基于密码子优化的FAD依赖葡萄糖脱氢酶在毕赤酵母中的高效表达及酶学性质
2019-07-29董聪高庆华王玥罗同阳
董聪 高庆华 王玥 罗同阳
(河北省科学院微生物研究所,保定 071051)
FAD 依赖的葡萄糖脱氢酶(FAD-dependent glucose dehydrogenase,简称 FAD-GDH,EC 1.1.99.10),同葡萄糖氧化酶、吡喃糖脱氢酶、胆碱脱氢酶和甲醇氧化酶一样都属于 GMC 氧化还原酶(Glucosemethanol-choline-oxidoreductase) 家 族[1]。 它 是 以FAD 为辅基能够在 NAD(P)+等存在的情况下催化β-D-葡萄糖生成 D-葡萄糖酸-δ-内酯,而 D-葡萄糖酸-δ-内酯会自发形成葡萄糖酸。
研究表明,FAD-GDH是一类重要的临床检测和工业用酶,可应用于血糖试纸或葡萄糖生物传感器的制备、辅酶的再生、新型燃料电池[2]和心脏起搏器[3]的研制等。尤其是可以作为血糖检测用酶,FAD-GDH不以氧气作为酶促反应的电子受体,检测结果更准确,且具有转化率高、底物特异性好等优点,因此有很大的应用前景。
虽然到目前为止已经有不少FAD-GDH被分离和鉴定,但是针对特定FAD-GDH全面深入的研究很少,主要原因是FAD-GDH很难实现可溶的重组表达,在大肠杆菌进行重组表达时形成包涵体,不易分离纯化。重组表达是克服这一障碍的有效方法。在真核表达系统,毕赤酵母中有过实现可溶表达,得到有活性的FAD-GDH的报道[4]。毕赤酵母(P. pastoris)表达系统在进行外源基因表达时,具有生长周期短、可进行胞外分泌表达、目的蛋白易于纯化和易于高密度发酵培养等优点[5-7]。由于毕赤酵母对密码子的偏爱性,目的基因的密码子序列对产物的表达量也有很大影响。大量研究表明,密码子优化对提高产物表达量有明显的作用[8-13]。因此,如果想提高产物的产量,不但要优化表达条件,还需要对密码子进行优化,以期为FAD依赖的葡萄糖脱氢酶进一步扩大生产提供理论和技术支持,使其更好的应用于血糖检测及其他相关领域[14]。
本研究以NCBI上登录号为XM_001216916的土曲霉NIH2624基因为基础,根据毕赤酵母的密码子偏爱性对该基因的密码子进行优化,然后委托公司进行基因合成。接着将该基因克隆到毕赤酵母中,并在基因的3′端接上6个组氨酸标签的核苷酸序列,以便于重组表达蛋白的纯化。经过筛选和优化,得到了高产该FAD-GDH的重组菌株。采用该菌株摸索发酵条件,实现了该蛋白的高效表达,并对其酶学性质进行了研究。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 FAD依赖葡萄糖脱氢酶基因由安徽通用生物公司合成,合成的基因连接在pUC57T载体上;大肠杆菌DH5α购自上海生工;酵母表达菌株X33由中科院微生物研究所提供;质粒pMD由本实验室前期构建。
