曹妃甸近岸海域大型底栖动物群落特征
2019-07-23李德鹏柳圭泽陈兆林于大涛
梁 淼,姜 倩,李德鹏,柳圭泽,陈兆林,崔 雷,2,于大涛
( 1.国家海洋环境监测中心,辽宁 大连 116023; 2.天津大学 建筑工程学院,天津 300072 )
由于底栖动物运动能力差、生命周期长、可快速响应海洋环境污染压力的特点[1],常被用作监测和评估海洋生态环境质量健康的指示生物[2]。底栖动物群落结构的变化可以指示其栖息环境受扰动的状况[3],海洋生态系统健康与否也将直接影响其空间分布和群落结构的变化[4]。近年来,对于近岸海域生态环境健康的评价方法较多,其中建立在大型底栖动物群落基础上的生物指数法已经成为其监测和评价的重要方法之一[5]。丰度/生物量比较曲线[6]、建立在大型底栖动物生态分组基础上的AZTI海洋生物指数(AMBI)和融入了多因子分析的AZTI海洋生物指数(M-AMBI)在近岸海域的生态状况评价和底栖群落结构健康评价中得到了广泛的应用[7-11]。
曹妃甸海域位于渤海湾东北部,属于半封闭内海,海水的交换能力较弱。近年来,随着该区域内的填海造陆、港口开发、工业区建设和矿产开采等一系列开发活动的进行,给海洋生态环境带来了巨大压力,生物资源和环境质量也面临着前所未有的考验[12]。因此,如何科学、准确、有效地评价曹妃甸近岸海域生态系统健康具有重要的生态意义。笔者利用2015年9月和2016年5月春、秋两季的大型底栖动物现状调查数据,阐述海域大型底栖动物群落结构特征和生物多样性的时空分布,并利用丰度/生物量比较曲线法、AMBI指数法和M-AMBI指数法对研究海域大型底栖动物群落健康和生境质量进行评价,以期为曹妃甸近岸海域生态系统健康评价及生态恢复提供有效的科学管理决策。
1 材料与方法
1.1 采样点设置
本研究共布设5个断面20个调查站位,于2015年9月和2016年5月对曹妃甸近岸海域进行大型底栖动物样品的采集。根据曹妃甸海域特征及开发利用现状,将调查区域分为3个区,分别为曹妃甸填海区东侧(调查断面5,共计4个站位)、西侧(调查断面1,共计4个站位)及填海区附近(调查断面2、3、4,共计12个站位)(图1)。
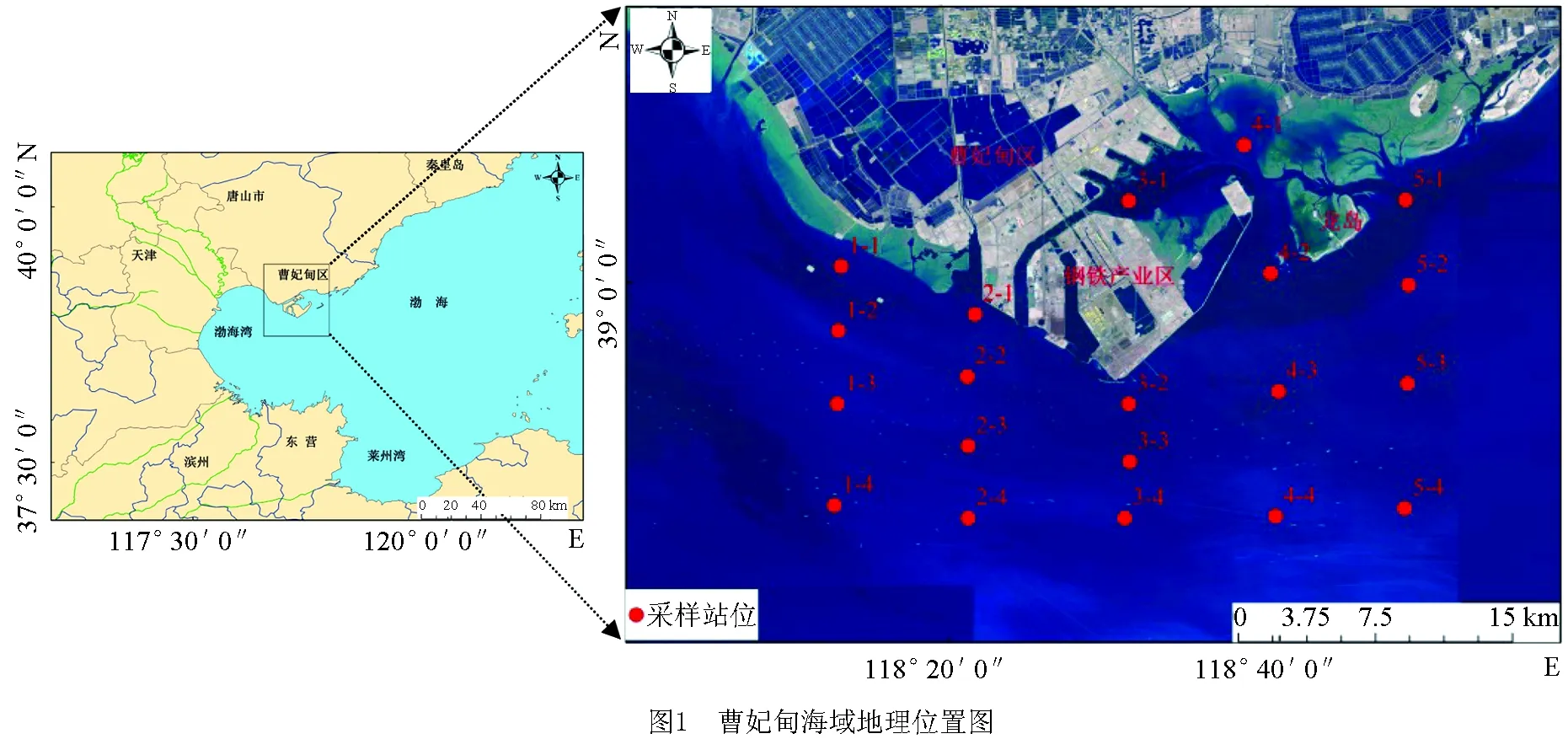
图1 曹妃甸近岸海域大型底栖动物采样站位示意
1.2 样品的采集和处理
大型底栖动物的采集和处理按照《海洋调查规范》(GB/T 12763.6—2007)[13]进行。泥样采用0.05 m2曙光振动式采泥器采集,每站取样5次,取样面积为0.2 m2,取样深度为10~20 cm。将采集到的沉积物样倒入网目为0.5 mm孔径的网筛中,用水冲洗掉底泥。拣出所有样品,装入样品瓶内,放入标签,用5%福尔马林固定液固定,样品带回实验室分析(包括种类鉴定、称量及计算等)。底栖动物种类鉴定依据《中国近海多毛类环节动物》[14]和中国动物志系列丛书等资料进行。物种鉴定及名录登记均通过《中国海洋生物名录》[15]进行校对和复核。
1.3 数据分析
1.3.1 群落特征分析
大型底栖动物群落特征分析主要通过优势度指数(Y)[16]、Shannon-Weaner多样性指数(H′)[17]、Pielou均匀度指数(J)[18]及相对重要性指数(IRI)[19]进行分析,其公式如下:
IRI=(M+N)×F×104
式中,n为该海域出现所有大型底栖动物种类的总个体数,ni为第i种大型底栖动物的个体数,fi为第i种大型底栖动物在各站位出现的频率。当Y>0.02时,即为该海域底栖动物优势种[16];Pi为第i种的个体数(ni)与样品中总个体数(N)的比值(ni/N),log2S为多样性指数的最大值,S为样品中的物种总数;M为某一种类的生物量占总生物量的百分比,N为某一种类的丰度占总丰度的百分比,F为该种类的出现频率。
