三七育种研究综述
2019-07-22胡永媛高丽芳孙玉琴陈中坚
熊 高,王 勇,胡永媛,高丽芳,孙玉琴,陈中坚
(文山学院 文山三七研究院,云南 文山 663099)
三七 [Panax notoginseng(BurK.)F.H.Chen]为五加科人参属多年生草本植物,其主要活性成分皂苷对代谢、血管、中枢神经系统类疾病具有很好的疗效[1-3],在400余年的栽培历史中,已发展成为产业化程度高、市场规模大、研究较为深入的名贵中药材品种[4]。同时,在2016年云南省政府提出的实现三七产业千亿目标的大背景下[5],将为三七及其产业在种质资源保护、开发利用、改良创新等方面提供良好机遇。
品种作为一个产业得以持续健康发展的基础,而育种则是实现品种高产、优质、抗性优良最有效、最根本的途径。三七从驯化到人工种植至今,其品种选育工作经历了漫长的岁月,在此期间,通过许多三七育种工作者的辛勤付出,三七在品种选育研究上取得了重要的成就,为我国中药现代化科技产业发展和农民脱贫致富提供了产业支撑。
近年来,随着三七栽培面积的不断扩大,三七病害严重,加之连作障碍的影响,三七道地产区逐渐向非道地产区转移,这将导致三七药材品质下降,药效降低与道地性丧失。同时,由于三七抗病资源的研究利用进展缓慢,无很好的抗病品种,种植中人为的滥用农药,导致三七农残超标[6]。再者,随着公众日益增长的健康要求以及中药材品质升级无公害行动计划的提出[7],三七品种目前还远远满足不了产业的需求,特别是优质高产、抗病性强的新品种相对缺乏。因此,培育优良三七品种是三七科技产业发展的迫切需要,也可为三七生产优质药材奠定基础。本文综述了三七育种的发展历程,并对三七育种的未来研究进行了的展望,以供三七育种领域的研究者借鉴、参考和讨论。
1 三七传统育种技术的早期应用
传统育种主要基于现有的自然变异资源,通过系统选择、杂交、多世代筛选和纯化,最终获得具有目标性状的品种,整个选育过程周期长,成本高。由于三七生态适应性差、生长周期长、地理分布窄等特性,三七育种进展相比于其他粮食作物显得十分缓慢。自上个世纪90年代才开始对三七变异类型进行系统鉴定,获得了茎色、茎粗、块根、株高、叶宽等性状的变异类型[8]。之后,陈中坚等[9-10](2004年、2010年)进行相关和通径研究,最终认为宽叶三七、复叶柄平展型三七、紫根三七和长形根三七等4种变异类型可作为高产、优质三七新品种的育种目标。
以系统选育的传统育种技术是目前三七育种工作中最基本、最重要的育种途径。苗乡三七科技有限公司通过12年的系统选育,在2015年培育出“苗乡三七一号”(绿茎)和“滇七一号”(紫茎)两个三七品种并获得认定,这是文山州人工种植三七数百年来首获的品种,开创了三七育种新历史;同样,在多家单位的共同努力下,经过15年5个世代典型性状选择和纯化的系统选育,2019年文山学院与苗乡三七科技有限公司共同培育出以紫根性状为主的“文院紫七1号”和以复叶柄夹角小为性状的“苗乡三七2号”两个三七新品种。