1.1.2 酶和主要试剂EcoR I、NotI和SacI限制性内切酶为NEB公司产品;T4 DNA连接酶为Thermo公司产品;DNA Marker、DNA凝胶回收试剂盒和质粒DNA小提试剂盒为天根生化科技有限公司产品;G418为Sigma公司产品;2,6-二氯靛酚钠(DCIP)为BBL Life Sciences公司产品,BeyoGoldTMHis-tag Purification Resin为碧云天生物科技有限公司产品;其他试剂均为国产分析纯试剂。
1.1.3 培 养 基 LB、YPD、YPDS、YPCS、BMGY和基础盐培养基等培养基配方和X33培养条件参照文献[14]。
1.2 方法
1.2.1 葡萄糖脱氢酶密码子优化及人工合成 以NCBI 上登录号为 XM_001216916 的土曲霉 NIH2624基因为基础,根据毕赤酵母的密码子偏爱性对该基因的密码子进行优化,利用密码子优化软件http://www.jcat.de/分析,发现蛋白酶基因中有多处是毕赤酵母的稀有密码子,利用密码子优化软件http://www.jcat.de/,对蛋白酶基因进行密码子优化,在不改变氨基酸序列的前提下,获得优化后的蛋白酶的基因序列。利用信号肽预测服务器SignalP,服务器网址http://www.cbs.dtu.dk/services/SignalP/,预测信号肽,然后去除,在序列的5′ 端加上EcoR I酶切位点,3′端加上6个组氨酸标签后再加NotI酶切位点。然后将优化构建的基因序列送安徽通用生物公司合成,合成的基因连接在pUC57T载体上。
1.2.2 重组表达载体的构建 人工合成的pUC57-GDH和载体pMD经EcoR I和NotI双酶切后切胶回收纯化,然后用T4 DNA连接酶连接,转化大肠杆菌DH5α感受态细胞中,在含氨苄的LB平板上筛选阳性克隆,对阳性克隆提取质粒进一步进行酶切验证,由安徽通用生物公司进行测序分析。
1.2.3 阳性转化子的获得 构建好的重组表达载体pMD-GDH经SacI线性化后电转表达宿主P. pastorisX33感受态细胞,电转条件:4 Kv,3 ms,然后快速加入1 mL 预冷YPDS液体培养基,于30℃培养箱静置培养2-6 h,涂布在终浓度为250 μg/mL G418的YPD平板上,3 d后获得阳性转化子。
1.2.4 重组蛋白的诱导表达 将重组菌株X33/pMDGDH挑取单菌落接种于5 mL YPCS试管培养基中,14-18 h后加 1%(V/V)甲醇,之后24 h和48 h后均各加 1%(V/V)甲醇诱导,72 h收菌,8 000 r/min离心 5 min收集上清,测定酶活。至少重复3次。
1.2.5 FAD依赖葡萄糖脱氢酶的酶活测定 FAD依赖葡萄糖脱氢酶的酶活测定方法参照文献[4],使用DCIP作为电子受体,在30℃反应180 s,测定520nm的吸光值变化。
