黄芪幼苗对镉胁迫的生理响应机制
2019-07-19马月花郭晓瑞唐中华王洪政
马月花 郭晓瑞 杨 楠 张 野 唐中华 王洪政
(东北林业大学森林植物生态学教育部重点实验室,哈尔滨 150040)
镉(Cd)是植物的非必需元素,也是毒性最强的重金属污染物之一。目前,由于采矿、冶炼、垃圾焚烧以及含Cd肥料的过量使用,土壤环境面临着巨大的Cd污染危机[1]。据统计,我国土壤中度和重度镉污染点位超标率为1%[2]。Cd对植物的毒害通常表现为根系和地上部生长受阻、水分和养分吸收受到抑制、生理代谢紊乱,并最终导致植物产量下降甚至引起植物死亡[3~4]。更为重要的是,镉容易积累在植物中进而通过食物链进入人体,对人体健康造成巨大威胁[5]。
在面对Cd胁迫的过程中,植物也进化出了多种应对Cd胁迫的耐性机制。目前,国内外学者一般都将植物对Cd毒害的耐性机制概括为两类:避性机制和耐性机制[6~7]。避性机制主要是指植物通过根系以及根际微生物的分泌物等对周围的Cd2+进行螯合降低其毒性、或通过限制Cd2+的跨膜吸收减少进入植物体内Cd的数量[8~10]。耐性机制则是指植物体内进化出特定的机制来减缓进入植物体内的Cd对植物生长发育造成的危害,如Cd2+的区域化、将Cd2+螯合至无毒状态、提高抗氧化活性避免引起次级伤害等[11~12]。一般来说,一种植物可能会具有不止一种的耐性机制,以提高其对Cd胁迫的抵抗能力。
黄芪(Astragalusmembranaceus(Fisch.)Bunge)是我国有名的传统药用植物,其干燥后的根茎中含有黄酮、皂苷和多糖等多种活性成分,具有补气固表、利尿排脓、敛疮生肌的功效[13]。虽然目前黄芪的主产区还未出现严重的土壤Cd超标情况,但是随着北方土壤中含Cd农药和化肥的过量使用以及工业排放的增加,黄芪主产区的土壤也已处在Cd污染的威胁之下。已有调查结果显示,不同产区黄芪中药材样品中已出现不同程度的Cd超标情况:山西29.41%、内蒙古25%、黑龙江7.65%、甘肃13.16%[14]。因此,研究Cd对黄芪生长发育的影响并探讨黄芪对Cd胁迫的耐性机制具有重要的理论和实践意义。
本研究拟以黄芪幼苗为材料,在水培条件下研究不同程度Cd胁迫对其生长形态、矿质元素吸收转运、次生代谢物质合成等的影响,并研究黄芪对Cd的吸收和积累情况,以期在生理水平上探讨得到黄芪对Cd的耐性机制。
1 材料与方法
1.1 实验材料