1.3.2 群落结构分析
利用PRIMER 6.0软件进行大型底栖动物群落结构分析,对大型底栖动物的物种丰度进行log(x+1)转换后,计算站位间的Bray-Curtis相似性系数,构建相似性矩阵,并采用等级聚类方法和非度量多维尺度分析大型底栖动物群落的空间分布,通过双因素相似性分析方法分析不同聚类组间的差异性,并通过相似性百分比分析不同物种对各群落组内相似性和组间差异性的平均贡献率[20]。
1.3.3 底栖生境质量评价
底栖生境质量评价方法采用丰度/生物量比较曲线法、AMBI指数法和M-AMBI指数法。
丰度/生物量比较曲线法监测环境污染对底栖动物群落结构的扰动[6],是将生物量和丰度的k-优势度曲线绘入一张图中,当生物量的k-优势度曲线始终位于丰度k-优势度曲线之上,则群落未受扰动,优势种类执行k-选择繁殖策略;当丰度和生物量曲线接近重合或者出现部分交叉,则群落受到中等程度的污染扰动;当丰度k-优势度曲线位于生物量k-优势度曲线之上,则群落受到严重污染扰动,优势种类执行r-选择繁殖策略[6]。
AMBI指数和M-AMBI指数应用AMBI 5.0软件进行计算。首先,对所有底栖动物的命名根据《中国海洋生物名录》[15]进行核实,然后根据2017年6月的生物分组表进行底栖动物生态分组(http:∥ambi.azti.es),其中,EGⅠ为对扰动敏感的物种,EGⅡ为对扰动不敏感的物种,EGⅢ为对扰动有耐受力的物种,EGⅣ为第二级机会种,EGⅤ为第一级机会种。本次春、秋两季调查的物种矮指蚕(Pedinosomacurtum)和吻蛰虫(Genusartacama)未分组(它们不属于AMBI物种目录中的任何一种)。秋季,未分组的矮指蚕出现在4-4站位、吻蛰虫出现在1-3和5-4站位;春季,未分组的矮指蚕出现在2-4和5-3站位,并且它们在各站位中所占的比例均低于10%,不影响M-AMBI指数的计算结果。
M-AMBI是建立在AMBI指数基础上,结合底栖群落AMBI指数、Shannon-Weaner多样性指数和物种种数3项指标评分得出的多变量生物指数[2]。本研究数据处理中的参数设置按照欧盟对生态环境质量的定义,“差”生态质量的参照状态值选自于无生命沉积物下的群落结构参数和AMBI值[21],因此,多样性指数、物种数参照值取0,AMBI的参照值取6;“好”生物质量的参照状态值选取2006—2016年近10年研究区底栖动物物种数和Shannon-Weaner多样性指数的最高值以及AMBI指数的最低值,即春季物种数为13、多样性指数值为3.44;秋季物种种数为13、多样性指数值为3.40,AMBI指数均为0。AMBI和M-AMBI指数分级、扰动等级以及生态环境质量状况等级标准见表1[10]。
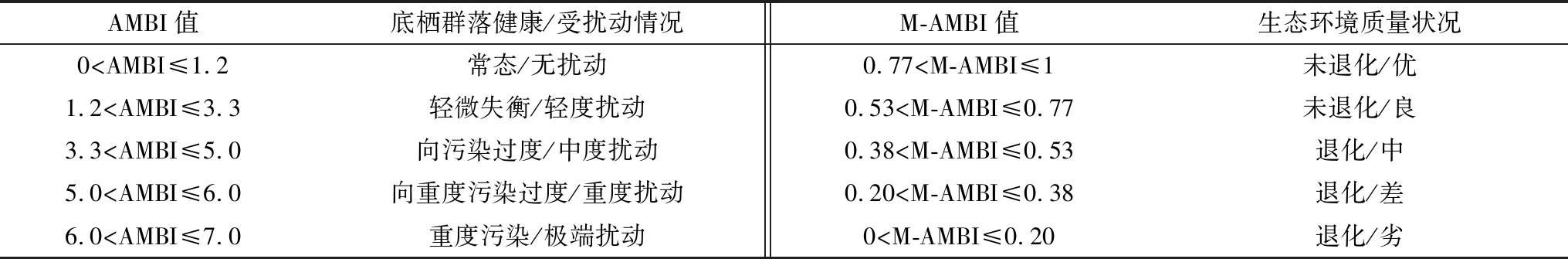
表1 AMBI和M-AMBI指数分级、扰动等级以及生态环境质量状况分级标准
1.4 数据统计分析
利用SPSS 13.0软件在对变量进行方差齐性检验(Levene检验)后,再对春、秋两季的底栖动物群落参数(种类数、丰度、生物量、多样性指数、均匀度指数、AMBI指数和M-AMBI指数)进行独立样本t检验;图件绘制使用ArcGis 10.2制作完成。
2 结 果
2.1 大型底栖动物群落特征分析
2.1.1 物种组成
