西南山地小径竹的重金属富集能力
2019-07-19段晨松杨彪韩燕张全建龚旭张远彬
段晨松 ,杨彪 ,韩燕 ,张全建 ,龚旭 ,张远彬*
1. 中国科学院、水利部成都山地灾害与环境研究所,四川 成都610041;2. 中国科学院大学,北京 100049;
3. 西南野生动植物资源保护教育部重点实验室,四川 南充 637002;4. 西华师范大学,四川 南充 637002
中国土壤重金属污染形势总体不容乐观,随着城镇化和工业化水平的提高,西南地区的土壤Cu、Zn、Pb、Cd、Cr超标范围较大,迫切需要修复和防控土壤重金属污染,保障生态安全(周建军等,2014;陈能场等,2017;陆静,2018)。植物修复是近些年来应用于重金属污染治理的重要手段之一,尤其是利用超积累植物进行的修复,由于其重金属耐性高且富集能力强而广泛受到关注(Merchant et al.,2010;Li et al.,2018)。国际上已发现的重金属超积累植物有700多种,但大多数因适生范围狭窄、生物量小、富集总量少、修复周期较长,在实践中应用成功的事例较少,从而限制了超积累植物的应用与推广(Reeves et al.,2000;Yang et al.,2014;胡鹏杰等,2014;杜俊杰等,2018)。
竹类植物广泛分布于热带和亚热带(Das et al.,2008)。中国是世界上竹子资源最丰富的国家,竹类植物有50多属500余种(Zhou et al.,2011);竹林面积约有7.2×104km2,占中国森林总面积的3.5%。竹子因其一次造林后可永续连年利用,且生长迅速,生物量大,为社会提供了大量的竹材、竹笋和竹制品;并且,竹子种类繁多,具有很高的观赏价值,是生态环境建设和城市园林建设的优良树种(Kleinhenz et al.,2001)。目前,已有一些学者对竹类植物的重金属吸收转移规律进行研究,发现污染土壤中的重金属主要聚集在竹子地下部分(竹根、竹鞭等)(张志坚等,2011;Collin et al.,2013;Chen et al.,2015;Li et al.,2015;李娟等,2016;Bian et al.,2017;Were et al.,2017),这主要由于竹类植物具有错综复杂而发达的根系统,有利于吸收重金属元素(Li et al.,2016;Zhong et al.,2017);但同时竹子具有克隆生长的特性,其地上部分的收获能够更高效地转移土壤重金属,而目前对竹子地上部分,尤其是西南山地小径竹地上各部位的重金属积累鲜有研究报道。Collin et al.(2012)对印度洋留尼汪岛16种竹子茎和叶的Cu、Zn含量进行测定,发现叶的 Cu平均含量比茎高27.51%,Zn平均含量比茎高44.04%。