稻田体系中铁的生物地球化学过程及铁同位素分馏机制研究进展
2019-07-19李芳柏李勇珠
李芳柏*,李勇珠 , **
1. 中国科学院广州地球化学研究所,广东 广州 510640;2. 广东省生态环境技术研究所,广东 广州 510650;3. 中国科学院大学,北京 100039
铁是植物维持正常生命活动的必需微量元素之一,参与众多生物代谢过程,如生物体内光合作用、呼吸作用、氮素同化和固定及激素合成等(Briat et al.,1997)。植物自身不能合成铁元素,只能从外界环境中获得铁来满足自身生命活动的需要;但是铁的溶解性很低,植物可以利用的生物有效铁有限(Guerinot et al.,1994)。据统计,全世界有20亿人口贫血,膳食缺铁是人类贫血的首要成因,而水稻是全球三分之一人口的主食来源,因此了解水稻吸收铁的机制对提高其对环境中铁的吸收、转运和富集将有重要作用。
铁同位素已被广泛地用来追踪生物地球化学过程中的铁循环(Beard et al.,2003),比如异化铁还原(Crosby et al.,2005)、亚铁的生物和非生物氧化以及吸附、沉淀(Croal et al.,2004;Schuth et al.,2015)等过程。稻田体系中,水稻会通过改变铁吸收机制来应对不断变化的生境,而铁吸收机制的改变会形成不同的铁同位素组成特征。当土壤水位降低时,土壤中的铁从还原态变成氧化态,铁的溶解性和生物可利用性降低,水稻主要利用机理II途径,即螯合途径来吸收铁;反之,土壤水位升高时,铁多以还原态存在,生物可利用性提高,此时水稻可以通过机理I途径直接吸收亚铁。进入到水稻植株内以后,铁在各器官组织之间的运输涉及到铁的氧化还原、螯合和转运等过程,这些过程均会导致不同程度的铁同位素分馏,这些特定的铁同位素组成信息不容易受环境改变而保存在水稻植株内。因此阐明稻田体系铁同位素的分馏特征,对于研究水稻吸收铁的机制,以及铁在土壤-水稻体系中的循环转运有重要意义。
本文从3个方面进行文献综述:(1)水稻土中铁的迁移转换及铁同位素分馏过程;(2)铁在水稻内吸收和转运的生理机制;(3)水稻内铁同位素的分馏特征及机制。以期从这3个方面综合理解稻田体系中铁的生物地球化学过程。
1 水稻土中铁的同位素分馏特征及影响因素
铁是地球上丰度第四的元素,其地球化学行为作为土壤元素循环的重要组成部分而具有重大意义。铁稳定同位素方法在示踪铁元素地球化学行为上取得了很大的进展(Johnson et al.,2008;Wiederhold,2015)。在水稻土的风化发育过程中,轻铁会优先淋溶,且铁形态和同位素组成随着土壤发育的进行而发生变化。近期对于年代序列水稻土的研究结果显示不同发育年龄土壤之间的铁同位素组成范围为0.1‰-0.18‰,铁同位素分馏值大小与水稻土发育年龄成正相关关系(Huang et al.,2018)。同时,随着发育年龄的增加,土壤中弱结晶态和硅酸盐结合态铁的比例下降,而氧化态铁比例上升,这可能是长期人为的落干与淹水调控的结果。水稻对不同形态铁的利用效率不同,交换态和弱结晶态相对于氧化态和硅酸盐结合态更容易被水稻吸收利用,铁同位素的测定结果显示水稻吸收的交换态、弱结晶态富集 0.3‰轻铁,而基本不被水稻吸收的硅酸盐结合态富集0.4‰的重铁(Guelke et al.,2010),同时水稻植株相对土壤富集轻铁(Garnier et al.,2017)。这些研究表明稻田体系发育过程是一个轻铁缺损的过程,铁的循环涉及氧化还原、吸附、溶解、沉淀及生物作用等多个过程;同时这些过程因受 pH、Eh、微生物过程等的影响而呈现出复杂多样性。