2015年9月秋季,曹妃甸近岸海域共发现大型底栖动物5门50种。其中环节动物28种,占56.00%;软体动物4种,占8.00%;节肢动物16种,占32.00%;棘皮动物1种,占2.00%;纽形动物1种,占2.00%。2016年5月春季,调查海域共发现大型底栖动物6门62种。其中环节动物34种,占54.84%;软体动物6种,占9.68%;节肢动物17种,占27.42%;棘皮动物3种,占4.84%;纽形动物、腔肠动物各1种,占1.61%。春、秋两季调查海域种类分布主要为个体较小的环节动物、节肢动物,且物种种类数差异不显著(t=0.718,P=0.477>0.05)。
2.1.2 优势种分布特征
2015年9月秋季,曹妃甸近岸海域大型底栖动物优势种有4种,其中日本大螯蜚(Grandidierellajaponica)的优势度(0.090)和平均丰度(70.00 个/m2)最高,相对重要性指数较高(909.31),但其平均生物量最低,为0.03 g/m2,出现频率较低,为45.00%,生态分组属于EGⅢ对扰动有耐受力的物种;日本刺沙蚕(Neanthesjaponica)的优势度略低于日本大螯蜚,为0.075,但其相对重要性指数、平均生物量及出现频率均最高,分别为1414.91、1.41 g/m2、60.00%,其生态分组也为EGⅢ,为对扰动有耐受力的物种。2016年5月春季,仅出现一种优势种,为囊叶齿吻沙蚕(Nephtyscaeca),优势度为0.026,相对重要性指数为291.68,均低于秋季囊叶齿吻沙蚕的优势度(0.047)和相对重要性指数(574.37)。
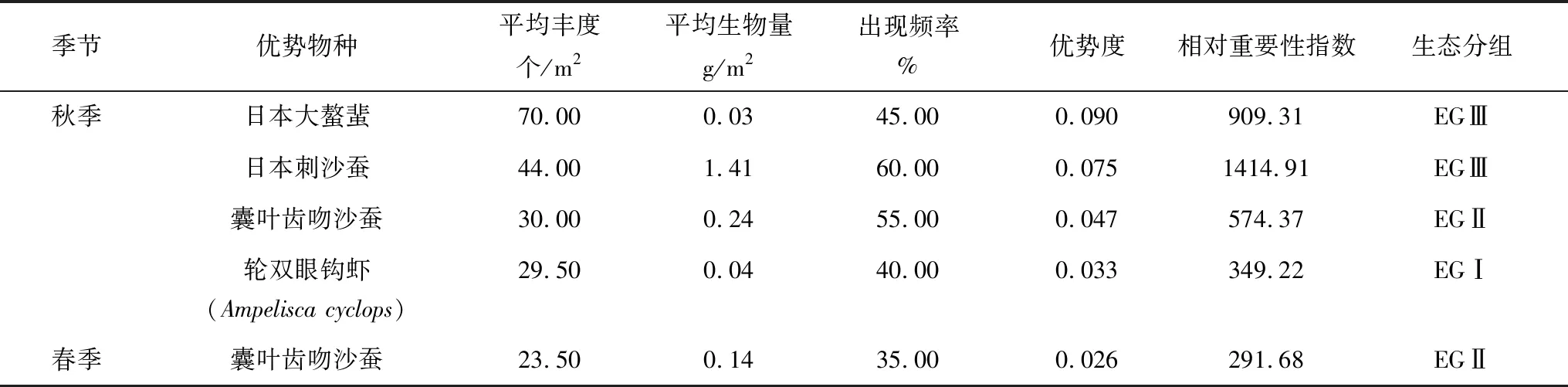
表2 曹妃甸近岸海域大型底栖动物优势种统计情况
2.1.3 丰度及生物量特征
曹妃甸近岸海域2015年9月和2016年5月大型底栖动物平均丰度分别为(350.50±396.50)个/m2和(317.50±190.80)个/m2,且平均丰度在春、秋两季无显著差异(t=0.327,P=0.746>0.05)。其中,2015年9月秋季,从丰度占比来看,贡献率最高的为节肢动物,其总丰度为3680.00个/m2,占比52.50%,其次为环节动物2970.00个/m2,占比35.84%,棘皮动物260.00个/m2(3.71%),软体动物80.00个/m2(1.14%)和纽形动物20.00个/m2(0.25%)(表3);从空间分布来看,大型底栖动物丰度最大值出现在4-1站位(1990个/m2),最低值出现在1-1站位(30.00个/m2),底栖动物丰度高值区主要集中在曹妃甸填海区东侧龙岛附近海域,低值区主要在曹妃甸填海区西侧调查断面1区(图2)。2016年5月春季,从丰度占比来看,贡献率最高的为环节动物,其总丰度为3160.00个/m2,占比50.00%,其次为节肢动物2590.00个/m2,占比40.90%,软体动物430.00个/m2(6.81%),棘皮动物100.00个/m2(1.58%),纽形动物30.00个/m2(0.47%)和腔肠动物10.00个/m2(0.24%)(表4);从空间分布来看,大型底栖动物丰度最大值出现在4-3站位(870.00个/m2),最低值出现在1-1站位(60.00个/m2),底栖动物丰度高值区主要集中在龙岛南侧海域,低值区分布与秋季一致,也为曹妃甸填海区西侧调查断面1区(图2)。秋季的最高值出现在4-1站位,而到春季4-1站位的丰度值显著降低,仅为70.00个/m2。