也有学者对不同竹子在重金属胁迫下的生理生态响应进行了探讨,发现不同竹子对重金属的耐受程度不同。姜黎(2009)通过盆栽实验发现刚竹属竹子的Cu耐受性强于苦竹属的竹子;李娟等(2016)研究表明菲白竹(Sasa fortunei)对Pb的耐受性高于阔叶磐竹(Indocalamus latifolius)和黄条金刚竹(Pleioblastus kongosanensi);王兵(2011)通过综合评价金镶玉竹(Phyllostachys aureosulcata)和紫竹(Phyllostachys nigra)的重金属耐抗性,认为金镶玉竹的Pb耐受性高于紫竹、Cd的耐受性低于紫竹。但这多是对长江-南岭竹类植物的研究,西南山地竹区的小径竹对土壤不同重金属的富集能力如何、不同小径竹的富集能力是否存在差异,尚未见研究报道。
本文以西南山地森林群落重要的林下优势层片缺苞箭竹(Fargesia denudate)和拐棍竹(F.robusta)为研究对象,通过分析两种小径竹茎、叶、笋的Cu、Zn、Pb、Cd、Cr含量和富集系数,拟揭示小径竹地上各部位对不同重金属的富集能力,比较缺苞箭竹和拐棍竹的重金属富集差异;并通过与常见超积累植物对比,明确西南山地小径竹的重金属富集潜力,为进一步研究其重金属修复机理和应用提供科学参考和新的研究视角。
1 材料与方法
1.1 试验材料
本试验的缺苞箭竹及其根际土壤采自四川王朗国家级自然保护区,地理坐标为 103°55′-104°10′E、北纬 32°49′-33°02′N;拐棍竹及其根际土壤采自四川省卧龙国家级自然保护区,地理坐标为东经 102°52′-103°25′E、北纬 30°45′-31°25′N(图 1)。四川王朗自然保护区地处岷山山系腹心地带,总面积约323 km2,属于丹巴-松潘半湿润气候,年均温2.7 ℃,年降水量860 mm;土壤垂直分布规律明显,从低海拔到高海拔依次有山地棕壤、山地暗棕壤、亚高山草甸土、高山草甸土、高山流石滩荒漠;该地区植被保存较完整,在海拔梯度上分布有针阔混交林/落叶阔叶林(2300-2600 m)、亚高山暗针叶林(2600-3500 m)、亚高山灌丛草甸(3500-4400 m)、高山流石滩植被(4400-4900 m)、高山荒漠带(>4900 m)。四川卧龙自然保护区位于邛崃山脉东翼,面积约2000 km2,属于典型的亚热带内陆山地气候,年均温8.5 ℃,年降水量890 mm;土壤随海拔变化依次为山地黄壤、山地黄棕壤、山地暗棕色森林土、山地棕色暗针叶林土、高山草甸土、寒漠土等;植被垂直分布规律明显,在海拔梯度上依次为亚热带常绿阔叶林(1120-1600 m)、常绿/落叶阔叶混交林(1600-2000 m)、针阔叶混交林(2000-2600 m)、亚高山针叶林(2600-3600 m)、高山草甸(3600-4400 m)和流石滩植被(4400-5000 m)。
