不同放牧方式下的草场植被群落特征及其与土壤因子的关系
——以新巴尔虎左旗为例
2019-07-19周建琴田赟吴雨晴刘建康张克斌
周建琴,田赟,吴雨晴,刘建康,张克斌
水土保持国家林业局重点实验室/林业生态工程教育部工程研究中心/北京林业大学,北京 100083
放牧作为草地最简单而又有效的利用方式,其实质是一种生态干扰(Mcintyre et al.,1999)。放牧干扰可以通过改变土壤理化性质,增加生境的异质性,从而造成草地群落物种分布、结构和多样性格局发生变化,进而对整个生态系统的结构和功能产生影响(Keeley et al.,2003;Hoop er et al.,2005;韩路等,2015;张小红等,2017)。国内外诸多学者研究了不同干扰方式对植物群落的影响,研究发现,不同放牧措施对草地植物群落的影响存在差异(Ruifrok et al.,2014)。禁牧是恢复退化草地生态系统的有效途径,但长期禁牧造成牧草资源的浪费(肖金玉等,2015);中度放牧和短期禁牧有利于物种多样性的维持(宋向阳等,2018);休牧可提高草原生产力,对遏制草地退化、恢复草地功能起到了积极的作用(杨勇等,2015);连续放牧会导致植被盖度、高度、密度和生物量的大量下降,而适度放牧可以促进植被生长,增加地上生物量(Hejcman et al.,2010;Pucheta et al.,2010)。
物种组成和物种多样性指数是最能反映生态系统退化/恢复程度的生态学指标,它标示着群落内部及周围环境关系的变化(朱珣之等,2007;赵丽娅等,2018)。探讨草地群落物种多样性变化规律,对评价植被恢复状况和制定合理的草地管理措施具有重要的科学意义。植被的空间分布是植物与环境相互作用、共同发展的结果(钟娇娇等,2019)。研究植被分布与环境因子之间的关系可为生态系统功能的恢复和重建提供理论科学依据(Zuo et al.,2012;崔丽娟等,2013)。目前,数量分析是研究植被格局的重要生态手段,也是能揭示植被与环境因子之间复杂生态关系的较为合理有效的途径(Khan et al.,2016;周欣等,2015;王慧敏等,2018)。其中,排序是研究群落生态关系的重要数量学方法,通过排序研究能够更好地解释植物群落的空间分布与环境因子之间的关系,同时也能反映不同区域所在生境的环境因子差异(张金屯,2011)124-127。
呼伦贝尔草原位于中国北方干旱半干旱区,由于长期的超载过牧,造成人-草-畜关系失衡,草地退化成为制约当地草地畜牧业持续发展及生态环境改善的重要因素之一(彭飞等,2017)。2005年,新巴尔虎左旗实施了以转变传统畜牧业放牧方式为主要内容,以生态良好及草畜平衡为目标,以退化草地生态修复后草地资源高效利用为核心的“七个一”家庭生态牧场建设工程(杨威等,2013)。建设要求以家庭为单位在草牧场严重退化的草地实行全年禁牧(植被盖度小于 30%),一般退化的草地(植被盖度在30%-70%左右)实行牧草返青季节或结实期休牧、轮牧。