孔隙水的铁是水稻土中最容易被吸收利用的铁源,铁主要以二价和三价铁存在,其中二价铁占主导地位。二价铁主要由铁的异化铁还原(DIR)过程产生(Crosby et al.,2007;Garnier et al.,2017),同时还受到硫酸盐还原过程的影响(Severmann et al.,2006)。水稻土处于长期淹水时,异化铁还原过程活跃,亚铁富集轻铁,铁同位素分馏值达到3‰,而且随着DIR过程的进行,不断有轻铁释放到孔隙水中,因此DIR过程被认为是孔隙水中轻铁同位素的主要来源。但是随着淹水时间的增加,以及土壤深度的增加,土壤Eh会大幅度下降。当Eh下降到-200 mV左右,土壤中 SO42-会被还原成H2S,H2S电离成HS-,之后与溶液中的Fe2+生成黄铁矿(FeS。Butler et al.,2005)。黄铁矿沉淀初始,轻铁优先被沉淀在 FeS中,且动力学分馏系数ε=0.85±0.30‰,随着沉淀过程的进行,动力学分馏被平衡分馏结果掩盖而使黄铁矿富集重铁,分馏值为0.32‰,这一数值远远小于DIR过程导致的分馏值(3‰。Romain et al.,2011)。异化铁还原和硫酸盐还原过程共同影响孔隙水中铁的同位素组成,硫酸盐还原过程使孔隙水富集轻铁的程度远小于异化铁还原过程。
除了铁还原过程外,水稻土中铁的氧化沉淀过程被认为是影响土壤铁同位素组成的主要因素之一,且受到pH值和氧气浓度的影响。当pH值高,亚铁氧化主要是非生物过程,当pH低于3时,亚铁氧化主要受非生物过程调控,比如光合细菌氧化,亚硝酸依赖亚铁氧化以及嗜酸性亚铁氧化过程(Balci et al.,2006)。Welch et al.(2003)的研究表明溶液中二价铁和三价铁之间存在平衡分馏,建立了铁同位素分馏值与温度间的关系:ΔFe(III)-Fe(II)=0.334×106/T2-0.88,且实验测定 22 ℃下的分馏值为2.76‰。Balci et al.(2006)在低pH下测定生物诱导亚铁氧化生成三价铁的平衡分馏值 ΔFe(III)-Fe(II)=2.9‰,且三价铁与随后生成的沉淀物之间的平衡分馏范围为0.58‰-0.98‰。氧化沉淀过程中,氧化占主导作用,产物总体富集重同位素。溶液中的Fe(II)与针铁矿之间的总体分馏值为-1.05‰,且溶液 Fe(II)与针铁矿上表层 Fe(III)的最小分馏值为-2.1‰。这些结果都表明铁的氧化以及成矿过程均发生铁同位素分馏,且氧化成矿的产物总体富集重铁同位素。
含铁矿物的溶解过程也会发生同位素分馏。溶解过程主要有异化铁还原溶解、质子溶解和吸附溶解等过程。HCl溶解针铁矿被认为是不会导致同位素分馏的溶解方式,而草酸诱导的吸附溶解和还原溶解过程都会导致轻铁优先溶解,平衡分馏值为0.5‰(Wiederhold et al.,2006)。而草酸吸附溶解优先释放重铁,原因是形成的oxalate-Fe的键强要高于 Fe-O键。跟氧化还原作用相比,吸附作用是造成铁同位素分馏的次要原因。Dideriksen et al.(2008)测定溶液中有机和无机铁之间的平衡分馏值为 0.6‰,其中有机铁为脱铁敏物质与铁的络合物。除实验测定外,理论计算结果也表明有机配合物可以络合重铁同位素,例如 Fe(III)-phytosiderophore与Fe(III)之间的理论分馏值为 1‰左右(Moynier et al.,2013),但是在土壤发生过程中,有机物仍然对铁同位素分馏起重要作用。例如Wiederhold et al.(2007a,2007b)分别测定了氧化还原土和灰壤在不同剖面和不同形态铁之间的同位素分馏值,其中灰壤提前经过一年的曝气处理,限制其氧化还原过程。研究结果表明两种土壤均表现出轻铁先淋溶,但是氧化还原土中土壤剖面间的分馏值只有0.3‰,而在灰壤中,分馏值达到1‰。两种土壤之间存在较大铁同位素组成差异可能是因为灰壤形成于充分水分淋溶的环境下,且存在强酸性腐殖质等有机络合物,有机物的络合作用可以帮助铁同位素发生迁移而增大同位素分馏效应。
土壤中铁的分馏还受到诸多因素影响,其中土壤铁含量是一个必须被重视的因素。国内外文献数据汇总结果呈现出铁同位素分馏值与铁浓度成负相关关系的特征(Huang et al.,2018),Wiederhold(2015)提出储库对同位素分馏的影响很大,大的储库可以削弱同位素分馏效应,反之,小的储库会增大同位素分馏效应。正是由于土壤铁同位素的产生是多因素的,将铁同位素方法用于稻田铁元素循环时应结合土壤发育背景、土壤化学生物性质等因素。