曹妃甸近岸海域2015年9月和2016年5月大型底栖动物平均生物量分别为(12.70±21.70) g/m2和(16.20±23.30) g/m2,且平均生物量在春、秋两季无显著差异(t=-0.470,P=0.641>0.05)。其中,2015年9月秋季,从生物量占比来看,贡献率最高的为节肢动物,其总生物量为88.30 g/m2,占比34.65%,其次为软体动物84.40 g/m2,占比33.12%,环节动物70.30 g/m2(27.59%),棘皮动物9.40 g/m2(3.69%)和纽形动物2.40 g/m2(0.95%)(表3);从空间分布来看,大型底栖动物生物量最大值出现在1-1站位(79.50 g/m2),最低值出现在3-1站位(1.00 g/m2),底栖动物生物量空间分布与其丰度分布呈相反趋势,即生物量高值区主要集中在曹妃甸填海区西侧调查断面1区,低值区主要在曹妃甸填海区东侧龙岛附近海域(图2)。2016年5月春季,从生物量占比来看,贡献率最高的为软体动物,其总生物量为136.60 g/m2,占比42.25%,其次为环节动物80.20 g/m2,占比24.81%,节肢动物75.50 g/m2(23.35%),棘皮动物15.90 g/m2(4.92%),腔肠动物13.30 个/m2(4.11%)和纽形动物1.80 个/m2(0.56%)(表4);从空间分布来看,大型底栖动物生物量最大值出现在2-2站位(103.90 g/m2),最低值出现在1-3站位(1.20 g/m2),底栖动物生物量空间分布也与其丰度分布成相反趋势(图2)。
2.1.4 生物多样性特征
曹妃甸调查海域2015年9月秋季的大型底栖动物多样性指数、均匀度指数均高于2016年5月春季,但其差异不显著(P>0.05)。其中,秋季,调查区内多样性指数平均值为2.44±0.54,最低值出现在1-1站位(0.92),最高值出现在2-2站位(3.25);均匀度指数平均值为0.87±0.06,最低值出现在4-1号站(0.72),最高值在1-2号站(0.97)。春季,调查区内多样性指数平均值为2.24±0.14,最低值出现在1-3站位(0.42),最高值出现在2-4站位(3.44);均匀度指数平均值为0.84±0.14,最小值出现在1-3号站(0.41),最高值在5-2号站(0.98)(图3,表5)。
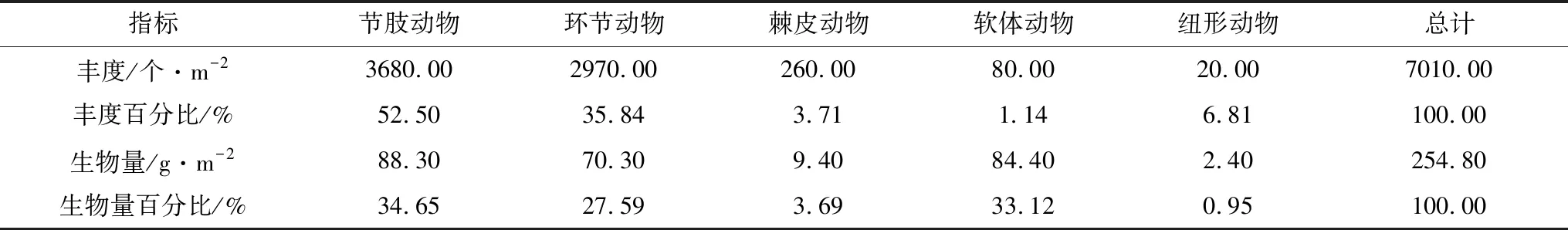
表3 曹妃甸近岸海域2015年9月大型底栖动物各类群的栖息丰度与生物量
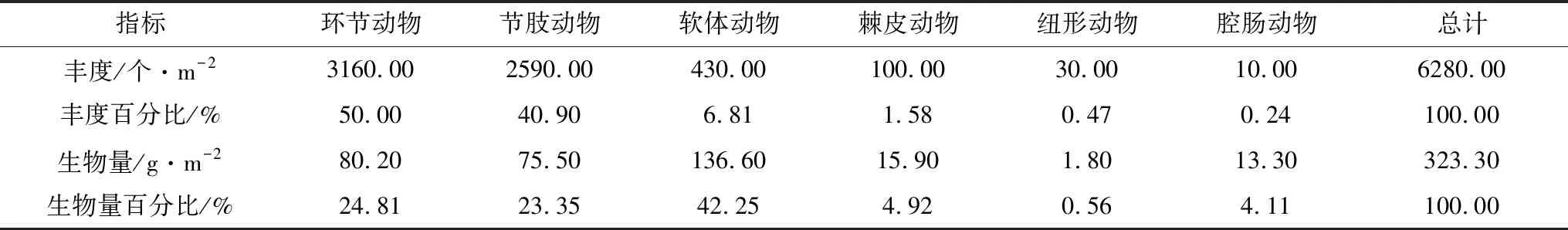
表4 曹妃甸近岸海域2016年5月大型底栖动物各类群的栖息丰度与生物量
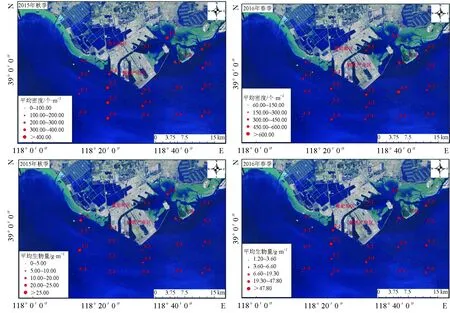
图2 曹妃甸近岸海域大型底栖动物丰度和生物量的空间变化
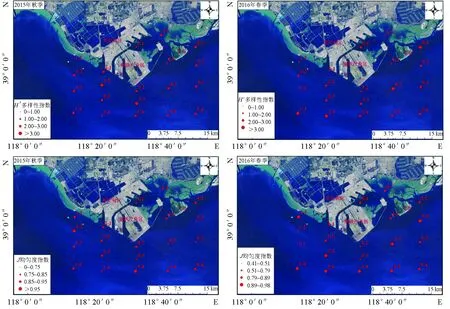
图3 曹妃甸近岸海域大型底栖动物多样性指数空间分布特征
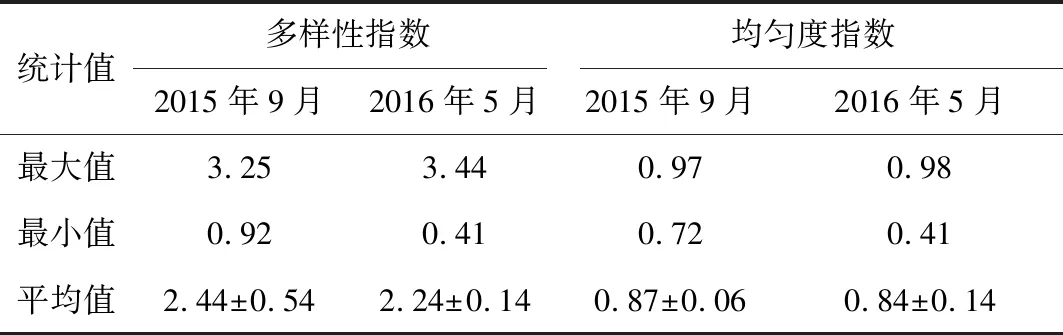
表5 曹妃甸近岸海域大型底栖动物多样性指数动态变化
2.2 大型底栖动物群落结构分析
大型底栖动物群落结构聚类分析和非度量多维尺度分析结果一致(图4)。双因素相似性分析表明,两次调查的不同聚类组间均存在显著差异(2015年,R=0.612,P<0.01;2016年,R=0.495,P<0.01)。
以20%的相似性,2015年秋季研究区域的大型底栖动物群落可以分为4组,其中1-3站位为单独一组,其他3组分别为:
组Ⅰ:由调查站位3-1、4-3、4-1、5-4、4-2、4-4、3-2、2-2、2-3、3-3、3-4、1-4、2-1共计13个站位组成,组内平均相似性29.14%,主要贡献种为囊叶齿吻沙蚕32.85%和日本刺沙蚕26.89%。