目前,针对生态家庭牧场不同放牧方式对退化草地植被恢复及生态系统结构和功能的影响仍缺乏系统研究。本文通过研究“七个一”生态家庭牧场 4种放牧方式(禁牧、休牧、轮牧和连续放牧)下植被群落特征变化及植被-土壤因子之间的相互关系,阐明不同放牧方式之间的差异及环境因子对植被分布的影响,揭示不同的放牧方式与植被恢复之间的关系,探明可持续草地牧业管理方式,旨在保护草地生物多样性,促进已退化草地植被恢复。
1 研究区概况

图1 研究区位置图Fig. 1 Location of the study area
新巴尔虎左旗地处呼伦贝尔草原西南腹地,是呼伦贝尔市牧业四旗之一,地理坐标 117°33′-120°12′E,47°10′-49°47′N(图 1)。全旗地势东南高,西北低,平均海拔700 m,总面积18380000 hm2。气候属于温带半干旱大陆性季风气候,年均气温 0.3 ℃,极端最高气温 39.5 ℃,极端最低气温-47.8 ℃,年日照3076 h,无霜期100 d左右,积雪期140 d左右,年主导风向为西北风,年平均风速为2.8 m·s-1。旗内降水集中在7-8月,年均降水量287 mm,年均蒸发量2650 mm。土壤有栗钙土、黑钙土、暗灰色森林土3个地带性土类和草甸土、风沙土、沼泽土、粗骨土、碱土、盐土等6个隐域性土类。草原植被分布较广,代表性牧草有羊草(Leymus chinensis)、大针茅(Stipa grandis)、糙隐子草(Cleistogenes squarrosa)、雀麦(Bromus japonicus)、冷蒿(Artemisia frigida)、紫花苜蓿(Medicago sativa)、赖草(Leymus secalinus)等;可食用的植物有金针(Hemerocallis fulva)、山丹(Lilium pumilum)、蒙古韭(Allium mongolicum)等;野生药物植物有甘草(Glycyrrhiza uralensis)、黄芪(Astragalus membranaceus)、列当(Orobanche coerulescens)、草麻黄(Ephedra sinica)、柴胡(Bupleurum chinense)等。
2 研究方法
2.1 样地设置
根据对新巴尔虎左旗草原总体退化现状及草牧场经营状况的调查,选择当地满足“七个一”生态家庭牧场要求,且牧业方式管理较好的典型牧场一处(48°19′32″-48°19′06″N,118°35′59″-118°36′55″E)。牧场属于羊草+大针茅典型草原,土壤为栗钙土,地形地貌不存在较大的起伏,土壤类型基本一致。通过调查得知,该牧场2012年之前为统一的自由放牧草地,放牧强度一致,之后实行禁牧、休牧、轮牧和连续放牧。不同放牧方式区相邻,边界采用铁丝网围栏。