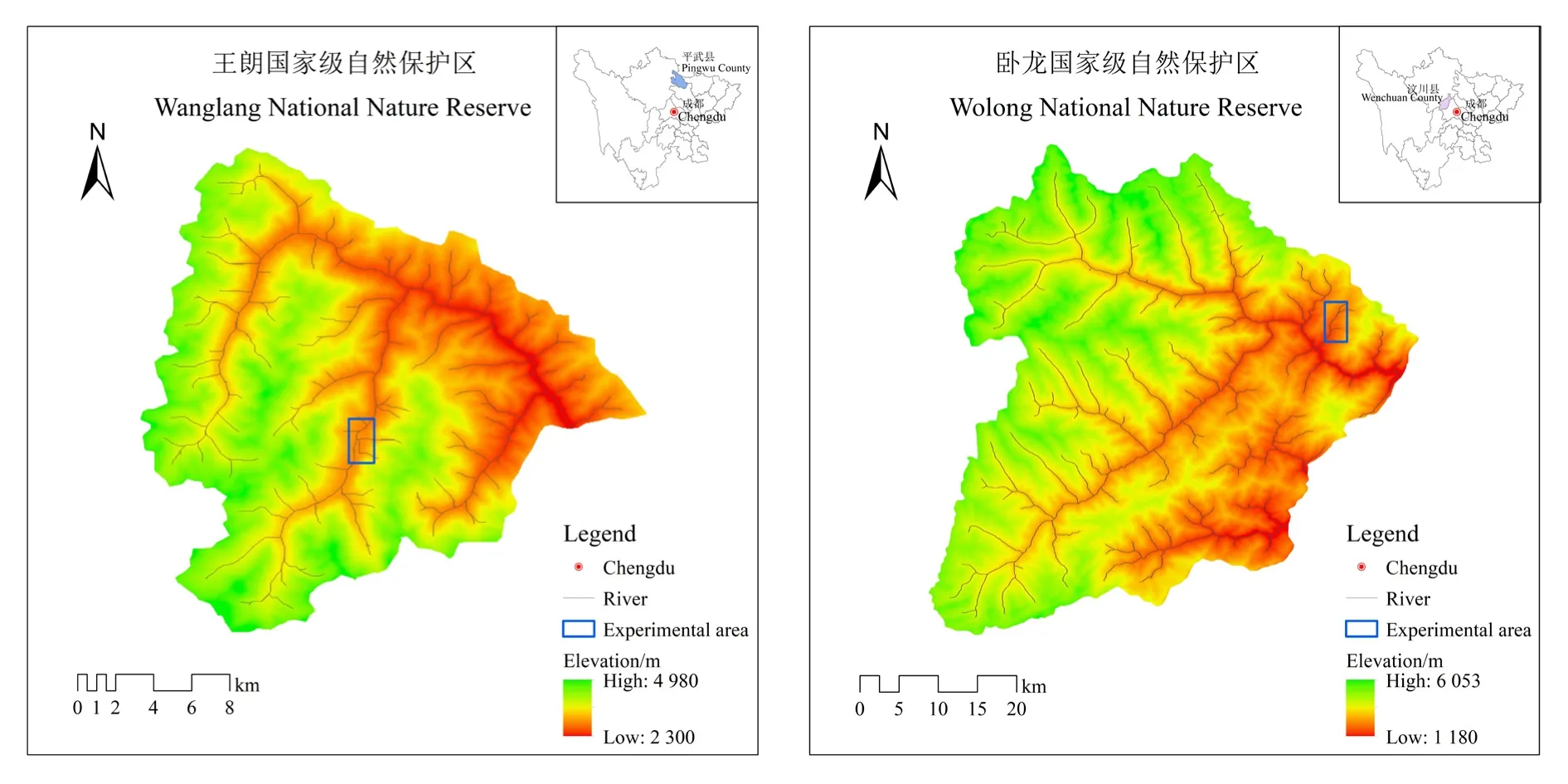
图1 研究区分布图Fig. 1 Distributions of experimental areas
缺苞箭竹和拐棍竹均是禾本科箭竹属的克隆植物。缺苞箭竹是亚高山森林群落中最具优势的层片,占岷山山系竹林面积的27.8%,在四川王朗国家级自然保护区主要分布于海拔1920-3200 m的针阔混交林和暗针叶林下;缺苞箭竹高3-5 m,直径0.6-1.3 cm,密度约24 plant·m-2,地上生物量约 9.75 kg·m-2(王开运,2004)。拐棍竹是邛崃山系重要的林下优势层片,占该山系竹林面积的21.4%,主要生长在卧龙国家级自然保护区海拔1700-2800 m的常绿/落叶阔叶混交林下;拐棍竹高3-5 m,直径1-3 cm,密度约8 plant·m-2,地上生物量约2.79 kg·m-2(易同培等,2008)。
1.2 样品采集和测定
2017年9月,在四川王朗和卧龙国家级自然保护区分别选择海拔、坡度、坡向、乔木树种、郁闭度、土壤类型等基本一致的20 m×20 m缺苞箭竹和拐棍竹样地各6个,各样地竹子长势基本一致、生长良好,且样地间的直线距离≥50 m(表1)。在每个样地随机设置3个1 m×1 m的样方,沿地表剪取样方内所有的竹笋和多年生克隆分株,称量其鲜重;然后将3个样方的茎、叶、笋分别混合均匀,取其鲜样各1 kg作为样品,将样品在105 ℃烘箱中杀青30 min,再于70 ℃烘至恒重,然后研磨、过筛、装袋,用于测定小径竹地上各部位的 Cu、Zn、Pb、Cd、Cr含量。同时,在每个样方内,取竹鞭上下5 cm处的土样500 g,经风干、研磨、过筛、装袋后,用于测定土壤的 Cu、Zn、Pb、Cd、Cr含量。
植物中的 Cu、Zn、Pb、Cd、Cr含量采用HNO3-HClO4消解-原子吸收分光光度法测定;土壤中的Cu、Zn含量用NaOH熔融-原子吸收光谱法测定,Pb、Cd含量用王水-高氯酸消煮-原子吸收光谱法测定,Cr含量用硝酸-过氧化氢分解-原子吸收光谱法测定。
1.3 数据处理
重金属富集系数是指植物某一部位的元素含量与土壤相应元素含量之比,是衡量植物将重金属吸收转移到体内的能力大小的一个重要指标,常用来反映植物对重金属的富集能力,其计算公式如下(时宇等,2018):