组Ⅱ:由调查站位5-2、5-3、1-2、5-1共计4个站位组成,组内平均相似性50.02%,主要贡献种为日本大螯蜚、马氏刺蛇尾(Stegophiurasladeni)、中华蜾赢蜚(Corophiumsinensis)、智利巢沙蚕(Diopatrachiliensis)、日本尾突水虱(Cymodocejaponica),贡献率依次为31.76%、24.46%、14.21%、10.16%、9.66%。
组Ⅲ:由调查站位1-1和2-4组成,组内平均相似性为23.15%,主要贡献种为日本大螯蜚,贡献率为100%。
以10%的相似性,2016年春季研究区域的大型底栖动物群落可以分为7组,其中2-2站位和3-1站位单独分组,其他5组分别为:
组Ⅰ:由调查站位5-1、5-2、5-4、2-3组成,组内平均相似性25.42%,主要贡献种依次为彩虹明樱蛤(Moerellairidescens)(49.62%)、深沟毛虫(Sigambrabassi)(10.47%)、滩拟猛钩虾(Harpiniopsisvadiculus)(9.81%)、蛇杂毛虫(Poecilochaetusserpens)(9.79%)、日本大螯蜚(8.20%)和不倒翁虫(Sternaspisscutata)(6.78%)。
组Ⅱ:由调查站位1-1和4-1组成,组内平均相似性21.83%,主要贡献种为智利巢沙蚕,贡献率为100%。
组Ⅲ:由调查站位3-2和3-3组成,组内平均相似性为38.91%,主要贡献种为海涟虫(Cumaceasp.)和海萤(Cypridinasp.),贡献率为52.09%、47.91%。
组Ⅳ:由调查站位2-1、1-2组成,组内平均相似性为46.12%,主要贡献种为日本大螯蜚和马氏刺蛇尾,贡献率分别为62.53%、37.47%。
组Ⅴ:由调查站位4-2、4-4、1-3、1-4、5-3、4-3、2-4、3-4共计8个站位组成,组内平均相似性24.50%,主要贡献种为囊叶齿吻沙蚕和小头虫(Capitellacapitata),贡献率分别为46.87%、56.93%。
2.3 底栖生境质量评价
2.3.1 丰度/生物量比较曲线法评价
2015年9月秋季和2016年5月春季对调查海域各站位的大型底栖动物生物量和丰度数据做丰度/生物量比较曲线并得到W统计值(图5)。2015年9月秋季,3-1(-0.13)、5-3(-0.04)和5-4(-0.007)站位W值小于0,其中3-1站位W值最小,且该站位的丰度/生物量比较曲线线中丰度远在生物量曲线之上,表明该站位底质环境受到剧烈扰动(图6)。2016年5月春季,1-3(-0.67)、4-2(-0.11)和4-1(-0.03)站位W值小于0,其中1-3站位W值最低,主要因为该站位仅出现两个物种,分别为囊叶齿吻沙蚕和纽虫(Cavernulariasp.),且囊叶齿吻沙蚕丰度高于纽虫丰度,但其生物量低于纽虫生物量,因此导致该站位丰度曲线在生物量曲线之上;另外,4-2站位的丰度曲线也在生物量曲线之上,说明1-3和4-2这两个站位底质环境均受到剧烈扰动(图6)。
分别汇总2015年秋季和2016年春季20个站位生物量和丰度数据得出的调查海域丰度/生物量比较曲线(图7)。由图7可知,2015年秋季,大型底栖动物丰度曲线位于生物量曲线之上,表明底栖群落受到严重污染扰动,而到2016年春季,底栖动物群落生物量优势显著上升,群落受扰动状况改善,达到未受污染扰动状态。
2.3.2 ABMI和M-ABMI指数评价