牧场总面积600 hm2,其中禁牧区200 hm2,禁牧时间为2012-2017年;休牧区67 hm2,实行夏秋两季休牧制度(5-9月,9月后视植被生长状况打草并作为冬春季牧场);轮牧区200 hm2,划分为2块,面积各占一半,全年在2块轮牧区交替放牧,每次放牧时间1个月;连续放牧区133 hm2,全年放牧。经过调查和计算,休牧区冬季放牧、轮牧区和连续放牧区全年放牧的放牧强度为2.40-2.52个羊单位/hm2。本研究在4种不同放牧方式区各设置2个定位观测样地,其中2块轮牧区各设置1个,调查时间为2017年7-8月。
2.2 试验设计与测定指标
在8个观测样地各设置间隔50 m的3条样线,作为3次重复,每条样线设置10个样方(1 m×1 m),样方间隔30 m,共计调查样方8×30个。在每个样方内调查植物物种、株数、高度、盖度和鲜重,在样方内分种齐地面剪下每个物种地上绿色部分,装入信封,在 70 ℃烘箱中烘至恒重。同时,在每个样方内测定土壤含水量(一周内每天 8:00-10:00取样分别测定,各 3次重复)、土壤紧实度和土壤容重,并用土钻分别采集0-20 cm土层土样。在实验室测定土壤有机质(重铬酸钾容量法-稀释热法)、全氮(凯氏定氮法)、全磷(NaOH熔融-钼锑抗法)、速效磷(0.5 mol·L-1NaHCO3法)、和速效钾(NH4OAc-火焰光度计法)和EC值(电导率法)(鲍士旦,2007)。
2.3 数据处理
分别计算样方重要值和植被群落多样性指数和群落相似系数,其中群落多样性指数采用Margalef物种丰富度、Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数。相关指标计算公式如下:
重要值:

式中,RB为相对生物量;RC为相对盖度;RD为相对多度;RF为相对频度;RH为相对高度。
Margalef物种丰富度指数:

Shannon-Wiener多样性指数:

Simpson优势度指数:

Pielou均匀度指数:

式中,S为样地物种数;Ni为调查样方中第 i种物种的重要值;N为调查样地中所有物种的重要值之和;Pi为第i个物种的相对重要值。
Sorenson群落相似系数(Cs):

式中,a为样地A的物种数;b为样地B的物种数;c为样地A和B共有的物种数。
4种不同放牧方式区的物种组成和多样性指数间的差异采用多响应置换过程分析方法(MRPP:multi-response permutation procedures)进行检验,该方法是一种用于两个或多个样本的差异性分析的非参数检验方法,并不要求数据呈正态分布和具有方差齐性,因此被广泛应用在与生态学相关的研究中(Zimmerman et al.,1985;Stallins et al.,2002)。物种与土壤因子的排序采用典范对应分析(CCA:canonical correlation analyses)来完成。植物群落多样性与土壤因子的相关分析采用 Spearman秩相关分析法(张金屯,2011)40-42。物种重要值的计算及物种-重要值曲线绘制在 Excel软件中进行,MRPP检验和 CCA分析均在生态群落多元统计分析软件 PC-ORD 5.0中进行,秩相关分析在 SPSS软件中进行。