2 水稻吸收转运铁的生理机制
植物吸收铁的机理有两种,分别为机理I和机理II,其中双子叶植物和非禾本科单子叶植物运用机理I吸收铁,禾本科单子叶植物运用机理II吸收铁(Marschner et al.,1986;Römheld,2010)。机理I基于铁还原吸收铁途径,同时伴随着3种物质的分泌:(1)三价铁螯合物还原酶,它由铁缺乏诱导生成,可以还原三价铁至亚铁螯合物;(2)H+,它能降低根际pH值,提高铁化合物的溶解度,促进三价铁的还原;(3)酚类物质,具有还原和螯合作用,具有多种潜在功能。机理I吸收过程依赖于可溶性铁的提供,受pH影响大。机理II过程基于铁的螯合吸收,具有两大特征:(1)分泌植物铁载体类化合物(phytosiderophore,PS),这些化合物在缺铁环境下诱导生成,能螯合溶解度很小的铁化合物,如氢氧化铁;(2)拥有一套高亲和性的铁载体运输系统,能够特异性运输铁螯合化合物进入植物根系。机理II过程主要由植物铁载体(PS)释放速率来调控,受pH影响很小。
2.1 水稻根吸收铁的机制
水稻作为禾本科单子叶植物,除了采用机理II吸收铁,也能通过机理I吸收铁(Ishimaru et al.,2006),研究发现玉米中也存在机理I吸收通道(Li et al.,2018)。Charlson et al.(2006)分析发现机理I和机理II植物均含有机理I铁吸收基因,但是机理I植物并没有机理II铁吸收基因,且裸子植物和单子叶中的祖先品种-拟南芥,也不含有机理 II基因,同时研究也表明机理II途径比机理I途径拥有更高的铁吸收效率,受pH和土壤含水量变化的影响小(Chaney,1984;Marschner et al.,1986),因而推测机理 II是从机理 I进化得到的铁吸收途径。水稻可能通过调节铁吸收机理来适应生长环境的变化,特别是当铁的供给发生较大变化时。
当水稻处于淹水环境,根际铁充足且可能因过量造成毒害作用。溶液中的亚铁直接通过二价金属转运蛋白进入根表皮细胞。水稻中已知的二价金属转运蛋白有 OsIRT1/2(Lee et al.,2009a)蛋白和OsNRAMP1/5(Ishimaru et al.,2012;Takahashi et al.,2011)蛋白,其中OsIRT蛋白对亚铁的亲和力高于OsNRAMP蛋白。根际溶液中多余的铁会形成铁的氢氧化物吸附和沉淀在根表形成铁膜,铁膜既可以作为缓冲带减少亚铁毒害,也可以作为备用铁库在铁供给不足时被水稻重新利用(Mi et al.,2013;Sebastian et al.,2016)。当铁缺乏时,水稻会通过OsPEZ1和OsPEZ2蛋白分泌酚类物质(原儿茶酸和咖啡酸)到根外,这些酚类物质可以增大根际土以及根质外体中铁的溶解性。
处于干旱条件时,铁主要以氢氧化铁存在时,水稻的三价铁螯合物还原酶基因(OsFRO)在根部的表达水平很低(Ishimaru et al.,2007),机理I途径的效率太低,因此水稻根细胞通过机理II的螯合途径获得铁。麦根酸类物质(Mugineic Acids,MAs)是一类植物铁载体(PS),包括麦根酸(MA)、2′-脱氧麦根酸(DMA)及3-羟基麦根酸(epiHMA)等多种物质。其中DMA是第一个生成的MAs,水稻根细胞内形成的DMA被转运输出蛋白运输到细胞外螯合 Fe(Ⅲ),OsTOM1(Nozoye et al.,2011)和OsTOM2(Nozoye et al.,2015)是两个已被鉴定的铁载体输出蛋白。根细胞外形成的 Fe(Ⅲ)-PS螯合物通过 YSL(yellow-stripe1-like)家族蛋白进入植物根表皮细胞,水稻根中已鉴定出的运输Fe(Ⅲ)-PS螯合物的YSL家族蛋白有OsYSL15(Lee et al.,2009b)和 OsYSL16(Kakei et al.,2012)蛋白。水稻分泌PS的活性在一天中波动很大,Ueno et al.(2009)对多年生草本植物的研究发现,PS的分泌主要受温度控制,且水稻分泌 PS的能力弱于大麦,燕麦等禾本科植物,这可能是水稻在石灰土壤中易缺铁的原因之一(石灰土壤的高pH值会降低铁活性)。