2015年9月秋季,曹妃甸近岸海域AMBI值为0.42~2.44(表6),最低值出现在4-3站位,该站位主要由对扰动敏感的物种组成(EGⅠ占比77.8%),最高值出现在4-1和4-2站位,第二级机会种(EGⅣ占比分别为62.8%和75.05%)为该站次的主要优势种。AMBI评价结果显示,1-3和4-2站位为底栖群落结构无扰动状态,其余站位为轻微扰动状态。M-AMBI值介于为0.39~0.89,最低值出现在1-1站位,该站位的种类数和多样性指数最低,最高值出现在3-3站位,该站位的种类数和多样性指数最高。M-AMBI生态环境状况评价结果显示,1-1和3-1站位,底栖生态环境已经开始退化,处于中等级别;2-2、2-3、3-3站位底栖群落处于正常状态,底栖生态环境未退化,处于优状态;其余站位底栖群落处于轻微失衡状态,底栖生态环境未退化,但处于良好状态。由空间分布上来看(图8),曹妃甸填海区东侧龙岛附近海域AMBI值较高,但其仍属于轻微扰动界定范围内;其余调查站位AMBI值相对较低。M-AMBI值呈斑块状镶嵌分布,这与曹妃甸海域独特的地理环境有关。
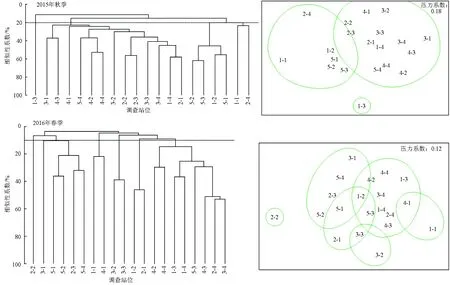
图4 曹妃甸近岸海域大型底栖动物站位聚类分析和多维尺度排序分析示意
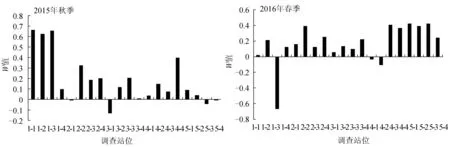
图5 曹妃甸近岸海域各站位W值
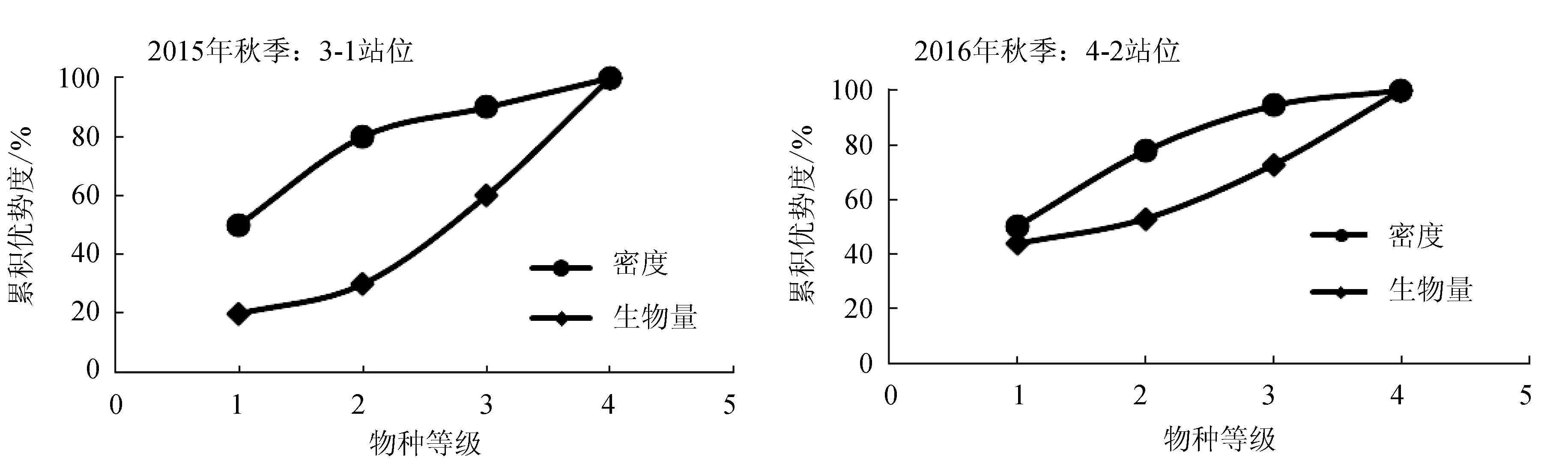
图6 2015年9月调查海域3-1站位和2016年5月4-2站位丰度/生物量比较曲线
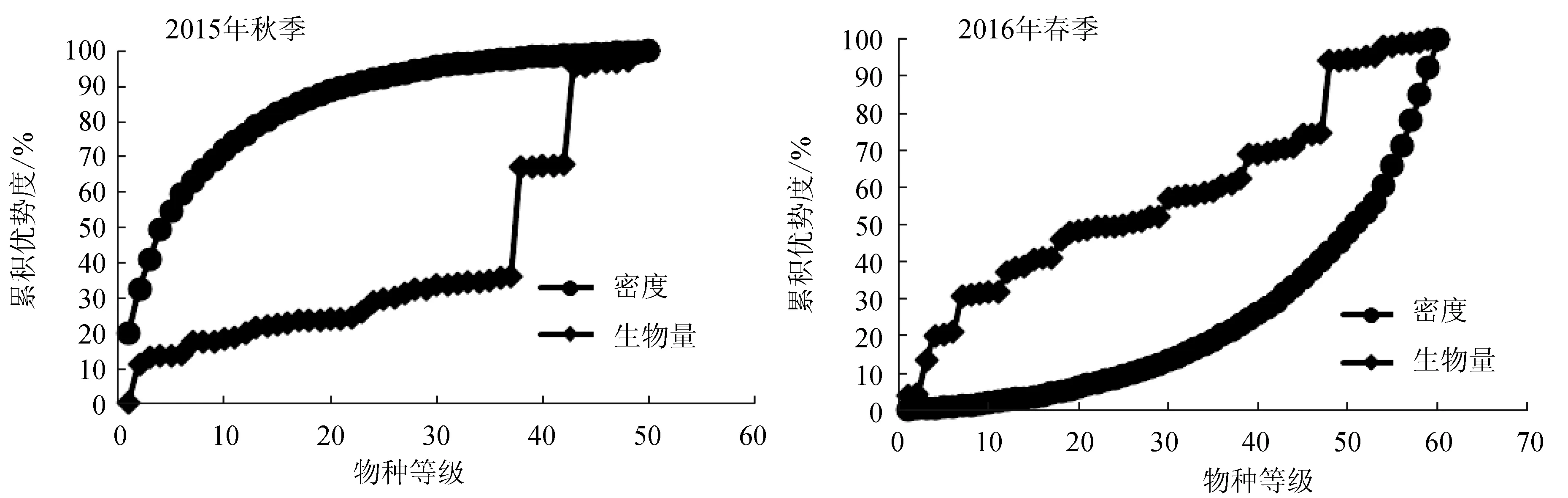
图7 曹妃甸近岸海域大型底栖动物群落2015年和2016年丰度/生物量比较曲线
2016年5月春季,曹妃甸近岸海域AMBI值为0.66~3.00(表6),最低值出现在3-3站位,该站位主要由为对扰动敏感的物种(EGⅠ占比55.80%),最高值出现在1-1和4-1站位,此两个站位主要由对扰动不敏感的物种(EGⅡ分别占比50.00%、41.70%)和第二级机会种(EGⅣ分别占比50.00%、41.70%)组成。AMBI评价结果显示,3-3和5-2站位为无扰动状态,其余站位为轻微扰动状态。M-AMBI值为0.37~0.84,最低值出现在1-3站位,该站位的种类数和多样性指数最低,最高值出现在4-3站位,该站位的种类数和多样性指数最高。M-AMBI生态环境状况评价结果显示,1-3站位底栖生境质量退化较严重,处于差的等级,其主要原因为此站位的物种数量仅为两种,而且生态等级为EGⅡ的囊叶齿吻沙蚕的丰度占该站位总丰度的91.70%,过低的物种数和EGⅡ较高的丰度比例是导致1-3站位M-AMBI指数低、生境质量等级差的原因;1-1、2-1、4-1、4-2站位底栖生态环境已经开始退化,处于中等级别;2-4、4-3、5-1站位底栖底栖生态环境未退化,处于优状态;其余站位底栖生态环境未退化,处于良好状态。由空间分布上来看(图8),曹妃甸填海区东侧龙岛附近海域AMBI值有所降低,扰动减轻,在曹妃甸填海区东侧港池附近AMBI值升高,扰动有所增强,但均属于轻微扰动范围内;M-AMBI值也呈斑块状镶嵌分布。
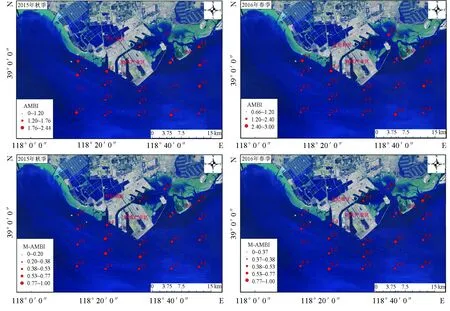
图8 曹妃甸近岸海域2015年和2016年航次调查各样点AMBI值和M-AMBI值空间分布
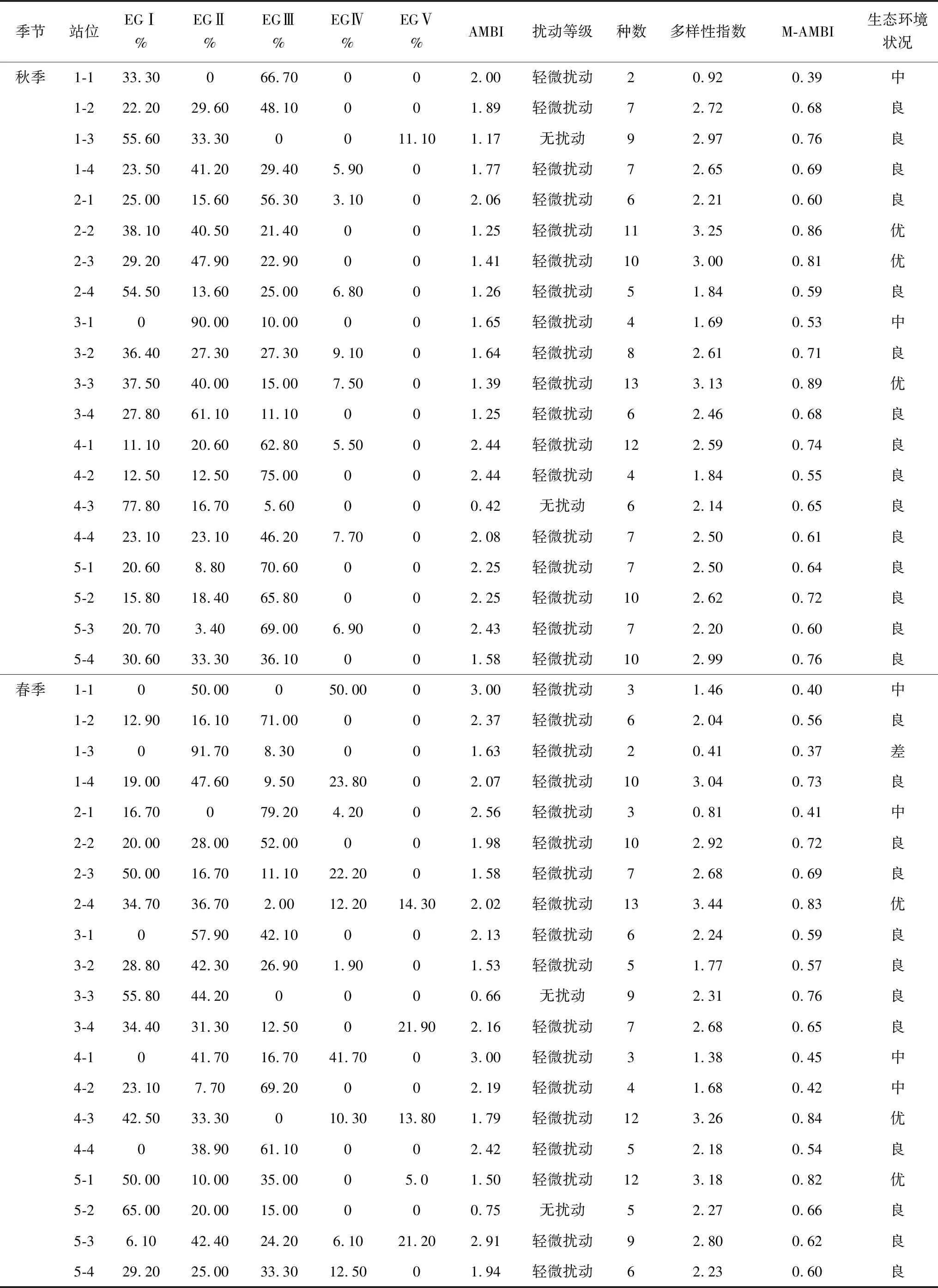
表6 曹妃甸近岸海域AMBI、M-AMBI指数值及生境质量状况
续表