这些品种具有生活力强、优质、高产的特点。此外,杂交选育、倍性育种、辐射诱变等技术在三七传统育种研究中都有报道,比如,孙玉琴[11-12]等对三七开花、散粉规律研究,确定了三七杂交育种的最佳授粉时间为10∶00~14∶00,花粉常温下可保存8 h,低温储藏时也不宜超过20 d;王朝梁等[13]利用秋水仙素对三七种子进行染色体加倍,获得株形高大、叶片增大、叶色浓绿等明显特征的四倍体植株;三七60Co-γ辐射诱变研究显示种子的半致死剂量为54 Gy,为后续三七辐射育种工作奠定基础[14]。
2 三七现代育种技术的初步实践
随着分子遗传学和分子生物学的不断发展,现代育种技术在动植物育种领域得到大量应用,以随机扩增多态性(RAPD)、扩增片段长度多态性(AFLP)、简单重复序列(SSR)和单核苷酸多态性(SNP)等分子标记作为辅助进行的品种选育技术日趋成熟[15-16],这将有助于缩短育种年限,提高育种效率。
在三七分子育种研究进程中,早期的分子标记技术应用主要着重开展对三七的遗传多样性研究。2003年时就报道了以10bp寡核苷酸引物长度的RAPD分析方法检测三七栽培群体的变异情况,结果表明了不管是三七不同形态性状间,还是同一形态的不同个体间的多态性都很高,说明三七遗传背景复杂,还是一个杂合居群,具有丰富的遗传多样性,并指出需通过套袋自交、花粉培养等途径来进一步纯化三七遗传背景,形成可供育种选择具有稳定遗传变异类型的材料[17]。在2005-2008年间,主要以AFLP或荧光AFLP标记对三七的遗传多样性进行研究,特别是在三七栽培群体中不同植物学性状(株高、茎色、叶片、根重)的遗传多样性[18]、三七不同个体根部皂苷含量差异的遗传多样性[19]以及三七与屏边三七的遗传分化[20]等方面,这些结果显示了三七仍保持较高的生物多样性,不过经人工栽培后,其遗传分化相较于野生近缘种降低。其后,Liu等人[21]和张金渝等人[22]分别开发出了用于三七生物多样性研究的SSR和EST-SSR标记,这些标记在三七中均显示出高多态性,为评估三七遗传多样性及纯系提供了标记资源,其中发现通过集团选择方式可逐渐纯化选育群体,不过不同的选育材料之间仍存在较大的遗传分化[22]。随着二代测序技术的发展,可对三七在全基因组水平的遗传差异进行全面评价,通过简化基因组测序(RAD-Seq)技术对12个不同区域的36份三七进行分析,获得近六千多个SNP位点,群体结构显示三七的主要遗传变异(96.5%)发生在居群内,而居群间遗传分化水平低(3.5%)[23]。以上研究不仅开发出大量的分子标记,还提供了对三七遗传背景和变异情况的认识,为三七分子育种奠定理论基础。