3 结果与分析
3.1 不同放牧方式下物种组成及群落多样性指数分析
研究区4种不同放牧方式区共有物种78种,其中轮牧区45种,禁牧区37种,连续放牧区34种,休牧区 42种,生长生态型多以多年生旱生、中旱生为主。在不同放牧方式区,物种的重要值分布有所不同(表1),但都以莎草科和禾本科为优势科,以寸草苔(Carex duriuscula)的重要值为最高,其中禁牧区为最高(IV=38.30)。此外,轮牧区以菊科(Asteraceae)、百合科(Liliaceae)和蔷薇科(Rosaceae)为主;禁牧区以百合科、十字花科(Brassicaceae)为主;连续放牧区以蔷薇科、菊科、藜科(Chenopodiaceae)为主;休牧区以豆科(Leguminosae)、鸢尾科(Iridaceae)和罂粟科(Papaveraceae)为主,其次为蔷薇科、紫草科(Boraginaceae)等。不同放牧方式区物种-重要值曲线(图2)说明,除物种序前10物种外,其他物种的重要值在物种序上的变化趋势相对较为平缓。休牧区物种重要值在物种序上的变化趋势相对平缓,禁牧区较为倾斜。物种序曲线趋势越倾斜或越急陡则表示物种分布的不均匀程度越高(Blackham et al.,2004)。休牧区物种重要值在物种序上的变化趋势相对平缓,禁牧区较为倾斜,表明休牧区物种分布较为均匀,禁牧区物种分布的不均匀程度较高。
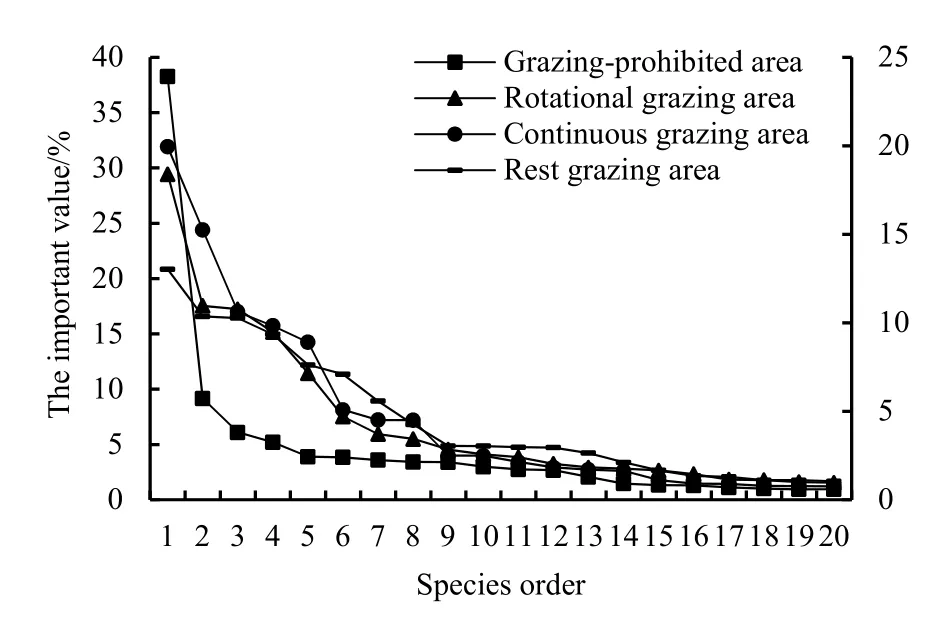
图2 不同牧业放牧方式区重要值-多样性曲线Fig. 2 Important value-diversity curve of species of different grazing management area

表1 物种重要值Table 1 Important value of species
不同放牧方式区的植被群落特征指数如表2所示。植被盖度和地上生物量呈现出相同的规律,都表现为禁牧区>休牧区>轮牧区>连续放牧区,其中,禁牧区植被盖度和地上生物量显著大于其他区(P<0.05)。物种丰富度指数Ma、物种多样性指数H和生态优势度指数C在4个区都表现为轮牧区>休牧区>连续放牧区>禁牧区,均匀度指数Jsw在4个区内相差不大,休牧区和轮牧区的均匀度指数相对较高。
样地内物种的MRPP分析结果(表3)表明,轮牧区和禁牧区、禁牧区和连续放牧区、连续放牧区和休牧区之间存在显著差异(P<0.05),轮牧区和连续放牧休牧区、连续放牧和休牧区之间差异不显著。T值是表示组间分离程度的统计量,其绝对值越大,则组间分离越强。轮牧区和休牧区的T值绝对值最小,表示组间分离程度较低,且组内同质性也较低。对植被群落多样性的MRPP分析结果表明,轮牧区和禁牧区的丰富度指数Ma和物种多样性指数 H之间存在显著差异(P<0.05),休牧区和连续放牧区之间的优势度指数 C存在显著差异(P<0.05),禁牧区和休牧区之间的均匀度指数 Jsw之间存在显著差异(P<0.05)。