另外,对比2015年9月和2016年5月的评价结果可知,AMBI指数和M-AMBI指数差异不显著(AMBI: t=-1.504,P=0.141>0.05;M-AMBI:t=1.423,P=0.163>0.05)。
3 讨 论
3.1 曹妃甸近岸海域大型底栖动物群落组成和优势种分析
根据本研究结果,2015年秋季曹妃甸近岸海域共发现大型底栖动物5门50种,到2016年春季,物种数有所上升,为62种,两次调查主要由个体较小的多毛类物种组成。李晓静等[11]对曹妃甸近岸海域2013年和2014年大型底栖动物的调查结果显示,其物种数分别为128种和143种。与之相比,本次调查的物种数明显下降。一方面可能是由于渤海近年来受到污染日趋严重,水交换能力减弱,渤海底栖生态系统衰退严重[22-23]。历史上,渤海湾的大型底栖动物种类非常多,如1983年调查的物种数为122种[24]。进入21世纪以来,渤海湾大型底栖动物发生较大的变化,物种数呈现下降趋势[25-26]。渤海湾作为我国重要的海洋经济区之一,各种生产活动产生的废水较多;加之渤海湾是一个封闭的海湾,水体交换能力弱,加剧了水质下降趋势,造成许多底栖动物无法生存,表现为种类降低[27]。与临近海湾相比,渤海湾的大型底栖动物种类仅高于辽东湾(46种)[28],低于胶洲湾(59种)[29],更低于莱洲湾(214种)[30]。这表明,渤海湾环境压力依然严峻。另一方面,随着曹妃甸工业区填海造地的完成,一些工业及码头工程陆续进入运营状态,海上运输及溢油的频发给该海域造成了巨大的生态压力,其水环境质量和沉积环境质量的变化都可能直接或潜在地影响大型底栖动物群落组成的空间分布格局[31],进而造成物种数明显下降。渤海由于其海底地形、海流和水团结构复杂的特点,导致其海洋生物栖息环境复杂多样[9]。然而,生物群落中优势种的优势度与群落稳定性密切相关,结构稳定的生物群落中优势种的优势度并不显著,脆弱的生物群落中优势种的优势度会比较高[32]。2015年和2016年调查结果显示,曹妃甸近岸海域主要以节肢动物和环节动物为主,其分别占比为52.50%和50.00%,日本大螯蜚和囊叶齿吻沙蚕分别为两次调查的主要优势种,其优势度分别为0.090和0.026,由此推测,该海域的大型底栖动物群落结构相对脆弱。另外,从调查种类多为节肢动物、环节动物等个体较小的种类,同时结合多样性指数和均匀度指数的分布来看,曹妃甸附近海域多样性指数和均匀度指数较低,说明曹妃甸近岸的底栖生物出现小型化和低质化现象,进一步证实了本次调查区域底栖生态群落结构相对比较脆弱。本研究结果与李晓静等[11]研究结果一致,曹妃甸海域底栖物种小型化依然明显[11]。
3.2 曹妃甸近岸海域大型底栖动物群落结构
根据大型底栖动物群落结构聚类分析和非度量多维尺度分析结果表明,调查海域大型底栖动物群落的相似性较低,其中,2015年可将不同调查站位的大型底栖动物群落在20%的相似性水平上划分为4个不同的聚类组,并且非度量多维尺度排序结果的压力系数高达0.18。2016年可将不同调查站位的大型底栖动物群落在10%的相似性水平上划分为7个不同的聚类组,并且非度量多维尺度排序结果的压力系数高达0.12。造成各调查站位大型底栖动物群落低相似性的原因可能与底栖动物分布海域的底质沉积环境有关[31]。渤海海域底质条件多变、海流复杂及温盐的变化和差异等不稳定[9],这些特点导致其群落结构空间异质性较高,相似性较低。在其他海域也出现类似现象,张蒙生等[34]对山东半岛南部近海的大型底栖动物的群落特征进行了分析,在23%相似性水平上,调查区域大型底栖动物群落呈明显地域性分布,且沉积物类型是影响调查区域群落分布的主要因素。另外,严重的陆源污染输入、高强度的渔业捕捞压力[35]等人为扰动也可能会改变底栖生境的多样性和底栖生物的群落结构,底质环境受到强烈的扰动,在一定程度上限制了大型底栖动物的生存与发展[36],进而导致底栖动物群落结构异质性增强。类似的情况在长江口海域和渤海湾均有发生[37-38]。