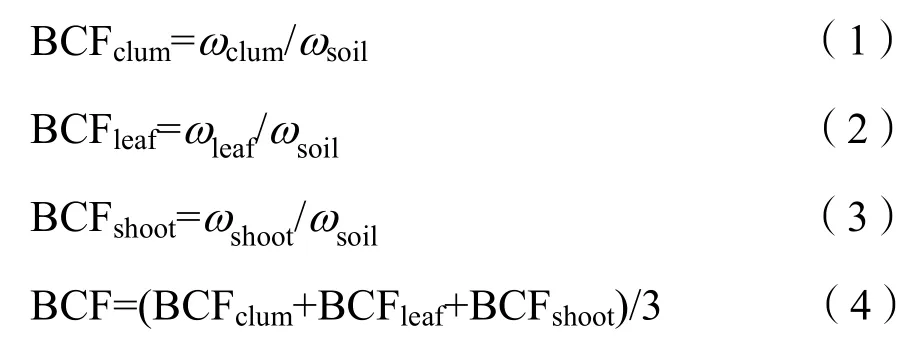
式中,BCFclum、BCFleaf、BCFshoot分别为小径竹茎、叶、笋的富集系数,BCF为地上部富集系数,ωclum、ωleaf、ωshoot分别为小径竹茎、叶、笋中的某种重金属含量,ωsoil为土壤中相应重金属的含量。
本文数据运用IBM SPSS Statistics 22进行分析,对缺苞箭竹和拐棍竹不同部位的重金属含量差异和富集系数差异做单因素方差分析(One-way ANOVA),并采用Tukey方法进行显著性检验;对缺苞箭竹和拐棍竹的地上部富集系数差异进行t检验。所有数据均用平均值±标准误表示,显著性水平为 α=0.05。
2 结果与分析
2.1 土壤重金属含量
缺苞箭竹和拐棍竹根际土壤的 Cu、Zn、Pb、Cr含量均低于土壤环境质量二级标准值,而Cd含量均高于土壤环境质量二级标准(0.30 mg·kg-1)。其中,缺苞箭竹根际土壤的 Cd含量最高为 1.25 mg·kg-1,平均为 0.54 mg·kg-1,超标率达 80.03%;拐棍竹根际土壤的Cd含量最高为0.39 mg·kg-1,平均为0.32 mg·kg-1,超标率为6.67%(表2)。
2.2 缺苞箭竹和拐棍竹的重金属含量
缺苞箭竹和拐棍竹叶中的 Cu、Zn、Pb、Cd、Cr含量均显著高于茎和笋(表3,P=0.001)。缺苞箭竹和拐棍竹叶中的最高 Cu含量分别为 80.30 mg·kg-1和 11.04 mg·kg-1,茎中的 Cu 含量均显著小于笋(表3,P=0.001);叶中的最高Zn含量分别为230.62 mg·kg-1和 72.80 mg·kg-1,缺苞箭竹笋中的Zn含量显著高于茎(表3,P=0.001),而拐棍竹茎和笋中的Zn含量差异不显著(表3,P=0.460);叶中的最高 Pb含量分别为 8.61 mg·kg-1和 10.26 mg·kg-1,缺苞箭竹茎中的Pb含量显著高于笋(表3,P=0.048),而拐棍竹茎和笋中的Pb含量差异不显著(表 3,P=0.150);缺苞箭竹和拐棍竹叶中的最高 Cd 含量分别为 0.35 mg·kg-1和 0.66 mg·kg-1,茎、笋中的Cd含量差异不显著(表3,P=0.481、0.511);叶中的最高Cr含量分别为121.72 mg·kg-1和19.51 mg·kg-1,缺苞箭竹和拐棍竹茎中的Cr含量均大于笋,但无显著性差异(表 3,P=0.942、0.773)。
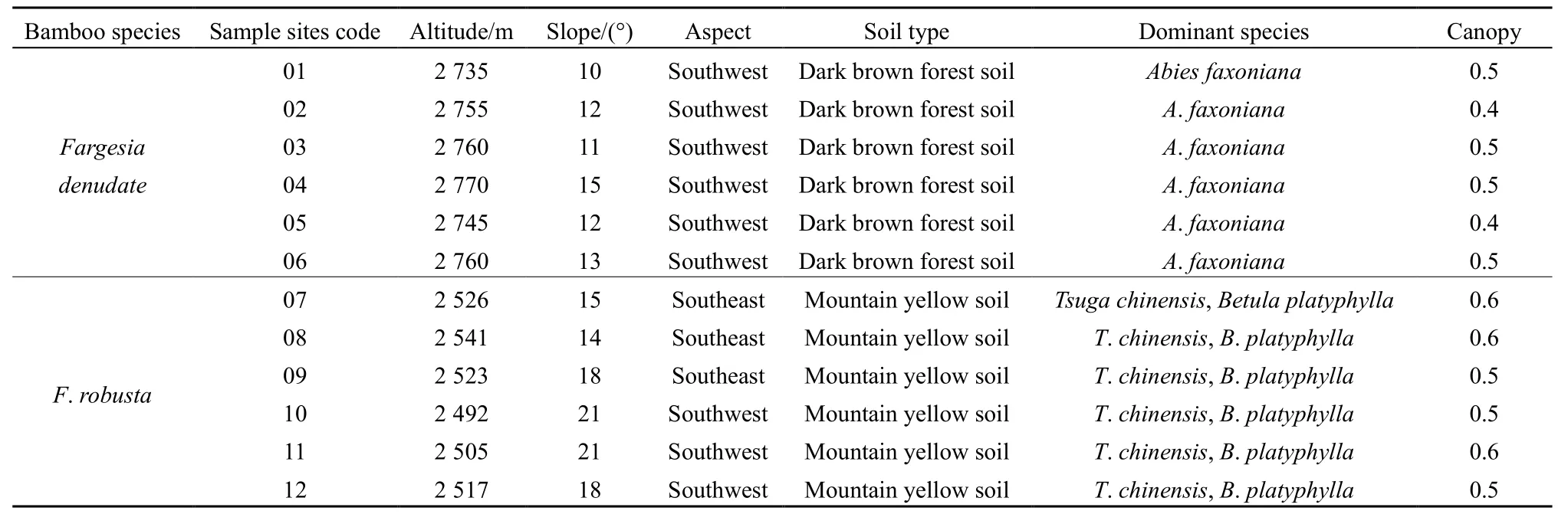
表1 样地基本信息Table 1 Situation of sample sites