结合不同放牧方式区内物种重要值分布整体来看,禁牧区的物种分布及多样性与其他区均存在显著差异(P<0.05)。不同放牧方式区的相似性系数计算如表4所示,轮牧区与连续放牧区的相似性指数最高,其次为禁牧区与轮牧区。说明轮牧区和连续放牧区在植被恢复演替过程中,主要植物种类组成变化不大。

表2 不同放牧方式区的植被群落特征指数Table 2 Vegetation community characteristics of different grazing management area

表3 物种组成MRPP分析结果Table 3 Result of multi-response permutation proceduresof species composition
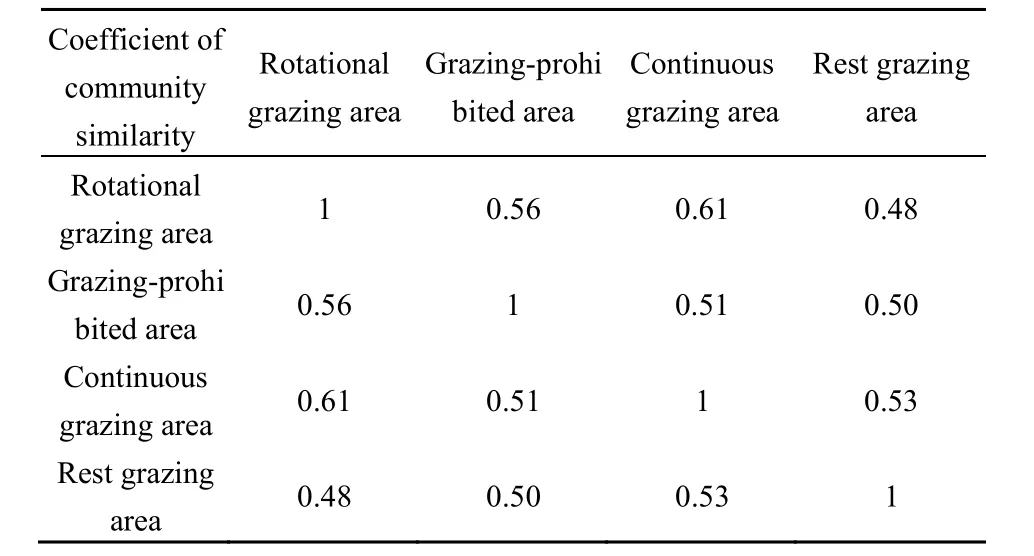
表4 相似性指数Table 4 Similarity indexes
3.2 植被分布与土壤因子排序分析
由表5可知,物种CCA排序轴与土壤因子相关。前3个排序轴的累积贡献率为88.3%,其中前两轴的累积贡献率达74.7%,说明CCA前两轴已能较好地反映研究区植物群落与环境之间的关系(Scudeller et al.,2001)。在前两个排序轴中,物种与环境的相关系数分别为 0.85、0.78,前两轴特征根值之和占总特征根值的71.2%。如果前3个排序轴的物种与环境关系方差累计贡献率大于40%,则排序结果较准确(Gauch,1982)。因此,本文的排序结果能较好地解释物种分布与土壤因子之间的相关关系。
由表6和图3可知,所选取的9个土壤因子对植被分布都有不同程度的影响。第一排序轴与土壤EC值呈显著正相关(r=0.68),与土壤紧实度和含水量呈显著负相关(r=-0.90,r=-0.62),说明沿着第一排序轴从左到右,土壤水溶性盐含量越来越高,含水量和容重越来越小;第二排序轴与土壤全磷含量和紧实度呈显著正相关(r=0.88,r=0.57),与土壤含水量呈显著负相关(r=-0.94),说明沿着第二轴从上至下,土壤全磷含量和紧实度逐渐降低,土壤含水量逐渐增大。另外,土壤容重、含水量和全磷含量箭头连线最长,说明它们在排序轴上的位置很大程度上反映了其生态特点。综合分析可知,影响左旗植被分布的主要土壤因子是容重、含水量和全磷含量。
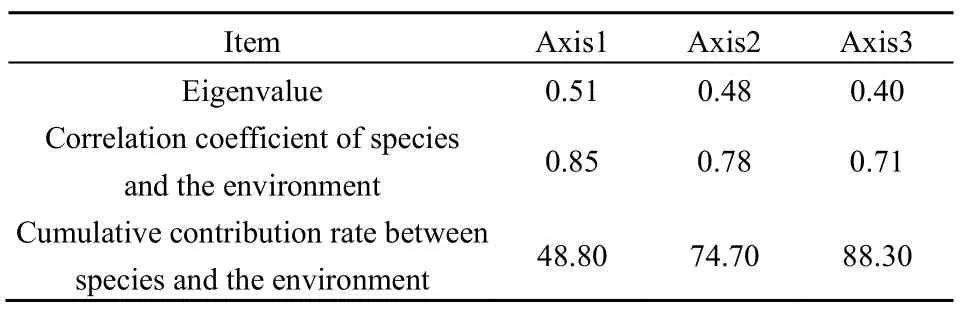
表5 CCA排序轴特征值及物种与环境的相关性Table 5 Eigenvalues and of CCA axes correlation coefficient between species and the environment
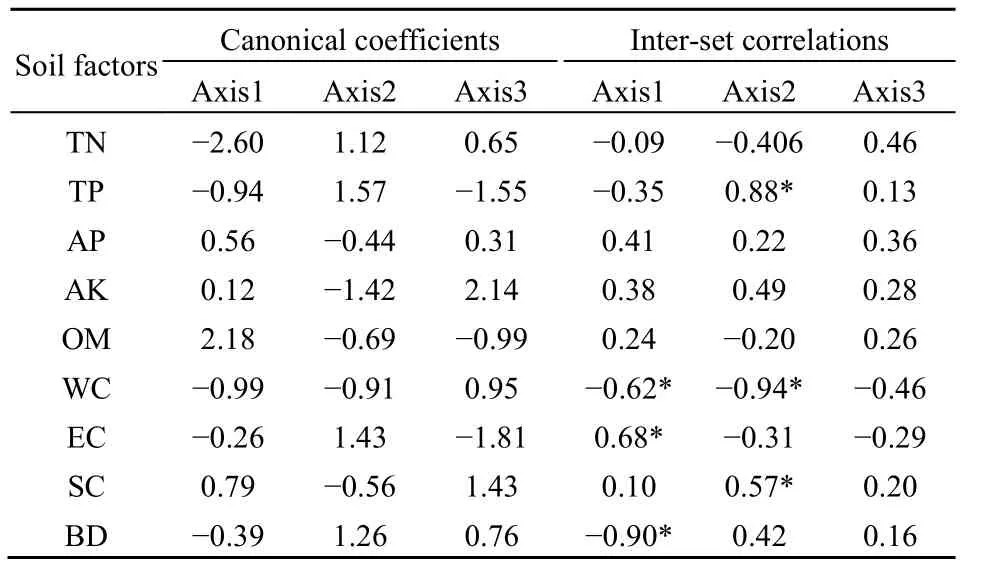