2.2 铁在水稻内的短距离运输
铁从根表皮细胞进入木质部的过程为径向运输过程,也叫短距离运输过程,主要有两条途径:一条是共质体途径,铁借助胞间连丝在细胞和细胞之间传输,是主动运输过程,铁的移动速度慢;另一条是质外体途径,铁在由细胞壁和细胞间隙组成的区域运输,是被动运输过程,铁的移动速度快。经质外体运输的铁到达内皮层后,被内皮层细胞壁上具有疏水性的凯氏带阻隔,而后加入共质体途径。由于根尖处的凯氏带未发育成熟,侧根处的凯氏带被破坏,当根际溶液中铁浓度很高时,可以经由根尖和侧根等质外体途径进入中柱。在共质体中,细胞液呈碱性,pH值为7-8,Fe(III)-DMA螯合物在输运前可以被维生素C等物质还原为Fe(II)(Weber et al.,2008),Fe(II)随后和尼克酰胺(NA)形成稳定的Fe(II)-NA螯合物。NA是形成DMA的前体物质,与二价铁有强结合能力(logK=12.1)(Beneš et al.,1983)。三价铁螯合物还原酶(OsFRO)、核黄素、NADH等物质也可以还原Fe(III)-DMA。共质体中的铁部分停留在细胞质和储存在液泡中,部分进入中柱的木质部,重新进入到质外体。
2.3 铁在水稻内的长距离运输
铁参与植物内运输是从进入木质部开始的,木质部是质外体的一部分。铁从内皮层细胞进入木质部所需要的蛋白还未研究清楚,但是已有研究发现OsIRT1和 OsYSL18均在内皮层细胞中有表达(Aoyama et al.,2009;Lee et al.,2009a),不过还无法确定OsIRT1和OsYSL18是将铁载入细胞还是载出细胞,铁以何种形式进入木质部也未有定论。木质部的pH为5-6,铁主要以Fe3+-柠檬酸复合物形式存在,少量以铁-PS复合物和离子态形式存在(Ariga et al.,2014)。柠檬酸由多药物和毒素排出(multidrug and toxin efflux,MATE)家族的 OsFRD1成员负责运输,OsFRD1主要在根的中柱鞘细胞中表达,异源表达结果显示其可以转运柠檬酸,并将铁-柠檬酸复合体运输至植物木质部(Yokosho et al.,2009)。木质部细胞为栓质化的死细胞,营养液运输至叶片,果实等库组织的动力来自于根压和蒸腾压力,运输方向为从下至上。营养液在水稻地上部器官中的分配取决于蒸腾速率和时间,各器官蒸腾速率强弱依次为叶片>果荚≥种子,水稻可能通过降低蒸腾速率的方法来抵抗高浓度铁的毒害作用(Shrestha et al.,2015)。在叶中,铁经木质部运输的终点是叶肉细胞。YSL家族蛋白担任铁从木质部卸载铁的载体,由于YSL蛋白不能运输铁-柠檬酸螯合物,在木质部卸载过程中,铁可能会发生形态的转化。其中NA既可以螯合Fe3+,也可以螯合Fe2+,可以作为穿梭体装载和卸载铁,对木质部和韧皮部之间的铁分布很关键:铁从木质部卸载之前与NA结合,经YSL蛋白转运至叶肉细胞(Curie et al.,2009)。YSL蛋白中的OsYSL2负责运输Fe-NA,但是不能运输Fe-PS,其表达部位主要有水稻根部、叶片韧皮部、花的维管束及发育的种子(Koike et al.,2004);相反,OsYSL15/16/18负责运输Fe(III)-PS(Aoyama et al.,2009;Inoue et al.,2009;Kakei et al.,2012),但不转运 Fe-NA。ZIP(Zn-Fe-regulated transporter)家族的OsIRT1、OsIRT2转运蛋白可以运输离子态铁进入叶肉细胞。