不同重金属在两种小径竹地上各部位中的含量有一定差异,但均为Zn的含量最高,Cd的含量最低(表3)。缺苞箭竹地上各部位的重金属含量均呈现出 Zn>Cu>Cr>Pb>Cd(表 3);拐棍竹叶和笋的重金属含量呈现出 Zn>Cr>Cu>Pb>Cd,而茎的重金属含量表现为 Zn>Cr>Pb>Cu>Cd(表 3)。
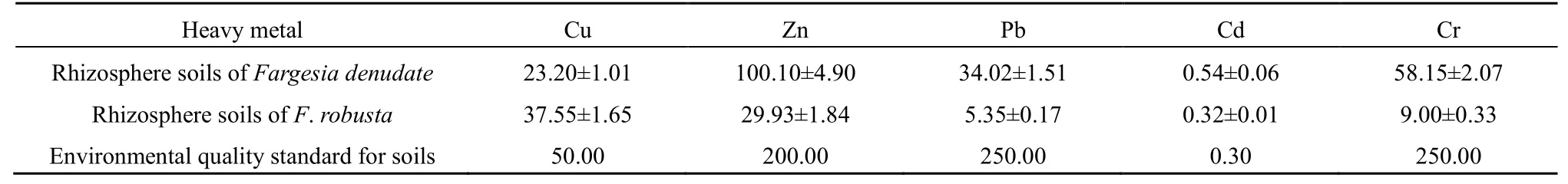
表2 缺苞箭竹和拐棍竹的根际土壤重金属含量Table 2 The contents of heavy metals in rhizosphere soils of Fargesia denudate and F. robusta mg·kg-1
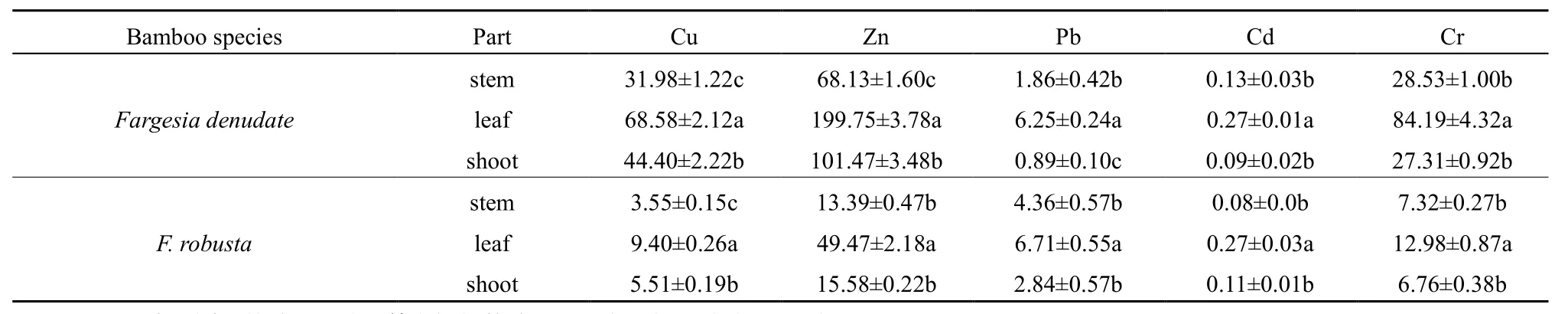
表3 缺苞箭竹和拐棍竹茎、叶、笋中的重金属含量Table 3 The heavy metals contents in the stems, leaves and shoots of Fargesia denudate and F. robusta mg·kg-1
2.3 缺苞箭竹和拐棍竹的重金属富集系数
缺苞箭竹和拐棍竹均表现出叶的富集系数显著大于茎和笋(图 2,P=0.001)。缺苞箭竹和拐棍竹叶的Cu最大富集系数分别为4.23和0.48,笋对Cu的富集系数显著大于茎(图2,P=0.013、0.012);叶的Zn最大富集系数分别为2.90和2.95,缺苞箭竹笋对Zn的富集系数显著大于茎(图2,P=0.001),而拐棍竹茎和笋的 Zn富集系数无显著性差异(图2,P=0.757);叶的Pb最大富集系数分别为0.32和2.05,茎和笋对Pb的富集系数差异不显著(图2,P=0.136、0.183);缺苞箭竹和拐棍竹叶的最大 Cd富集系数分别为1.13和2.19,叶的最大Cr富集系数分别为2.73和2.59,茎、笋对Cd和Cr的富集系数均无显著性差异(图2,P=0.760、0.947,0.497、0.876)。
不同小径竹对不同重金属的富集系数存在一定差异。缺苞箭竹地上部对Cu的富集系数大于Zn、Pb、Cd、Cr,而拐棍竹对Cu的富集系数较小(表4)。缺苞箭竹茎、叶、笋对重金属的富集系数依次为 Cu>Zn>Cr>Cd>Pb;拐棍竹茎的富集系数依次为Cr>Pb>Zn>Cd>Cu , 叶 的 富 集 系 数 依 次 为Zn>Cr>Pb>Cd>Cu , 笋 的 富 集 系 数 依 次 为Cr>Zn>Pb>Cd>Cu(图 2)。