表6 土壤因子与CCA排序轴的典型系数和序列间相关系数Table 6 Canonical coefficients and inter-set correlations of soil factorsand axes of CCA

图3 植被与土壤因子的二维排序图Fig. 3 CCA two-dimensional ordination diagram of vegetation and soil factors
植被的分布也基本上反映了不同放牧方式之间的差异。由图3可知,与含水量因子密切相关的物种寸草苔(41)、芦苇(Phragmites communis,21)和狭叶黄芩(Scutellaria regeliana,37)等主要为禁牧区物种;与土壤容重密切相关的物种蚊母草(Purslane Speedwell,36)、星毛委陵菜(Potentilla acaulis,39)、漏芦(Stemmacanthauniflora,20)、冷蒿(18)和骆驼刺(Phragmites communis,22)主要分布在轮牧区和连续放牧区;艾蒿(Artemisia argyi,1)、北柴胡(Bupleurum chinense,2)和飞廉(Carduus nutans,13)与土壤全磷相关,主要分布在禁牧区;与土壤速效钾相关的物种花旗杆(Dontostemon dentatus,15)和羊草(4)在禁牧、轮牧和连续放牧区都有分布;与土壤EC值相关的植物骆驼蓬(Peganum harmala,23)、斜茎黄芪(Astragalus adsurgens,38)和糙隐子草(28)主要分布在休牧区,轮牧区和连续放牧区也有少量分布。这说明影响禁牧区物种分布的主要土壤因子为含水量和全磷,影响轮牧区物种分布的主要土壤因子为容重和全磷,影响休牧区物种分布的主要土壤因子为EC值,影响连续放牧区物种分布的主要土壤因子为容重和速效钾。
3.3 植被群落多样性与土壤因子间的相关分析
土壤因子与植被群落多样性指数的秩相关分析结果显示(表7),土壤速效磷含量与丰富度指数Ma呈显著正相关(r=0.72,P=0.03),土壤含水量与物种多样性指数 H呈显著正相关(r=0.66,P=0.04),土壤容重与丰富度指数Ma呈显著负相关(r=-0.76,P=0.03),与均匀度指数 Jsw都呈显著正相关(r=0.80,P=0.002),生态优势度指数C与全磷含量呈显著正相关(r=0.68,P=0.03),与土壤EC值呈显著负相关(r=-0.64,P=0.04)。该分析结果与物种与土壤因子排序分析的结果有相似之处。
4 讨论
4.1 不同放牧方式对物种组成及群落多样性的影响