通过前期的不断研究和实践,2016年,以DNA标记(SNP)辅助选育的首个三七抗病新品种“苗乡抗七1号”通过审定,获得品种认证,实现了在三七中应用分子标记辅助的现代育种技术育成三七抗病新品种的突破。该品种基于三七抗病资源和高通量测序技术,挖掘三七抗根腐病关联性SNP位点,根据该位点对抗病群体进行快速检测和选择,最终选育而成[24]。与常规栽培种相比,接种根腐病致病菌Fusarum oxysporum后“苗乡抗七1号”种子、种苗、块根发病指数分别下降52%、72.1%、62.4%,表现显著的抗性[25]。“苗乡抗七1号”的获得标志着三七及其人参属植物育种研究进入了以分子生物技术为手段的分子育种时代,这将对加快药用植物新品种选育和推广具有积极推动作用。
3 组学大数据挖掘三七育种新潜力
组学大数据不仅可以快速揭示杂交育种潜在的深层次机理,也为品种选育提供大量分子靶标和重要的遗传信息。2011至2016年间,三七组学分析主要基于转录组和蛋白组研究,这些结果在三七的三萜皂苷生物合成、生物碱合成、响应重金属砷胁迫方面展现了许多关键基因[26-30]。2017年,三七完成全基因组测序,获得3万多个编码基因,并首次系统性的解析了三七皂苷的合成途径,获得大量皂苷合成的候选基因[31-32]。目前,已有报道显示基因组、转录组、代谢组等多组学大数据用于三七研究分析[33-36],这些研究为今后三七高品质育种奠定基础,以基因组为支撑的三七遗传育种时代已经到来。
功能基因挖掘一直是生物学基础研究的焦点,也是基因组育种时代生物学家追寻的目标。在三七基因组、转录组、代谢组等组学大数据推动下,三七功能基因研究也取得了一些突破。在三七皂苷合成方面,孙颖等[37]利用RNAi技术在细胞水平下调三七环阿屯醇合酶基因CAS表达,阻断甾醇的合成,间接增加了三七皂苷含量;杨延等[38]在三七细胞中过量表达三七法呢基焦磷酸合酶基因FPS可提高皂苷含量;李建等[33]通过对一年生和三年生三七的转录组学分析,鉴定到皂苷生物合成关键基因达玛烯二醇合成酶DS和P450单加氧酶CYP716A47-like在三年生三七中显著高表达,并利用基因转化技术在烟草中验证了功能。在三七抗病研究方面,杨丹等[39]克隆表达了三七病程相关蛋白PR10-2,通过体外抑菌活性分析显示PR10-2蛋白可抑制三七根腐病病菌腐皮镰刀菌和坏损柱孢菌的活性;另外,在个体水平的研究也报道了三七几丁质酶基因PnCHI1和多聚半乳糖醛酸酶抑制蛋白基因PGIP,对三七根腐病病菌茄腐镰刀菌具有很好的抗性[40-41]。上述三七功能基因研究结果为三七遗传育种工作提供了新的靶点。
4 现有三七新品种
在三七育种工作者的持续努力下,三七新品种实现了从无到有的突破,三七的新品种,最早是经过云南省非主要农作物品种登记的“文七1号”,但“文七1号”是在当时特定的历史背景下产生,并没有经过育种程序的选育,是将当时文山种植的三七统称“文七1号”,为今后选育的品种作对照。因此,目前经过育种程序选育产生的品种共有5个(表1),都是经系统选育的方法选育而成,其中“苗乡抗七1号”在系统选育的基础上,采用了分子标记辅助技术。
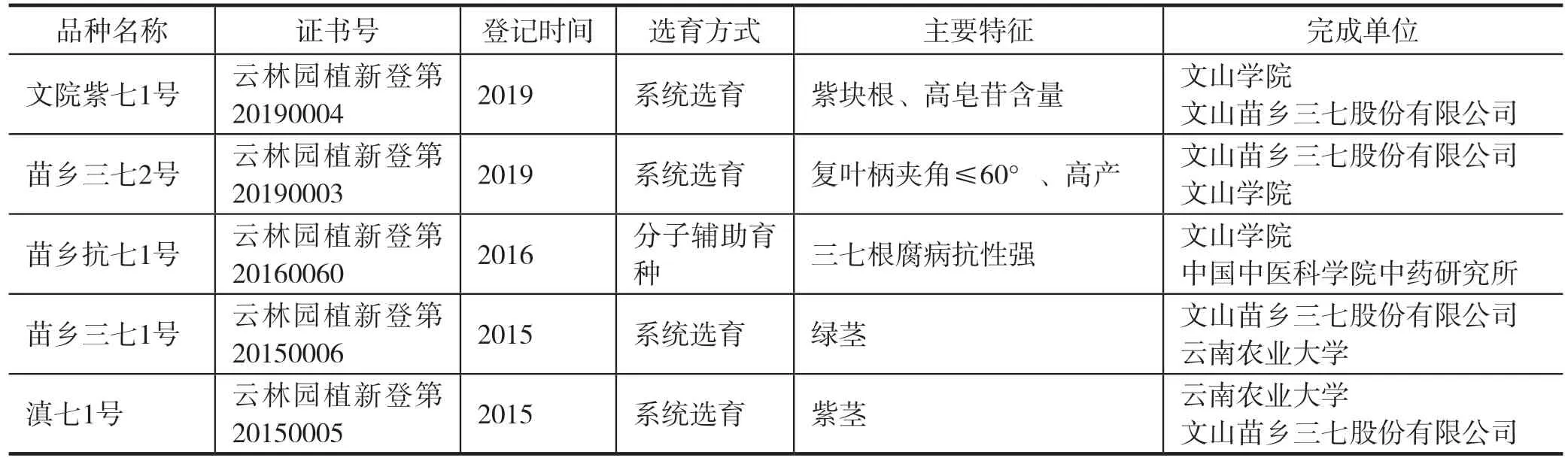
表1 三七品种(2015-2019年)
5 展望