缺苞箭竹地上部及其各部位对Cu的富集系数显著大于拐棍竹(图2和表4,P=0.001),而对重金属 Pb的富集系数显著小于拐棍竹(图2和表4,P=0.002、0.001、0.001、0.002);缺苞箭竹地上部对 Zn的富集系数显著大于拐棍竹(表4,P=0.001),茎和笋均具有显著性差异(图2,P=0.001),而叶的差异不显著(图 2,P=0.062);缺苞箭竹地上部对 Cd的富集系数显著小于拐棍竹(表4,P=0.049),叶和笋均具有显著性差异(图2,P=0.030、0.016),而茎的差异不显著(图 3,P=0.956);缺苞箭竹地上部对Cr的富集系数显著小于拐棍竹(表 4,P=0.001),茎和笋均具有显著性差异(图2,P=0.001、0.002),而叶的差异不显著(图 2,P=0.967)。
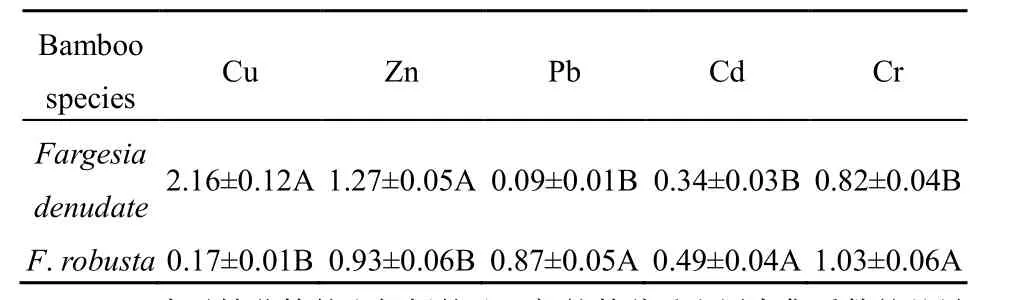
表4 缺苞箭竹和拐棍竹地上部的重金属富集系数Table 4 The heavy metals bioaccumulation factors in the aboveground part of Fargesia denudate and F. robusta
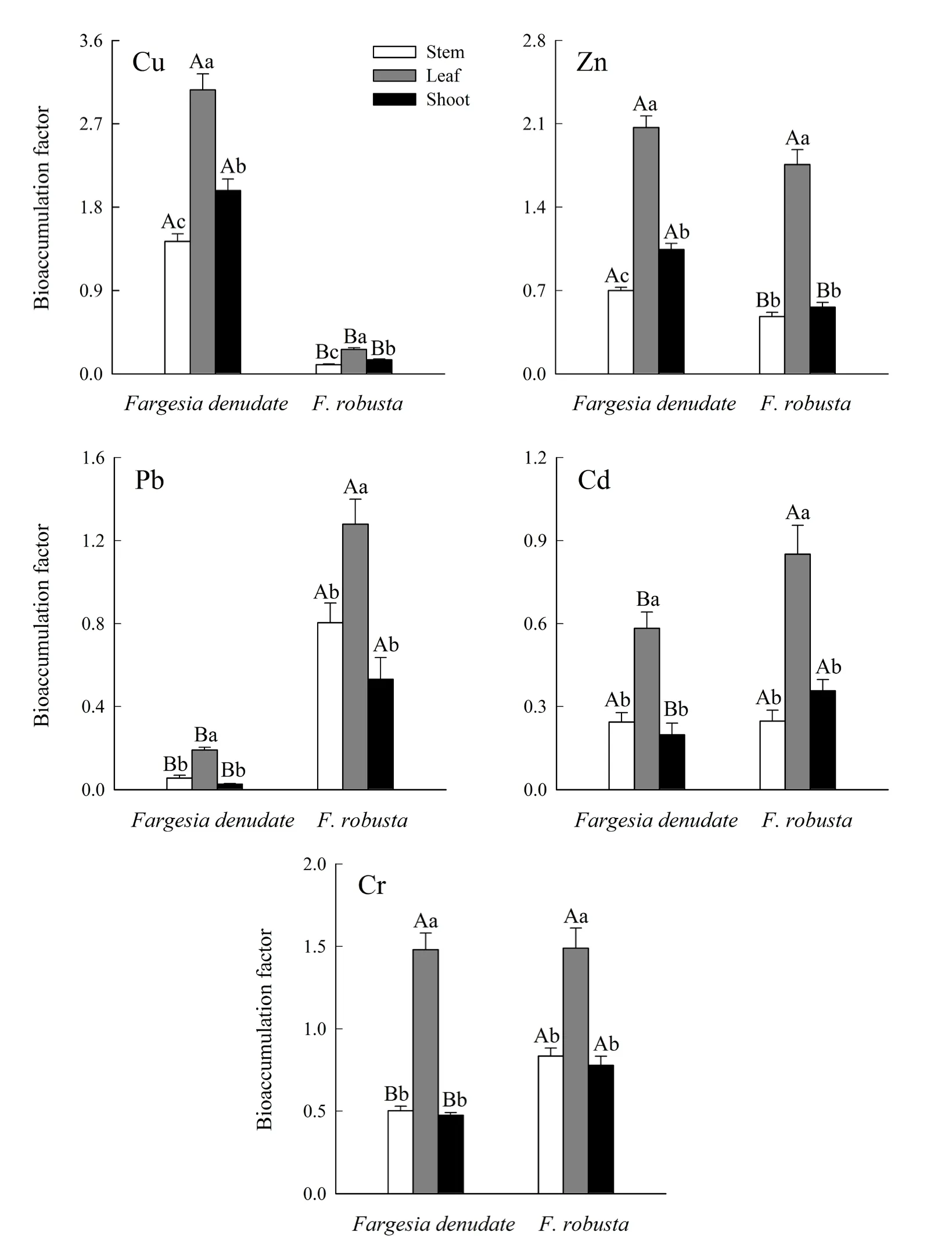
图2 缺苞箭竹和拐棍竹茎、叶、笋的重金属富集系数Fig. 2 The heavy metals bioaccumulation factors in the stems, leaves and shoots of Fargesia denudate and F. robusta
2.4 小径竹的重金属富集系数与超积累植物的比较
通过与生长在未被污染土壤中的超积累植物相比,结果表明,小径竹对多种重金属具有一定的富集潜力。由于取样区的土壤Cd含量高于国家土壤环境质量标准(二级),而缺苞箭竹和拐棍竹在此生长良好,可见其对Cd具有一定的耐受性。与已有研究结果相比,在未被污染土壤中,缺苞箭竹地上部的Cu富集系数高于荸荠、香薷、蓖麻、鸭跖草等常见的超积累植物(表5);拐棍竹地上部的Cu富集系数高于荸荠,而低于香薷、蓖麻、鸭跖草等超积累植物(表 5),说明缺苞箭竹具有较大的Cu富集潜力,而拐棍竹对Cu的富集能力还有待进一步研究。对于重金属 Zn而言,缺苞箭竹的地上部富集系数低于常见的Zn超积累植物东南景天(表5);拐棍竹地上部的Zn富集系数也较其低(表5),说明两种小径竹可能都不具有 Zn富集潜力。对于重金属 Pb而言,缺苞箭竹地上部富集系数低于羊茅、白莲蒿、圆叶无心菜等常见的 Pb超积累植物(表5);而拐棍竹地上部富集系数较常见的Pb超积累植物高(表5),说明拐棍竹对Pb具有较大的富集潜力,而缺苞箭竹可能不具有 Pb富集能力。对于重金属 Cr而言,缺苞箭竹和拐棍竹的地上部富集系数均高于狼尾草、假稻等典型的 Cr超积累植物(表5),说明两种小径竹都具有较大的Cr富集潜力。可见,缺苞箭竹对 Cu、Cr具有较大的富集潜力,而拐棍竹具有较大的Pb、Cr富集潜力。