膜荚黄芪(A.membranaceus(Fisch.) Bunge)种子购自河北省安国市元泰药用植物种子站。黄芪皂苷Ⅰ-Ⅳ(Astragaloside Ⅰ-Ⅳ)、毛蕊异黄酮(Calycosin)、毛蕊异黄酮苷(Calycosin-7-O-β-D-glucoside)、刺芒柄花素(Formononetin)和刺芒柄花苷(Ononin)标准品均购自上海纯优生物科技有限公司。色谱级甲醇和乙腈购于北京百灵威化学有限公司;K、Ca、Mg、Fe、Cu、Zn、Mn、Cd的标品购于国家有色金属及电子材料分析测试中心;去离子水(电阻率高于18.2 MΩ·cm-1)取自Milli-Q水纯化系统(Millipore,USA)。
1.2 实验方法
1.2.1 黄芪幼苗培养及Cd处理
黄芪种子用10%(v/v)次氯酸钠浸泡2 min消毒并用自来水冲洗3 min后,在蒸馏水中吸胀12 h,然后播种于湿润的蛭石中。待黄芪子叶出土并长至4~5 cm高时,选择长势一致的幼苗移栽至盛有Hoagland营养液(pH5.5)的培养箱中进行培养。所有培养实验均在植物培养室中进行,条件为:温度为30℃/20℃(白天/黑夜),白天时间为14 h,光照强度为400 μmol·m-2·s-1,相对湿度保持在60%~70%。水培3周后对黄芪幼苗进行Cd处理,处理时培养液中加入CdCl2溶液并使其终浓度达到25、50、100、200 μmol·L-1。每个处理设3个重复,每个重复包括10株幼苗。处理时间为7 d,期间每2 d更换处理液。处理结束后进行各指标的测定。
1.2.2 幼苗伸长抑制率及生物量测定
在处理前后用直尺(精度为1 mm)分别测定植株地上部的高度和根系长度,伸长生长抑制率计算如下:
(1)
处理结束后,将幼苗从根茎结合处分开,吸干表面水分后分别称重(鲜重),110℃杀青10 min后60℃烘干至恒重并称重(干重)。
1.2.3 光合色素和丙二醛含量的测定
取黄芪幼苗植株顶端第二叶序的叶片,按照张宪政的方法[15]测定叶绿素a、叶绿素b以及类胡萝卜素的含量。
地上部和根系丙二醛含量采用硫代巴比妥酸(TBA)法进行测定[16]。
1.2.4 植物中金属元素含量的测定
处理结束后,收获黄芪幼苗根系和地上部,并将地上部分为顶部、中部和基部3个部分。用去离子水将各部分冲洗干净后80℃烘干。将烘干样品磨碎后,准确称取0.300 g放入消煮管中,并加入10 mL硝酸—高氯酸混合液(v/v=5∶1)进行消煮。消煮结束后消煮管内加入5 mL 2%(v/v)硝酸并温浴后转移至25 mL容量瓶中,用2%硝酸溶液进行定容。
溶液样品中金属元素的含量使用电感耦合等离子发射光谱仪(ICP-OES Optima 8000)进行测定,其具体含量由各元素标准曲线计算得出。
1.2.5 次生代谢物质含量的测定
取黄芪幼苗根部干样0.50 g,加入80%乙醇10 mL超声提取45 min(100 Hz,40℃),过滤后将残渣再用10 mL溶剂提取一次,合并两次提取液。使用真空离心浓缩仪将提取液浓缩至干,用1 mL甲醇复溶样品,14 000 r·min-1离心10 min,取上清液保存于-20℃冰箱中备用。