1.2.6 FAD依赖葡萄糖脱氢酶酶学性质分析 酶的最适作用温度和pH 以及热稳定性、pH稳定性测定、金属离子的影响及底物特异性参照文献[14]。所用的缓冲液均为pH 5.2的磷酸盐缓冲液。
1.2.7 10 L发酵罐放大表达FAD-GDH 选取试管水平上FAD依赖的葡萄糖脱氢酶活性最高的转化子,在10 L 发酵罐条件下进行FAD-GDH 的表达。挑单克隆接种于YPD培养基培养10 h,按3%接种量转接于 100 mL BMGY 培养基至OD600≈6 接种于10 L发酵罐,初期装液量为 5.6 L BSM 培养基,高压灭菌20 min 后,以 10%发酵体积接种。期间每隔24h加维生素C水溶液(终浓度80 μg/L),28℃培养,每12 h取样测定发酵液酶活;上清液进行10% 聚丙烯酰胺凝胶电泳,观察FAD-GDH 蛋白质的表达。具体方法参照文献[15]。
2 结果
2.1 FAD依赖葡萄糖脱氢酶密码子优化
主要优化指标为CAI、密码子使用相对分布、GC含量、酶切位点等,在不改变氨基酸序列的前提下,在这段优化的密码子序列中,共替换了416个碱基,优化后的密码子序列几乎全部是毕赤酵母的偏爱密码子,为目的基因在毕赤酵母中成功表达提供了保障。基因序列如图1 所示。
2.2 酵母表达载体pMD-GDH的构建
将人工合成包含目的基因的载体和pMD载体分别利用EcoR I和NotI双酶切,并且利用琼脂糖凝胶回收试剂盒回收目的片段和表达载体片段,然后利用T4 DNA连接酶将目的片段连接到表达载体上,并转化大肠杆菌DH5α中,通过提取质粒,EcoR I/NotI双酶切鉴定质粒的正确性(图2)。有 1.7 kb左右的条带,说明FAD-GDH基因连接到了pMD载体上,然后将pMD-GDH质粒送华大基因测序,测序结果说明目的基因已连接到载体上,且序列正确。
2.3 阳性转化子的获得
将酶切正确的重组质粒pMD-GDH进行SacI线性化后电转表达宿主Pichia pastorisX33,构建重组菌X33/pMD-GDH,涂布在终浓度为250 μg/mL G418的YPD平板上,3 d后获得阳性转化子(图3)。
将重组菌株X33/pMD-GDH挑取单菌落接种于5 mL YPCS试管培养基中,14-18 h后加 1%(V/V)甲醇,之后24 h和48 h后均各加 1%(V/V)甲醇诱导,96 h收菌,8 000 r/min离心 5 min收集上清,上清液利用DCIP的方法测定 FAD 依赖葡萄糖脱氢酶的活力,酶活定义为反应体系中每分钟反应1 μmol电子受体所需的酶量,并以此来确定重组子是否为阳性。共挑取了18个单菌落诱导培养,测定酶活,重复3次,其中9号酶活较高且稳定(图4),试管中酶活为20.85 U/mL,选其做进一步的研究。
2.4 SDS-PAGE电泳分析