三七育种研究经过长期努力取得了一定成绩,不过目前三七基础研究较为薄弱,系统选育方法仍是三七新品种选育的主要途径,这些因素很大程度上制约了三七育种研究的步伐。为实现三七育种的快速发展,作者就今后三七新品种选育研究工作提出以下4个方面的展望。
(1)完善三七种质资源保护与评价是三七育种继续长期坚持的重点工作。种质资源蕴藏着各种潜在的可利用基因,是育种过程的重要物质资源和原始材料。育种工作者很早就重视三七种质资源的收集和保存,目前文山三七研究院已建立了初具规模的三七种质资源圃,不过对三七种质资源的评价还很薄弱,在植物学性状、农艺性状、生物学特性等方面未进行系统性研究,这也是三七遗传育种工作进展缓慢的重要原因之一。因此,在今后的育种工作中,应相当长的时间着重对三七种质资源进行系统评价,培育纯系,只要对种质资源隐藏的信息有了全面了解,将有助于增加育种和选种的预见性,通过各种育种方法和技术,才能不断培育出高产、优质、多抗、适应性强的新品种。
(2)推进三七的倍性育种研究是快速实现三七新品种创制的有效途径。倍性育种是植物育种的重要方式,其可以通过减少或增加染色体的数量获得具有优良性状的变异个体。一般地,单倍体植物通过染色体加倍,即在一个世代就可获得纯合株系,大大缩短育种年限;而多倍体植物往往具有植株巨大、抗逆性强、适应性好、营养成分高的特点。基于三七生长周期长的生物学特性和增产块根为目标的生产特性,开展三七的倍性育种来创制新种质、新品种将是一种有效途径,具有重要的应用前景。目前,三七的倍性育种相关工作还比较基础,染色体倍性诱导率低,仍需进一步探索诱导方式。所以,应积极推进三七倍性育种技术的研究,建立成熟的倍性诱导体系,使这一有效的育种手段早日应用于三七育种领域中。
(3)构建成熟的三七杂交体系是实现三七品种资源利用率高的基本保证。杂交育种是将两个或多个品种的优良性状通过交配集合在一起,快速获得拥有多个优良性状的单一品种,从而提高品种利用。三七的杂交育种研究已经在三七的花器结构、开花习性、散粉习性和花粉活力等领域取得了一定的进展,今后还将继续探索三七杂交技术、交配组合等一系列基础研究工作,建立成熟的三七杂交体系,实现多元杂交和多基因聚合育种,为三七杂交育种应用及实现品种有效利用提供保障。
(4)强化三七基础科学研究或可为助力三七分子设计育种应用奠定基础。三七的基因组研究得到了破译,转录组、蛋白组、代谢组等多组学也在三七中进行了大量应用,三七正在快步走向“发现基因、研究基因、利用基因”的研究模式。尽管目前三七的基础理论研究与创新方面获得了不错的进展,但是与其他农作物,比如水稻、玉米、小麦的科学研究相比还有一定距离,尚不能对三七育种研究提供有效支撑。那么,在如今生命科学高速发展的时代,为能实现三七在遗传育种研究中“一个基因一个品种”的模式,就亟需进一步强化三七基础科学研究,特别是针对三七在主要经济性状、抗性、适应性等方面的机理展开系统性研究,以获得更多效果显著、作用机制清晰的靶标分子,为三七分子设计育种应用奠定坚实基础。