样品中黄芪皂苷和类黄酮的含量使用超高效液相色谱—质谱联用仪进行测定,具体色谱和质谱条件等按照杨楠等的方法[17]进行。
1.3 数据处理
文中所列数据均为3次重复的平均值,用SPSS统计软件(19.0,IBM)对实验数据进行统计分析,并采用SigmaPlot软件(14.0,Systat Software)作图。
2 结果与分析
2.1 Cd处理对黄芪幼苗生长的影响
Cd处理显著抑制了黄芪幼苗根系和地上部的伸长生长,且根系对Cd处理的反应更为敏感,因为地上部在50 μmol·L-1Cd处理下才出现明显的生长抑制,而根系在25 μmol·L-1浓度时即表现出明显生长抑制。此外,根系和地上部生长抑制率均随着Cd处理浓度增加显著上升,但相同处理浓度下根系生长抑制率明显高于地上部,经拟合计算得知根系和地上部的IC50分别为62和248 μmol·L-1(图1)。与伸长生长相同,在Cd处理下,特别是较高浓度Cd处理下,黄芪幼苗地上部和根系的鲜重及干重也明显低于对照。
Cd处理引起了黄芪幼苗新生叶片明显变黄,经测定发现第二叶序叶片中叶绿素a、b、总叶绿素及类胡罗卜素的含量在较低浓度Cd下就比对照降低了50%左右(图1)。虽然光合色素的含量随着Cd浓度的升高有继续降低的趋势,但是不同处理浓度间的差异远远小于Cd处理与对照之间的差异。除叶片变黄外,Cd处理还使得黄芪幼苗根系的颜色由白色变为了棕色(图1),说明处理后黄芪根系表皮细胞受到伤害。MDA含量测定的结果显示,50 μmol·L-1Cd处理引起了黄芪幼苗根系MDA含量的明显升高。Cd处理虽然也提高了黄芪幼苗地上部MDA的含量,但是其影响程度远低于根系。
2.2 黄芪幼苗生长对Cd的吸收和区域化
随着Cd处理浓度的升高,地上部和根系的Cd含量逐步升高,并且其含量均与处理浓度之间呈现出良好的正相关关系。在不同浓度Cd处理下,黄芪幼苗根系的Cd含量始终远远高于地上部,最高可达地上部的13倍(图2)。从Cd转移系数(TF)结果来看,低Cd处理下黄芪幼苗Cd的TF值仅为0.055,虽然随着Cd处理浓度的升高TF值也有所升高,但是在200 μmol·L-1Cd处理下TF值也仅为0.157。
除此之外,我们还发现在所有Cd处理中地上部基部的Cd含量始终维持最高,中部Cd含量次之,而顶端Cd含量最低(图2)。当Cd处理浓度为200 μmol·L-1时,地上部基部Cd含量分别为中部和顶端的1.5和2.9倍。
2.3 Cd处理对黄芪幼苗矿质元素吸收和转运的影响