将在试管中诱导表达收集的酶液经超滤浓缩后进行SDS-PAGE电泳鉴定,结果如图5所示,重组毕赤酵母在60-70 kD之间有一条条带,与理论分子量相符,而pMD空载转化的酵母没有类似的条带。并且用DCIP法测定发酵液有活性,说明FAD依赖的葡萄糖脱氢酶在酵母体内实现了分泌表达。该基因3′端有His-tag序列,可通过与镍离子的亲和层析进行纯化,经BeyoGoldTMHis-tag Purification Resin纯化后SDS-PAGE分析结果如图6所示。
2.5 FAD依赖的葡萄糖脱氢酶酶学性质分析
酶学性质分析结果如图7-图9所示,毕赤酵母表达的FAD-GDH最适反应温度为55℃,在50℃下处理150 min仍有70%的活性。FAD-GDH的最适pH值为7.0,在 pH 4-7 范围内,37℃保温4 h,FAD-GDH 仍能保持 50%以上的活性。金属离子Cu2+对酶活抑制作用比较大。FAD-GDH的底物专一性较好,以葡萄糖为最适底物。当以木糖为底物时,其相对活性是以葡萄糖为底物的36%;对麦芽糖和海藻糖也有一定活性,对其他单糖或二糖不显示催化活性。
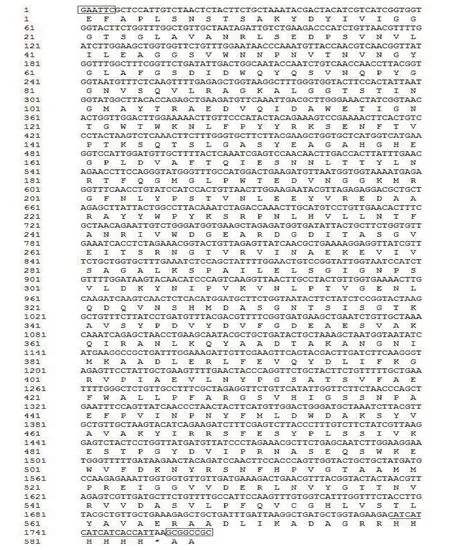
图1 优化后的FAD-GDH基因序列
2.6 10 L发酵罐扩大培养
重组菌P. pastorispMD-GDH在10 L发酵罐的放大培养,在菌体生长阶段当溶氧DO值在20 h迅速上升(培养基甘油耗尽)时以恒速补加甘油4 h,饥饿培养0.5 h 后开始甲醇诱导。在甲醇诱导阶段,通过控制DO 值来流加甲醇以高效诱导重组毕赤酵母产酶,同时可避免过高的甲醇对菌体产生毒害作用,甲醇流加[6 mL/(h·L)]与DO 偶联,即DO 值高于0%时补加甲醇,维持DO 0%左右。随着甲醇添加,FAD依赖葡萄糖脱氢酶酶活不断提升,甲醇诱导136 h时酶活达到257.6 U/mL(图10)。此外,对不同时间点的发酵上清液进行SDS-PAGE 电泳分析,结果显示各上清液在SDS-PAGE 图中有一条清晰条带,该条带随发酵时间增加而变得浓重清晰,表明发酵上清液中FAD依赖的葡萄糖脱氢酶含量随着发酵时间增加而显著提高,与酶活增长趋势保持一致。
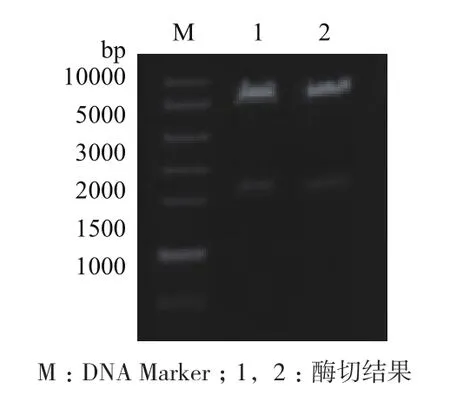
图2 pMD-GDH载体双酶切鉴定

图3 X33/pMD-GDH单菌落
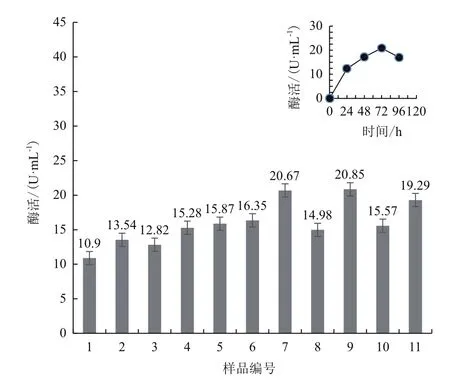
图4 高产FAD依赖葡萄糖脱氢酶重组毕赤酵母的试管筛选

图5 重组蛋白的SDS-PAGE检测结果

图6 重组蛋白纯化结果

图7 FAD-GDH 酶学性质

图8 不同金属离子对 FAD-GDH 的影响

图9 FAD-GDH 的底物特异性
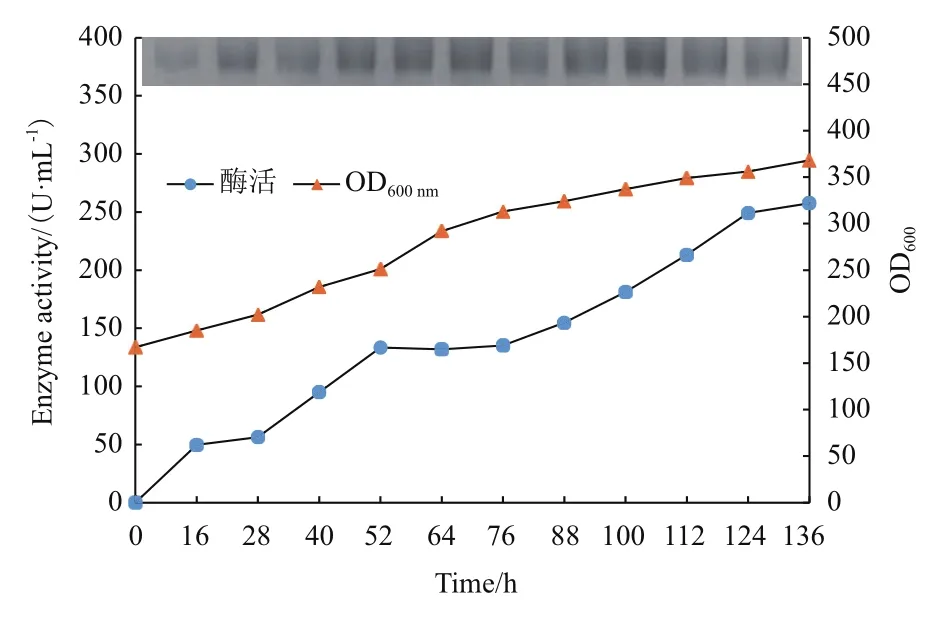
图10 上罐发酵实验
3 讨论