放牧干扰是影响草原植被群落特征,造成草地退化的主要因子,本研究中,4种放牧方式区优势物种都以莎草科和禾本科为主,生长型多以多年生旱生、中旱生为主,各区都以寸草苔的重要值为最高。寸草苔是多年生草本,生态幅度广,耐寒耐旱耐盐碱,在草原退化时可变为优势植物。因此,研究区草地仍属于退化草地,其植被的恢复还需要很长的过程。禁牧是恢复退化草地生态系统的有效途径(董乙强等,2018)。短期禁牧有利于维持较高的物种多样性和较好的土壤物理性质(敖伊敏等,2011;宋向阳等,2018),但长期禁牧反而对植物生长有抑制作用(Ruifrok et al.,2014),随着禁牧时间的延长,植被会出现衰退现象,物种多样性下降(Peco et al.,2005)。这与本研究结论相似。本研究禁牧区禁牧时间在5年左右,其植被盖度及生物量均最高,说明禁牧对退化草地植被恢复及生产力提高有显著促进作用。但禁牧区物种丰富度指数、物种多样性指数和生态优势度指数均最小,物种均匀度指数最低,禁牧区与其他区的物种分布及多样性均存在显著差异。原因是适度放牧条件下,许多物种可以共生,而禁牧区缺乏放牧干扰,物种间的竞争关系发生改变,少数竞争力强的物种,如寸草苔,成为群落的优势物种,抑制其他物种生长,导致物种多样性降低。研究发现,长期单一的禁牧反而使草地生态系统呈现退化趋势,在中国北方干旱半干旱草地,围封一段时间后,适当的割草或放牧有利于加速退化草地恢复(李学斌等,2013;刘小丹等,2015);呼伦贝尔草地封禁4-7年后,进行适当割草及放牧利用可改变牧草植物的组成并改善牧草质量,提高牧草的生产量,促进牧草再生(李怡等,2011;刘美丽,2016);相对于禁牧,刈割更有利于维持该地区生态系统的多功能性(李静鹏等,2016)。因此,本研究区的禁牧区,目前缺乏一定的干扰来维持较高的生物多样性,建议实行刈割、休牧或轮牧等放牧利用。
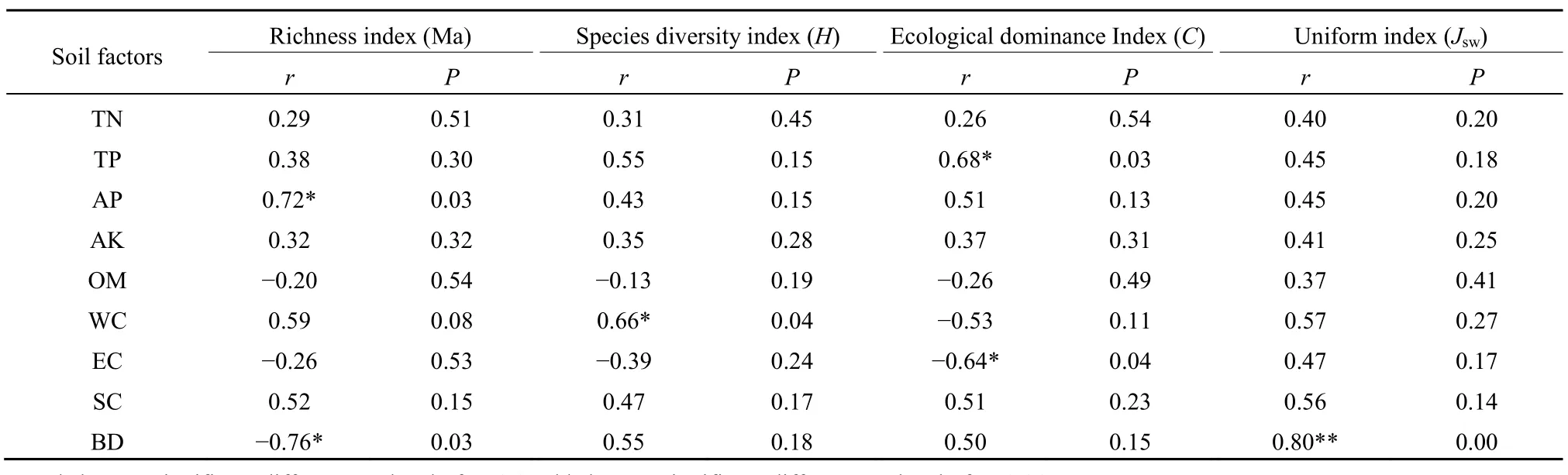
表7 多样性指数与土壤因子间的秩相关分析Table 7 Spearman rank correlation analysis of vegetation characteristics and soil factors
一般而言,群落在中度放牧强度条件下能够维持较高的多样性,并随着放牧强度的增加,草场逐步退化(蒙旭辉等,2009;刘佳慧等,2017)。本研究中,休牧区休牧时间为5个月,轮牧区轮牧时间为1月。休牧区物种分布较为均匀,休牧区和轮牧区的物种多样性指数及群落均匀度指数较高,物种MRPP结果显示它们之间无显著差异,而连续放牧区由于长期的放牧压力,羊群啃食和践踏,草地趋于干旱化,各物种竞争关系发生改变,抑制优势物种对草地资源的竞争,旱生植物逐渐增多。虽然耐牧草种(耐践踏、耐放牧)有所增加,但由于过度放牧,物种总数有所降低。连续放牧区放牧频率过高和禁牧区封禁时间过长无干扰均对植被恢复起抑制作用,研究结果符合“中度干扰”理论(Osman,2008)。适度放牧是保护植物功能群、物种多样性,维护生态系统基本功能和健康状态的有效途径,科学合理的放牧机制是草地资源可持续利用和畜牧业健康发展的长久之计。
4.2 植被分布及群落多样性与土壤因子的关系