3.3 不同评价方法对底栖生境质量评价合理性分析
丰度/生物量比较曲线、AMBI指数及M-AMBI 指数法均能在一定程度上反映底栖动物受扰动状态,近年来,已被用于港湾、近岸海域以及河口区的底栖生态环境质量评价[1,9,33,39]。丰度/生物量比较曲线能较客观地反映出海洋环境污染对底栖动物群落结构的影响[39],是一种根据物种栖息丰度和生物量计算其对环境中各类参数响应的方法[40]。AMBI指数及M-AMBI指数法是欧盟水框架指令评价方法组成之一,它们被广泛应用于河口和海岸带水域的水环境质量检测和生态研究中[41-42]。这些生物指数的应用,提升了人们对海洋底栖无脊椎动物群落的生态功能认知,使得底栖生物群落水平上的环境指示作用更加迅捷和有效[39]。
由于海洋生态系统本身的复杂性以及人为环境压力的影响,不同的评价指数在同一区域通常给出不同的评价结果[8]。在本研究中,不同生物指数的评价结果往往存在一定的差异。丰度/生物量比较曲线研究结果显示,2015年秋季底栖群落受到严重污染扰动,而到2016年春季,底栖动物群落生物量优势显著上升,群落受扰动状况改善,达到未受污染扰动状态。这可能由于2015年底,整个曹妃甸工业区填海工程已经基本实施完毕,随着围填海面积不断扩大,曹妃甸海岸带自然属性被改造程度日益加深,由此带来海岸线增长,潮间带宽度减少、近岸冲淤环境改变、洪水、生物多样性损失等一系列严重后果,原始栖息地大面积丧失,进而导致2015年调查海域底栖群落受到严重污染扰动,随着填海工程的结束,底栖生物群落结构逐渐恢复,物种种类数量及生物量较2015年调查有所上升,群落扰动消失并呈现恢复状态。
AMBI评价结果显示,2015年和2016年90%站位的底栖环境属于轻微扰动状况,10%的站位属于无扰动状况。M-AMBI指数评价结果显示,2015年,90%的站位底栖生境质量达到优良状态,10%的调查站位达到中等状态。而到2016年,生境质量为优良状态的站位比列有所下降,为75%;处于中等环境质量的调查站位比例有所增多,达到20%,其中2015年秋季2-1、4-1和4-2站位生境质量为良好,而到2016年春季其生境质量下降,达到中等水平;另外1-3站位由2015年秋季良好状态之间下降到较差水平。
AMBI、M-AMBI指数评价结果与丰度/生物量比较曲线的评价结果不一致,分析原因:首先,丰度/生物量比较曲线法是一种作图方法,不同于有精确数值的指数方法,同时该方法仅将污染分为3种程度,即严重污染、中度污染和无污染,因而与生物指数的评价无法一致;其次,AMBI指数是严格基于底栖生物敏感度分类的指数[42],M-AMBI为综合了AMBI、生物多样性和物种丰度的复合指数,并通过因子分析和判别分析计算指数值[21],被认为更能够反映人为环境压力[43]。由此可知,AMBI指数评价结果仅是基于大型底栖动物自身对周边环境变化的敏感程度,没有考虑到周边的其他环境因素的变化,其评价结果有些狭窄。而M-AMBI评价结果综合多种因素,包括底栖动物自身的敏感程度、周边生态环境变化情况及人类对环境造成的压力情况等等,因此其结果更能真实反映该区域的底栖生境质量状况。AMBI和复合指数M-AMBI已经被应用于不同的水体类型(近岸海域、河口以及湖泊),大部分的应用结果都表明,M-AMBI指标比单因子指数AMBI更能够反映环境的实际情况[44],这与本研究的结论一致。