进入叶肉细胞后,铁大部分与铁运输蛋白结合(iron transport protein,ITP)形成Fe-ITP复合物并储藏在叶绿体中(Curie et al.,2009)。铁在水稻中属于移动性中等的元素,可以通过韧皮部重新运输至水稻中蒸腾作用较弱的器官和组织,例如新叶、籽粒等,这些组织由于缺少蒸腾压力而不能从木质部得到足量的铁。与木质部长距离运输不同,韧皮部的长距离运输发生在活细胞筛管中,向上运输至籽粒,向下运输至根部。韧皮部汁液的 pH值为7-8,存在大量蔗糖、小分子蛋白质和代谢物,除此之外,三价铁的浓度是二价铁的 10倍以上。文献统计表明机理I植物韧皮部三价铁大部分和谷氨酸、NA结合,二价铁全部和NA结合(Harris et al.,2012)。机理I植物韧皮部中亚铁氧化到三价铁的电势计算结果为247 mV,而Fe(Ⅱ)-NA的氧化电势为-181 mV;推测在韧皮部中,亚铁极易被氧化,NA结合亚铁可以防止其被氧化和水解(Harris et al.,2012)。但是对于水稻韧皮部铁形态的研究则显示,三价铁几乎全部和DMA结合,而NA主要结合 Cu2+和 Zn2+等二价离子(Nishiyama et al.,2012)。造成这种差异的原因可能是机理I植物缺少合成DMA需要的关键酶。DMA的合成需要尼克酰胺转移酶NAAT,GUS染色结果显示OsNAAT1在根的伴胞和中柱鞘细胞,以及茎叶韧皮部的伴胞中表达(Inoue et al.,2008)。表明DMA不仅在根部表达,协助铁的根吸收过程;也在韧皮部表达,促进水稻内的铁平衡。
2.4 铁在籽粒中的积累过程
水稻进入生殖生长期后,光合作用产物对根的供给减少,根因活性减弱而无法吸收足够的铁,水稻植株内的铁会经历空间重新分布的过程。其中新叶的铁主要由老叶提供,籽粒中铁营养主要来自上部叶片和旗叶,且已由实验证明主要通过韧皮部传输,少量铁经由木质部传输(Yoneyama et al.,2010;Yoneyama et al.,2015)。小麦中重新活化进入籽粒的铁占植株总铁的77%,在水稻中为4%(Morrissey et al.,2010)。铁进入并保存在籽粒中需要多个转运和运输蛋白的协调,其中,NA-YSL运输系统发挥了重要作用:NA结合铁离子,经细胞质膜上的YSL通道蛋白进入籽粒细胞。现已证实超量表达NA合成酶基因OsNAS2可以增加水稻籽粒的铁含量(Lee et al.,2012),转入大麦 NA合成酶基因HvNAS1后,水稻籽粒中铁和锌含量分别增加了2倍和3倍(Masuda et al.,2009)。这些都表明尼克酰胺NA可以帮助运输铁元素进入籽粒。Fe-NA进入籽粒需要通过YSL蛋白家族,OsYSL2蛋白对发育中的水稻籽粒表现出极大的促进作用(Koike et al.,2004)。YSL家族的另一个成员 OsYSL9则对籽粒中胚乳和胚芽间铁的分配起决定作用,敲除OsYSL9基因的水稻籽粒中,胚芽中的铁减少,而胚乳中的铁增加(Senoura et al.,2017)。ZIP家族的 IRT1蛋白对籽粒铁积累发挥重要作用,表示铁也以离子态进入籽粒中。最近的研究显示,OsFRDL基因也与籽粒铁的运输有关。OsFRDL1在水稻的第一个节、穗以及旗叶中表达水平高。OsFRDL1主要位于节的伴胞桥中,可以分泌柠檬酸将质外体中的铁运输至花粉、种子等水稻生殖器官中(Yokosho et al.,2016)。
铁从土壤到水稻,涉及多个生物化学过程,经历了多种形态变化,图1是根据文献总结的铁在稻田体系的转运过程示意图,表1是可能涉及的生物化学过程。
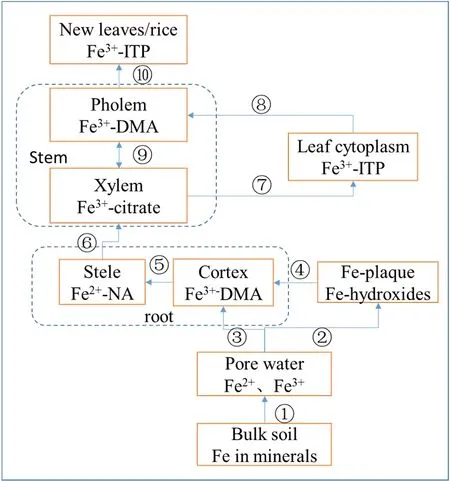
图1 稻田系统中铁转运过程Fig. 1 Iron transport process in paddy field system
3 水稻中铁同位素分馏特征及机制
目前仅在少数几种植物中开展了铁同位素组成的研究,结果显示植物与生长基质之间、植物内部均存在铁同位素分馏,且不同植物呈现出特有的铁同位素组成特征。植物间特有的铁同位素组成可能与铁在植物内的运输过程紧密相关,铁同位素的精确测定在揭示植物铁运输机制方面具备巨大潜力。