FAD依赖的葡萄糖脱氢酶作为血糖检测用酶有很大的应用前景,但是在天然宿主菌中的表达量很低,在大肠杆菌进行重组表达时形成包涵体,表达量低,不易分离[16],给FAD依赖的葡萄糖脱氢酶的工业应用带来一定的限制,所以利用基因工程技术实现FAD依赖的葡萄糖脱氢酶的高效表达成了目前研究的重点。酵母表达系统作为一种真核表达系统,与原核表达系统相比,具有很多优势,如生长速度快,可使用高效的AOX 启动子以及可实现异源蛋白的分泌表达等优点,广泛用于各种具有工业用途的重组蛋白的生产[58,17]。
本实验利用酵母表达系统表达FAD依赖葡萄糖脱氢酶,通过分析影响表达量的因素,根据毕赤酵母对密码子的偏爱性对该基因序列进行了优化[18],提高其表达量。蛋白酶传统分离纯化方法多采用超滤-阳离子柱-超滤-阴离子柱等步骤,工艺复杂,收率低。本研究在FAD依赖葡萄糖脱氢酶基因3′端加上了6 个组氨酸标签,可以通过亲和层析的方法进行纯化,效率高,适合大规模纯化。
在已有的报道中,杉木炭疽病菌来源的 FADGDH 在毕赤酵母中得到了较好的表达,通过采用 7 L 的发酵罐,发酵约 50.5 h,产量达到 48 000 U/L报道[14]。本实验利用酵母表达系统表达FAD依赖的葡萄糖脱氢酶,通过优化FAD依赖的葡萄糖脱氢酶密码子序列,在10 L发酵罐培养时经过136 h诱导培养,酶活达到257 600 U/L,是现有技术中报道的酶活的5.3倍。毕赤酵母可通过高密度发酵培养大量生产重组蛋白,在提高产量的同时有利于简化纯化步骤,降低生产成本。
酶学性质是表征一个酶的基本参数,通过酶学性质的测定和评估,可以了解一个酶的特有性质,为酶的应用和改造提供一系列必要的初始参数。在血糖检测用酶中,要求所用的酶有较高的催化效率,较窄的底物谱,良好的热稳定性[19]。周利伟等[14]报道的重组FAD-GDH最适温度为40℃,本研究毕赤酵母重组表达FAD-GDH的最适反应温度为55℃,且热稳定性良好,在50℃下处理150 min仍有70%的活性;其次该酶的最适pH值为7.0,在 pH4-7 范围内,37℃保温4 h,FAD-GDH 仍能保持 50%以上的活性,有较高的应用价值。金属离子Cu2+对该酶活抑制作用比较大,与前人研究结果一致。FADGDH的底物专一性较好,以葡萄糖为最适底物。当以木糖为底物时,其相对活性是以葡萄糖为底物的36%;对麦芽糖和海藻糖也有一定活性,与杨愈丰等[20]研究结果相符,对其他单糖或二糖不显示催化活性。
4 结论
本研究以 NCBI 上登录号为 XM_001216916 的土曲霉 NIH2624 基因为基础,根据毕赤酵母的密码子偏爱性对该基因的密码子进行优化,将该基因克隆到毕赤酵母中,经过筛选和优化,得到了高产FAD-GDH 的重组菌株。试管水平酶活达到20 850 U/L;在10 L发酵罐培养时经过136 h诱导培养,酶活达到257 600 U/L,是现有报道中酶活的5.3倍。然后研究了其酶学性质,pH、温度稳定性和底物专一性较好。
