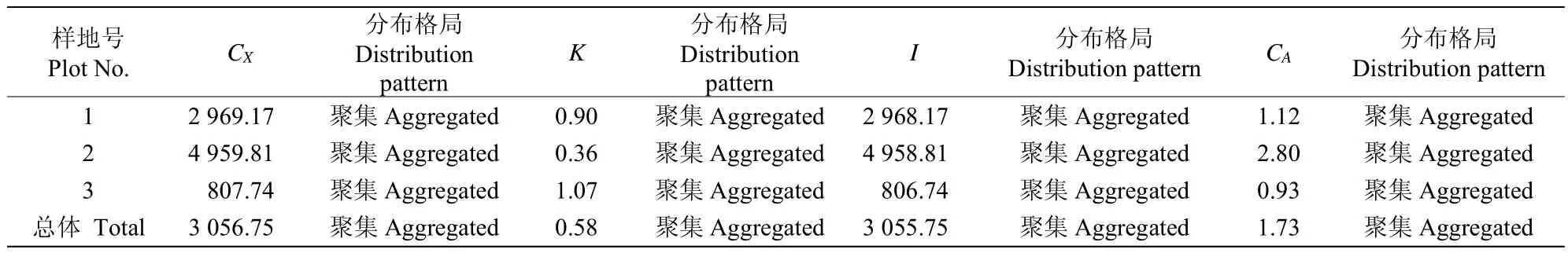
表3 种子雨空间分布类型Table3 Spatial distribution pattern of seed rain

图6 种子雨密度与球果数量的关系Fig.6 Relation between seed rain density and number of cones
3.2 种子雨的时间动态
种子雨在整个散落过程中随时间呈明显的异质性,且种子雨高峰期明显。木麻黄种子雨的时间动态变化规律与杉木[11]、油松[28]等树种相似,即在散落初期种子雨密度较低,在中期增大达到最大值,接着逐渐减少直至结束,出现1个高峰期。种子的落雨期一般为2~3个月,如黄土高原子午岭油松(Pinus tabuliformis)、川西南山地高山栲[25]和福建南方红豆杉[27],但木麻黄种子雨持续时间较长(9个月),且落雨期经历了海南雨季和旱季2个季节。这说明木麻黄种子雨散播期不会因旱季缺水妨碍其正常萌发。另外,93%收集网的种子雨密度峰值出现在12月上中旬,表明不同树木的落雨期基本同步,即木麻黄林分落雨期较长并非是不同树木落雨期不同步造成的。
3.3 种子雨活力的时间动态
在整个落雨期内,一般种子的萌发率随成熟度增加而增加,种子雨密度达到峰值时的萌发率最高,之后递减,如准噶尔盆地的梭梭[13],但木麻黄种子雨的活力并不服从这个规律,呈波动状态。这与种子的成熟度关系不大,因为种子雨散布初期的8月和高峰期的12月种子萌发率并无显著差异,说明种子雨散播初期种子成熟度已基本与峰期的相同;可能与种子雨在林内的老化程度有关。虽然木麻黄种子雨平均萌发率(11.31%)远低于准噶尔盆地梭梭(80%)[13]、川西亚高山人工云杉(71%)[29]、辽东山区长白落叶松(Larix olgensis,约30%)[30],但从树上采集的同期木麻黄种子萌发率为58.43%,远高于木麻黄种子雨的最高萌发率(30%)。这说明由于种子雨收集的间隔期为半个月,部分先掉落的种子在高温、高湿、多雨等环境下导致种子雨老化。这表明,种子雨的萌发率不仅取决于种子本身的萌发能力,还与种子的抗老化能力、生境的环境条件、种子雨收集间隔期长短等有关。
虽然种子雨的萌发率较低,但由于种子雨密度较大,所以具有萌发潜力的种子数量仍高达199.58 grain m-2a-1,说明种源质量也不是制约木麻黄天然更新困难的主要因子。
3.4 种子雨的空间分布格局
种子雨的散布往往存在空间异质性,而且其构成的空间格局会影响种群更新格局,进而影响种群分布[18]。大部分种子在空间格局上表现为种子雨分布在母树周围,并随距母树距离的增加散落量逐渐减少[31]。经4种空间分布类型评价方法判定,木麻黄种子雨均呈聚集分布,并且种子雨密度与周围母树结实量呈极显著的正相关(r=0.530)。种子雨的散落距离往往受种子重力、种子形态、地形、株高及风力等影响[32]。木麻黄种子小而轻,具有种翅,非常适合远距离散播,但研究结果表明,木麻黄种子雨呈极强的聚集分布,说明木麻黄林内,由于受到树冠的阻碍作用,木麻黄种子的扩散能力受到一定的限制。