表1 稻田体系中铁的生物化学过程Table 1 Biochemical processes of iron in paddy field systems
3.1 高等植物铁同位素分馏特征及机制
Guelke et al.(2007)最早测定了高等植物体内的铁同位素分馏特征,他们发现机理I和机理II植物在铁同位素组成和变化趋势上存在很大不同:机理I植物相对于土壤富集轻铁,分馏值达到-1.6‰,且 δ56Fe值随着根茎叶种子等器官由下至上逐渐减小;而机理II植物整体富集重铁,分馏值为0.2‰,且各个器官间的组成几乎没有差异。猜测机理I植物富集轻铁是因为在吸收铁之前存在三价铁还原过程,且植物体内存在的多个氧化还原过程扩大了分馏值;而机理 II植物螯合吸收铁的过程中,PS的螯合作用会富集重铁同位素;各植物器官间铁同位素组成无差异是因为植物内不存在铁的氧化还原过程,或者其铁氧化还原过程是定量的。Guelke-Stelling et al.(2011)随后采用沙培的方式,以EDTA-Fe(III)为铁源分别种植典型机理I植物大豆和机理II植物燕麦,分4个生长期收割作物。铁同位素的测定结果与之前的类似:大豆和燕麦均富集轻铁,δ56Fe值从下至上依次减小,在种子中达到最小,为-2.5‰。而燕麦的δ56Fe为-0.5‰,各个器官间的铁同位素组成差异很小。机理I植物的铁同位素分馏趋势相近,表明其同位素分馏与生长环境没有直接关系,而燕麦的分馏值相对其他机理II植物增大可能是根吸收铁之前存在还原过程。
Kiczka et al.(2010)测定了阿尔卑斯山3种植物的铁同位素组成,其结果与Guelke et al.(2007)的略有差异:机理I和机理II植物均富集轻铁,机理II的富集程度小于机理I植物。机理I植物总体铁同位素分馏值Δ56Feplant-granite=-1.3‰,而机理 II植物仅仅富集 0.2‰的轻铁,这个结果则与 Guelke(2007)的结果类似:机理I富集因子为-1.6‰,机理II为-0.2‰。不同的是本研究中两种植物的内部均存在很大分馏。植物内部同位素组成最轻的为根中柱,Fe=-0.39‰,最重为皮层,Δ56Fe中柱-皮层=-4.5‰,可能是因为皮层质外体的铁进入中柱之前被还原,同时伴随着瑞利分馏过程(图1中的过程⑤)。小糠草(Agrostis)和杜鹃花(Rhododendron)两种植物的叶子在不同生长阶段,分馏分别增加了0.5‰和0.8‰。机理II植物小糠草不同生长时期的花的同位素值变化最大,为 1.6‰。植物内铁的重新运移过程是导致新老叶以及不同时期花之间铁同位素分馏的原因:运移过程涉及配体转化过程和氧化还原过程。根据测定值计算出的选择性吸收导致的分馏值Δ56Feafteruptake-cortex=-1-(-1.7‰),这与铁的异化还原过程导致的分馏值[-1-(-1.5‰)](Johnson et al.,2008)很接近,猜测根部选择性吸收主要涉及铁的还原吸收过程。
这3项关于高等植物铁同位素值的研究表明,机理I和机理II植物相对于土壤和培养液均富集轻铁。但是这两类植物在铁同位素组成上各自有其不同特点:机理I植物总的轻铁富集程度大于机理II植物,且机理I植柱内部在同位素组成上存在较大变异,而机理II植株内部在同位素组成上相对均一(图 2)。目前认为造成两类植物铁同位素组成呈现较大差异的原因是机理 I植物通过还原途径吸收铁,而机理II植物通过螯合过程吸收铁,且铁形态在植物内部不会发生变化。这些目前都只是猜想,未来的研究应该综合植物的生理过程来解释铁同位素的分馏机制。例如测定跟铁还原过程和铁螯合过程相关的基因的表达量来确定铁吸收过程是否发生了铁还原过程,如 OsFRO(铁还原基因)和OsYSL(铁螯合相关基因)基因的表达量。植物间分馏特征的差异除了跟植物类型有关之外,可能还跟铁营养元素的生物可利用性有关。例如机理II途径在螯合铁的过程中富集的重铁是相对于环境中可利用铁的铁同位素组成而言的,当环境的铁同位素组成偏轻,则机理II植物仍有可能富集轻铁同位素。因此解释植物铁同位素分馏机制时应该充分考虑环境中铁元素的铁同位素值和可利用性。