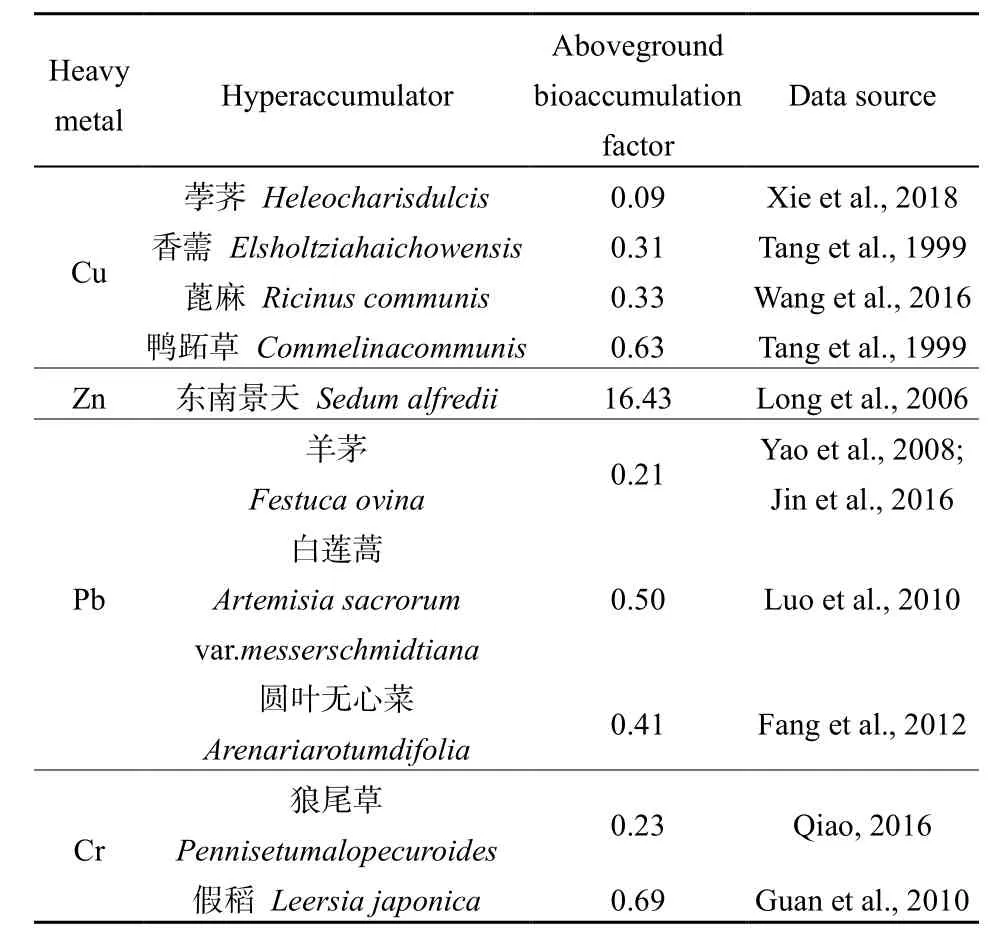
表5 常见超积累植物的地上部重金属富集系数Table 5 The aboveground bioaccumulation factors of common hyperaccumulators
3 讨论
3.1 小径竹不同部位的重金属富集能力
不同植物或同一植物的不同部位在离子累积方面存在较大的差异。一般情况下,植物体内的重金属离子在不同部位的含量大小顺序为根>叶>茎>花>种子(Seregin et al.,2001)。缺苞箭竹和拐棍竹这两种箭竹属小径竹叶的重金属含量和富集系数显著高于茎和笋,这与前人对金镶玉竹、紫竹、黄纹竹、篌竹、雷竹、白荚竹等刚竹属小径竹的研究结果不一致(姜黎,2009;王兵,2011),这可能与地下茎形态有关,缺苞箭竹和拐棍竹均是合轴型丛生竹,而刚竹属竹子的地下茎呈单轴散生,且有研究表明不同地下茎形态竹子各器官的钾含量存在差异,但对其重金属含量和分配格局等方面的差异鲜有报道,有待进一步研究(尹瑞安等,2018;Das et al.,2008)。而这两种禾本科小径竹的重金属含量和富集系数的分配格局与前人对狼尾草、假稻、扁穗牛鞭草、李氏禾等禾本科超积累植物的研究结果相似(Zhang et al.,2009;高洁等,2012;Chandra et al.,2018),说明重金属 Cu、Zn、Pb、Cd、Cr在缺苞箭竹和拐棍竹地上各部位的传输和分布规律与常见的禾本科超积累植物相似,均呈现出叶的重金属富集能力大于茎。
小径竹不同部位对各重金属表现出不同的富集能力。缺苞箭竹对不同重金属富集能力的大小顺序在地上各部位相同,茎、叶、笋的重金属富集系数均表现为 Cu>Zn>Cr>Cd>Pb;而拐棍竹茎、叶、笋对不同重金属富集能力的大小顺序不同,可见,重金属Cu、Zn、Pb、Cd、Cr在缺苞箭竹地上各部位的传输和转移规律相似,而在拐棍竹地上各部位的分布规律有所差异,这与Florijin et al.(1993)研究结果相似,说明不同小径竹各部位的重金属富集能力存在差异。
3.2 不同小径竹的重金属富集能力