生态系统中,生物与环境是两个相互影响、相互制约的要素(王蒙等,2015),排序是用来分析植被与环境之间生态关系的重要手段。本研究发现,土壤容重、含水量和全磷含量是影响左旗植被分布的主要土壤因子,土壤速效磷、含水量和容重对物种多样性指数有显著影响。国内外相关研究认为,土壤的容重受放牧家畜践踏程度的影响,动物践踏作用的增强,会导致土壤空隙分布的空间格局发生变化,土壤的总空隙减少,土壤容重增加(Trlica et al.,2002)。本研究轮牧区与连续放牧区,由于放牧改变植被覆盖度,大量动物对表层土壤的踩踏提高了土壤紧实度和容重,透气性变小,进而影响土壤水分的的存储和蒸发散,土壤结构发生变化,养分降低,植物生长受到抑制。因此,土壤容重成为该区影响植被分布的主要土壤因子。在干旱半干旱区,绝大多数群落依靠天然降水来维持其群落结构,国内外的大量研究均表明降水是决定草场结构和功能的决定性因子,生态系统的结构和功能在很大程度上依靠水分的平衡来维持(Ellis et al.,1994;杨晓晖等,2005)。新巴尔虎左旗近 3年较干旱,降水稀少,植物生长很大程度上受降水量的影响,而禁牧区植被分布的主要受含水量影响可能是因为禁牧区长时间无放牧干扰,增加了地表凋落物覆盖度,减少了水分的入渗,降低了植物对水分的有效利用,植物生长受到影响。其次,研究区草地退化,土壤全磷含量和速效磷含量均很低,养分贫瘠,加之土壤得不到雨水充分淋洗,蒸发强烈,致使盐分在土壤表层聚集,土壤条件不足以供应草地生长,因此,土壤全磷及EC值也成为影响植被分布的主要土壤因子。
5 结论
本研究以草地生态恢复学理论为指导,采用数量生态学的方法,通过对新巴尔虎左旗“七个一”生态家庭牧场禁牧、休牧、轮牧和连续放牧方式下的植被群落特征及其与土壤因子间的关系进行研究,得出如下结论:
(1)左旗植被以多年生旱生和中旱生植物为主,以莎草科和禾本科植物占比最多,其中寸草苔在4种放牧方式区的重要值都为最高,以禁牧区为最高。轮牧区和休牧区的物种数和群落多样性指数较高。连续放牧区植被盖度及生物量最低,禁牧区植被盖度和生物量最高,但禁牧区的群落多样性指数最低,且与其他放牧方式区的物种分布及多样性指数均存在显著差异。
(2)禁牧对植被盖度和生物量的提升有积极作用,但长期禁牧不利于维持较高的群落多样性,同时,放牧区放牧频率过高也不利于恢复退化草地生态系统结构和功能。总体而言,休牧和轮牧方式在退化草地恢复过程中优于禁牧和连续放牧。因此,建议对已经长期禁牧的牧场实行适度的割草或放牧利用,对植被退化严重,土壤沙化严重的连续放牧区,实施 3-4年的禁牧,进而根据恢复情况实施能保证植被恢复周期的牧业管理方式。
(3)土壤容重、含水量和全磷是影响新巴尔虎左旗植被分布最主要的土壤因子,且群落多样性指数的变化与容重、速效磷和含水量密切相关。在降水少、蒸发量大,土壤贫瘠的北方干旱半干旱牧业区,实施以草畜平衡和节水为基点的“七个一”生态家庭牧场模式非常有必要。然而,如何科学调整其管理模式,如放牧强度与放牧周期、土壤施肥与改良、草种补播等仍需要深入研究。