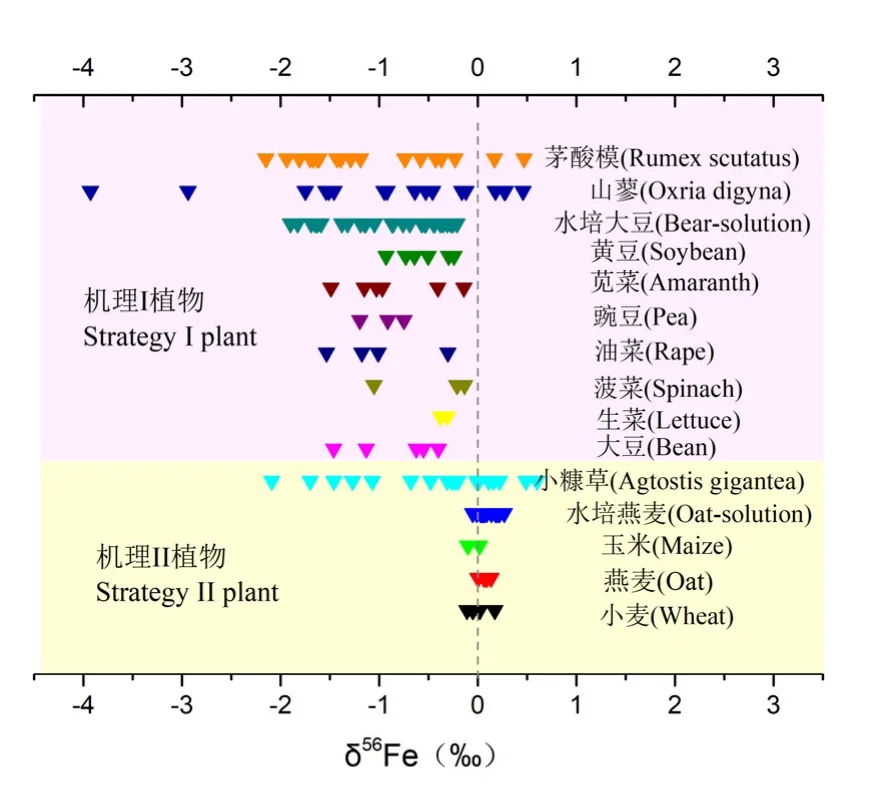
图2 高等植物铁同位素组成Fig. 2 Iron isotope composition of higher plants
3.2 水稻中铁同位素分馏特征及机制
水稻由于兼具机理I和机理II途径,其铁同位素组成表现出了独特的特征,这也表明铁在水稻内的转运过程有其特殊性。Arnold et al.(2015)分别测定了水稻在有氧和厌氧(落干和淹水)生长条件下茎叶和种子的铁同位素组成。茎叶的Fe值分别为-0.32‰(厌氧)和-0.40‰(有氧),种子的Fe值分别为-0.39‰(厌氧)和-0.23‰(有氧)。虽然同位素组成在两种培养条件下没有显著的变化,但是在分馏趋势上可以观察到微小变化:厌氧培育时,水稻种子相对于茎叶富集轻铁(Δ56Fegrain-shoot=-0.06‰),而在有氧培育条件下,种子相对于茎叶富集重铁(Δ56Fegrain-shoot=0.14‰)。造成这种微小差异的原因可能是制样和测定的误差导致的,也有可能是水稻的生长条件改变导致的。考虑到土壤中铁形态的改变可能诱导水稻采用不同的铁吸收机制,且本研究中缺少水稻植株的铁浓度变化数据,无法得知培养条件的改变是否影响水稻植株对铁元素的获取,因此进一步更加详尽的研究还有待开展。
Garnier et al.(2017)详细测定了土壤-水-水稻体系的铁同位素组成,揭示了水稻独特的铁同位素组成,如图3所示。不同于以往机理II植物少量富集轻铁的研究结果,水稻植株内部铁的分馏可以达到-0.821‰,暗示水稻内铁的吸收转运机制可能不同于典型的机理 II植物(燕麦),可能是因为水稻可以像机理I植物一样吸收Fe2+,拥有独特的铁吸收转运过程。土壤-水-水稻体系中,铁同位素组成最轻的为孔隙水,Fe=-1.377‰,同位素组成最重的为铁膜,Fe=0.867‰,而植物整体偏轻,猜测铁膜作为铁库为水稻提供铁营养,且轻铁进入水稻而重铁留在铁膜中(图 1中的过程④)。铁膜活化主要依靠螯合机制,因而机理II可能是水稻吸收铁的主要机制。铁膜的主要矿物成分为水铁矿,铁膜与孔隙水之间的分馏值 Δ56Feiron plaque-Fe(II)aq2.24‰,这个分馏值远远大于文献报道的水铁矿沉淀导致的分馏值1.5‰(图1中的过程②),同时亚铁氧化形成水铁矿物的分馏值也在 1.5‰左右,似乎铁膜的形成无法用单一的机理来解释,可能涉及还原、氧化和沉淀等多个过程。水稻植株中,铁同位素组成最轻的为种子,δ56Fe=-0.462‰,铁从根到达种子的过程中,会经历配体和价态的双重变化,正是这些变化过程导致了铁同位素在水稻内的分馏,因此清楚铁在水稻内的形态变化过程以及这些过程导致的分馏效应对于解释水稻铁同位素组成变化显得十分重要(参考图1)。
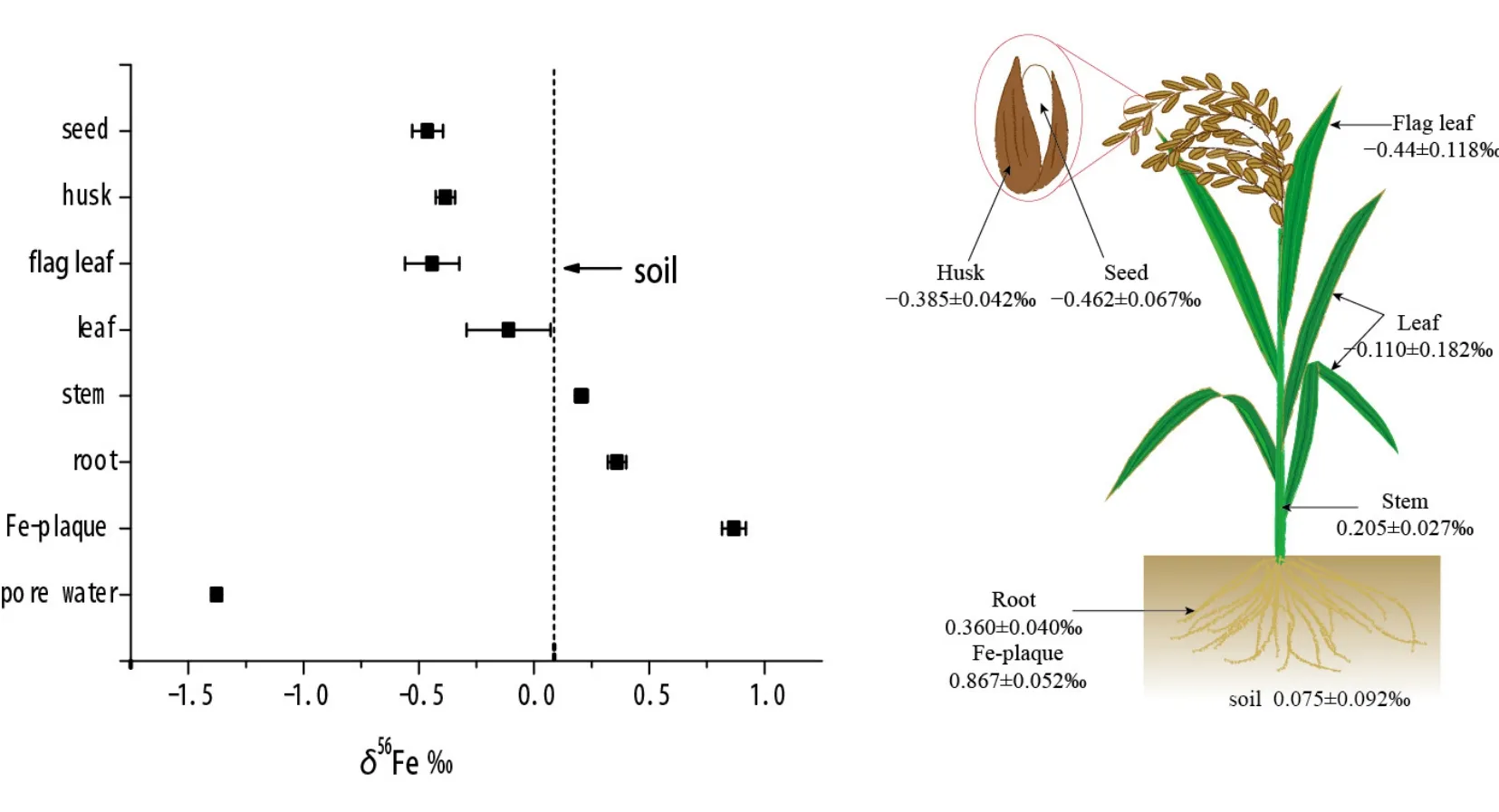
图3 土壤-水-水稻体系铁同位素组成(引自Garnier et al., 2017)Fig. 3 Iron isotope composition of soil-water-rice system (Cited from Garnier et al., 2017)
本研究初次较全面地测定了土壤-水-水稻体系的铁同位素组成,为揭示铁的循环过程提供了新的角度。但是本研究只是孤立地测定了水稻各组织的铁同位素组成,为了能更好地理解某一转运过程(如根与茎)导致的铁同位素分馏效应,应该运用质量平衡计算的方法来计算这一过程的铁同位素分馏效应,从而理解其铁转运机理。同时为了更加深入得解释水稻铁同位素机制,还可以结合水稻内诸如铁形态和价态的变化等生理过程来解释铁同位素数据,具体分析过程如下:
结合Moynier et al.(2013)的理论计算结果,仅改变铁元素所螯合的配体就可以导致 1.5‰的分馏:且 PS(phytosiderophore)、NA(nicotianamine)和 citrate间螯合重铁的能力依次为PS>NA>citrate。而配体改变加上价态变化,分馏效应可以达到3‰:Δ56FeFe(II)理论计算结果仅代表单个的转运过程,但是在水稻等植物中,运输过程往往都是复合转运过程,所以实际测定的分馏值代表的是多个转运过程的累积分馏效应。且水稻各器官是开放体系,也就意味着转运过程中导致的分馏效应不会被一直保存在转运的器官中,各个器官记录的是一定时空尺度内的累积分馏效应。已有的研究表明水稻木质部中铁主要以铁(III)-柠檬酸复合物形式存在,铁从木质部运输到韧皮部需要依靠NA作为穿梭体(Von Wiren et al.,1999)(图1中的过程⑨),由于NA相对于柠檬酸螯合重铁同位素(Δ56FeFe(II)-nicotianamine-Fe(II)-citrate≅1‰),因此进入韧皮部的为重铁。铁在植物内从根运往地上部的过程中,节和叶中会存在木质部和韧皮部之间的铁转运过程,且重铁进入韧皮部,轻铁留在木质部。水稻进入生殖生长阶段后,水稻根系退化,新叶和种子的铁主要通过韧皮部由老叶供给,特别是种子由于几乎没有蒸腾拉力作为动力,铁无法通过木质部获得,几乎全部通过韧皮部获得(图1中的过程⑩)。老叶中的铁主要以铁(III)-ITP复合物形式存在,活化进入新叶和种子的过程需要柠檬酸和尼克酰胺可以作为配体去运输铁,而ITP相对柠檬酸和尼克酰胺明显富集重铁,因此从老叶重新活化进入新叶和籽粒的铁为轻铁。特殊的是,PS作为一种具有强螯合能力的配体,只存在于机理II植物中,这也许能解释为何机理II植物相对机理I植物更加富集重铁。
4 展望
铁同位素方法为研究稻田体系铁的生物地球化学行为提供了全新的手段。铁同位素分馏行为可以反映水稻土的发育过程和特征,可以用来反演水稻土铁的生物地球化学过程,为土壤-水稻间以及水稻内部铁元素迁移机制提供了重要的科学依据。已有的研究表明氧化还原过程对土壤铁同位素组成的影响最大,但是在特定的土壤中,Eh变化、有机质含量、总铁含量的影响都可以超过氧化还原过程,因此解释土壤铁同位素组成时,应测定其主要化学性质,了解其发育背景,完善铁同位素分馏机制的理解。但是,要将铁同位素方法成熟地应用于铁生物地球化学行为研究还有诸多问题需要解决。
(1)清楚各个储库的铁同位素组成和同位素分馏机理是用铁同位素方法失踪铁地球化学行为的基础。目前已经测定了稻田体系的铁同位素组成,也摸清了大量的铁同位素分馏过程和机理。但是水稻各部分的铁同位素值只是得到了初步测定,且由于铁在水稻各部位的形态存在差异,因此如何制备具有代表性的植物样品对于解释铁的转运过程具有重要意义。例如在日后的研究中,应该尽量细致地测定具有同一形态铁的组织,例如分离叶片的叶肉细胞和叶脉;分别提取茎中的木质部和韧皮部汁液进行铁同位素值得测定。
(2)目前只是测定了水稻各部位的铁同位素值,但是这只能代表一定时间内累计的铁同位素分馏效应。为了更好地理解不同组织之间铁转运过程导致的铁同位素分馏效应,还应该利用质量平衡计算的方法来理解组织之间单个转运过程的铁同位素分馏机制。
(3)水稻铁同位素分馏效应除了受到平衡分馏过程的影响外,也有可能受到动力学平衡过程的影响。因此为了计算稻田体系动力学分馏效应,可以通过测定不同生长期的水稻样品(不同时期的水稻吸收的铁的比重不一样)的铁同位素组成来判断水稻铁同位素组成是否受到生长过程的影响,且可以计算动力学分馏系数。
(4)由于铁是变价元素,除了价态的变化,还会伴随配体改变等形态的变化。多个氧化还原过程的叠加,以及铁存在形态的改变,均会加深我们理解其分馏机理的难度。因此日后的研究应该集合同步辐射、红外光谱分析等手段来分析铁在水稻内的形态和价态,从而为铁同位素分馏机制提供多个角度的分析。