不同小径竹的重金属富集能力不同。缺苞箭竹地上部的Cu、Zn富集系数显著大于拐棍竹,而Pb、Cd、Cr富集系数显著小于拐棍竹,这与Broadley et al.(2007)和Collin et al.(2012)研究结果相似,说明小径竹对重金属的富集能力可能更多地受物种基因控制。缺苞箭竹地上部的富集系数呈现出Cu>Zn>Cr>Cd>Pb,拐棍竹地上部的富集系数为Cr>Zn>Pb>Cd>Cu,缺苞箭竹地上部对 Cu 的富集系数大于Zn、Pb、Cd、Cr,而拐棍竹对Cu的富集系数较小,这可能与小径竹体内的SOD、CAT、POD等抗氧化系统酶有关(Scandalios,1993;杨兵等,2004),也可能是受编码钙调蛋白的基因所控制(Jorge et al.,2001),还有待进一步研究。通过与超积累植物比较,发现缺苞箭竹对 Cu、Cr具有较大的富集潜力,而拐棍竹具有较大的 Pb、Cr富集潜力。因此,在使用小径竹进行重金属污染修复时,需根据基质的性质选择适宜的竹种。
竹类植物的地上生物量巨大,对修复重金属污染土壤具有重要价值。根据现有研究数据,中国常见的Cu超积累植物荸荠、海州香薷、鸭跖草的地上生物量分别是0.34、0.41和1.59 kg·m-2,Zn超积累植物东南景天地上生物量是0.18 kg·m-2,Pb超积累植物高羊茅的地上生物量是1.30 kg·m-2,Cd超积累植物宝山堇菜的地上生物量是 0.30 kg·m-2,Cr超积累植物假稻和牛鞭草的地上生物量分别是1.63 kg·m-2和 0.42 kg·m-2(表 6);缺苞箭竹地上生物量(9.75 kg·m-2)和拐棍竹地上生物量(2.79 kg·m-2)均高于中国常见的已报道超积累植物,这可使其富集重金属的总量较大。并且,小径竹能够积累更多的Si元素,这可通过调控基因表达缓解重金属的毒性,提高自身的耐受能力(Li et al.,2008)。另外,西南山地小径竹属于典型的多年生克隆植物,具有生长快、冬季不枯萎、叶片生长周期长、地上现存生物量高等特点,使其在土壤污染修复方面具有种植成本低、易管理、固持量大、归还率低等优势。
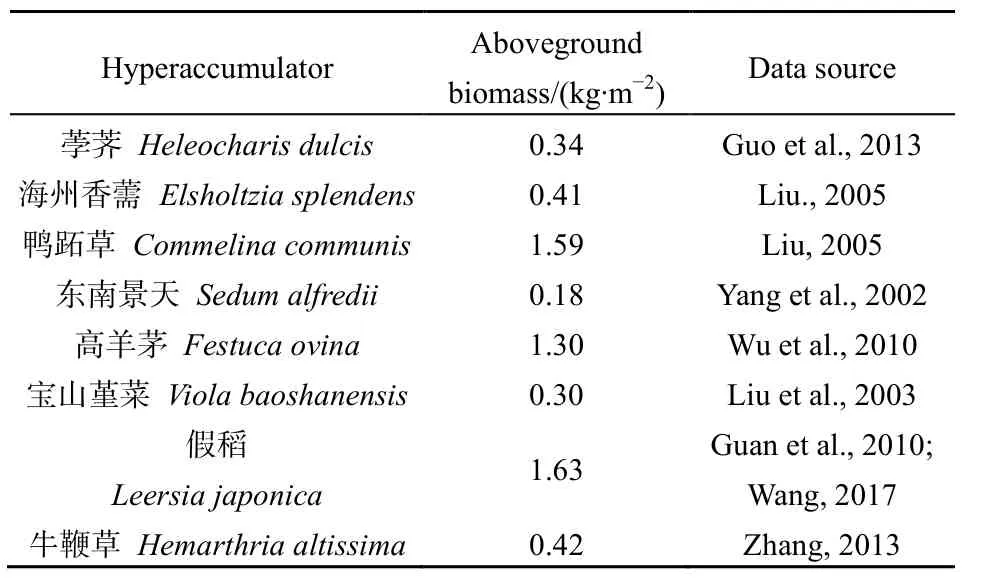
表6 常见超积累植物的地上生物量Table 6 The aboveground biomass of common hyperaccumulators
4 结论
缺苞箭竹和拐棍竹地上部分对重金属Cu、Zn、Pb、Cd、Cr的积累能力不同,叶的重金属含量和富集系数大于茎和笋。与生长在未被污染土壤中的超积累植物相比,缺苞箭竹地上部对Cu、Cr具有较大的富集潜力,而拐棍竹具有较大的Pb、Cr富集潜力。基于西南山地多年生克隆生长的小径竹具有适生范围广、地上现存生物量高、归还率低、经济价值高等特点,可通过控制实验进一步研究小径竹在重金属胁迫下的生理响应及其吸收规律,有望使其发展成为生态-经济型的土壤重金属修复植物。