黄芪幼苗根系矿质元素的吸收受到了Cd处理的严重影响(图3)。Cd处理降低了根系中K、Ca、Zn、Mn的含量,特别是K,其含量在200 μmol·L-1Cd处理下比对照下降了46.2%。与上述4种元素的情况相反,根系中Mg、Fe和Cu的含量则受Cd处理的影响而增加,并且其增加程度随着Cd处理浓度的升高而升高。

图1 不同浓度Cd处理对黄芪幼苗生长的影响 所有实验进行了3次重复,误差线代表标准误差。下同。Fig.1 Effect of different Cd treatments on the growth of A.membranaceus seedlings All experiments were performed in triplicate,and error bars represent SE.The same as below.
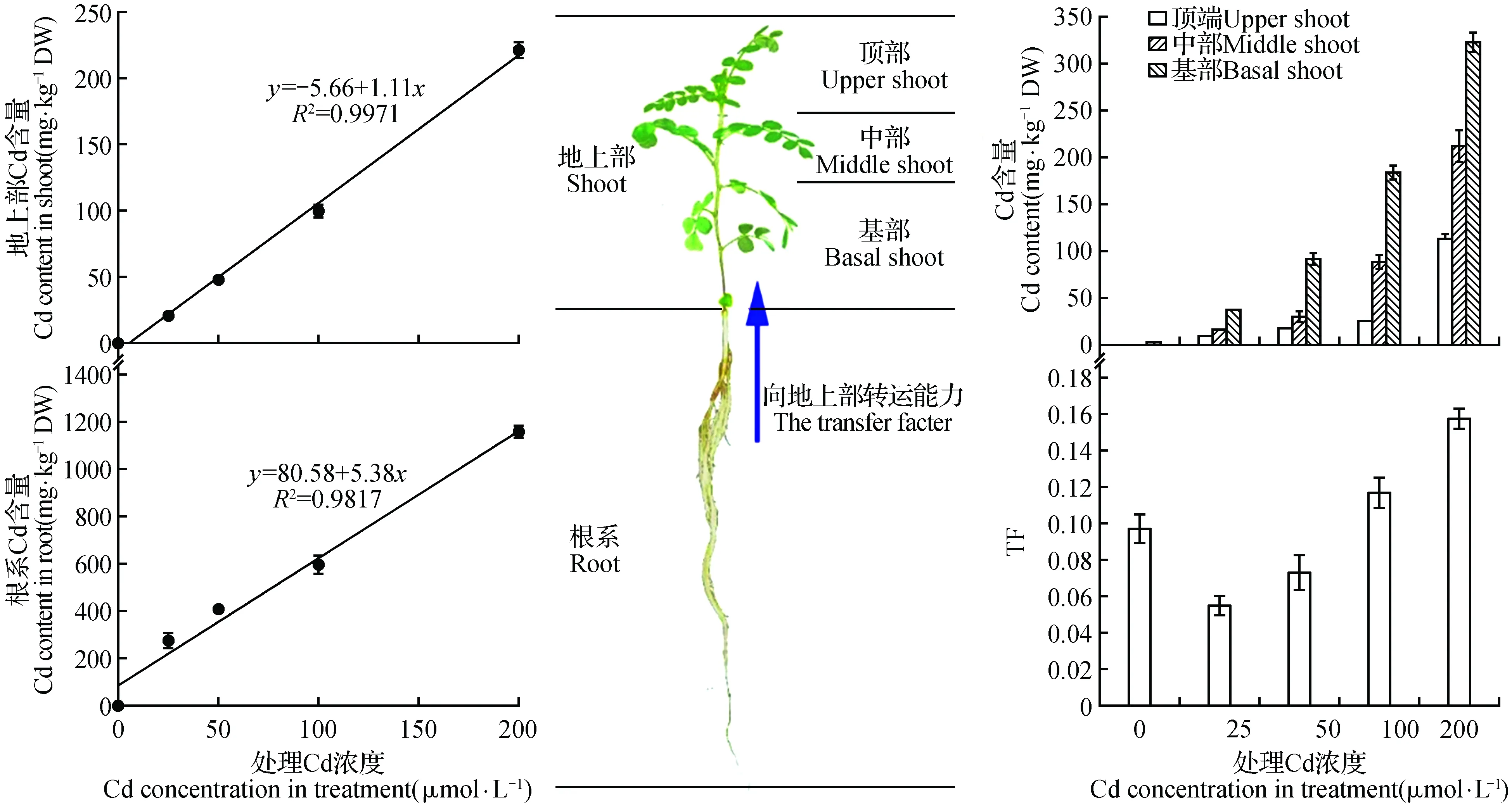
图2 黄芪幼苗对Cd的吸收和转运Fig.2 The adsorption and translocation of Cd in A.membranaceus seedlings
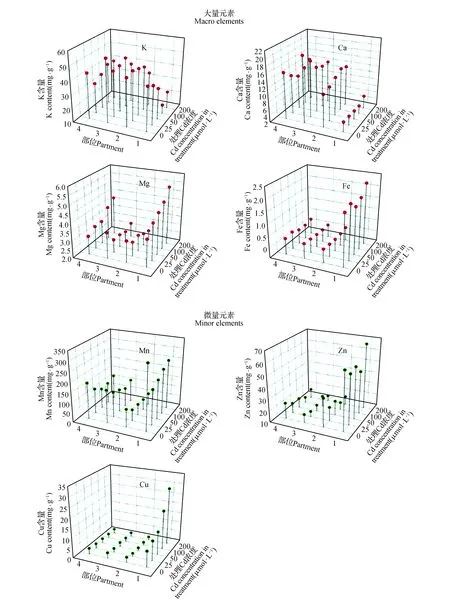
图3 Cd对黄芪幼苗矿质营养元素吸收和转运的影响 部位1~4依次为根系、地上部顶端、地上部中部和地上部基部。Fig.3 The effects of Cd treatments on adsorption and translocation of mineral elements in A.membranaceus seedlingsPartment 1-4 are root,upper shoot,middle shoot and basal shoot.
与根系不同,Cd处理下地上部顶端、中部和基部中几乎所有矿质营养元素的含量都明显减少(图3)。例外的是,随着处理中Cd浓度的升高,地上部顶端的Ca含量以及基部的Mg含量出现了显著的升高趋势。200 μmol·L-1处理下,地上部顶端Ca的含量比对照升高了20%,基部Mg的含量则升高了38.1%。
2.4 Cd处理对黄芪幼苗次生代谢的影响
异黄酮和皂苷是黄芪中最受人关注的两类次生代谢物质,它们在根系中的含量受Cd处理影响发生了明显变化(图4)。与对照相比,较低浓度的Cd处理即引起了毛蕊异黄酮、刺芒柄花素以及毛蕊异黄酮苷和刺芒柄花苷含量的明显升高。在50 μmol·L-1Cd处理下,4种异黄酮物质的含量都达到最高值,其中毛蕊异黄酮达到了对照的7.2倍。随着处理浓度的进一步升高,异黄酮含量出现了较为明显的下降趋势。与异黄酮的反应不同,Cd处理诱导了根系中黄芪皂苷Ⅰ-Ⅳ含量的显著降低,并且黄芪皂苷含量下降的幅度受Cd浓度影响的程度远低于Cd处理本身。
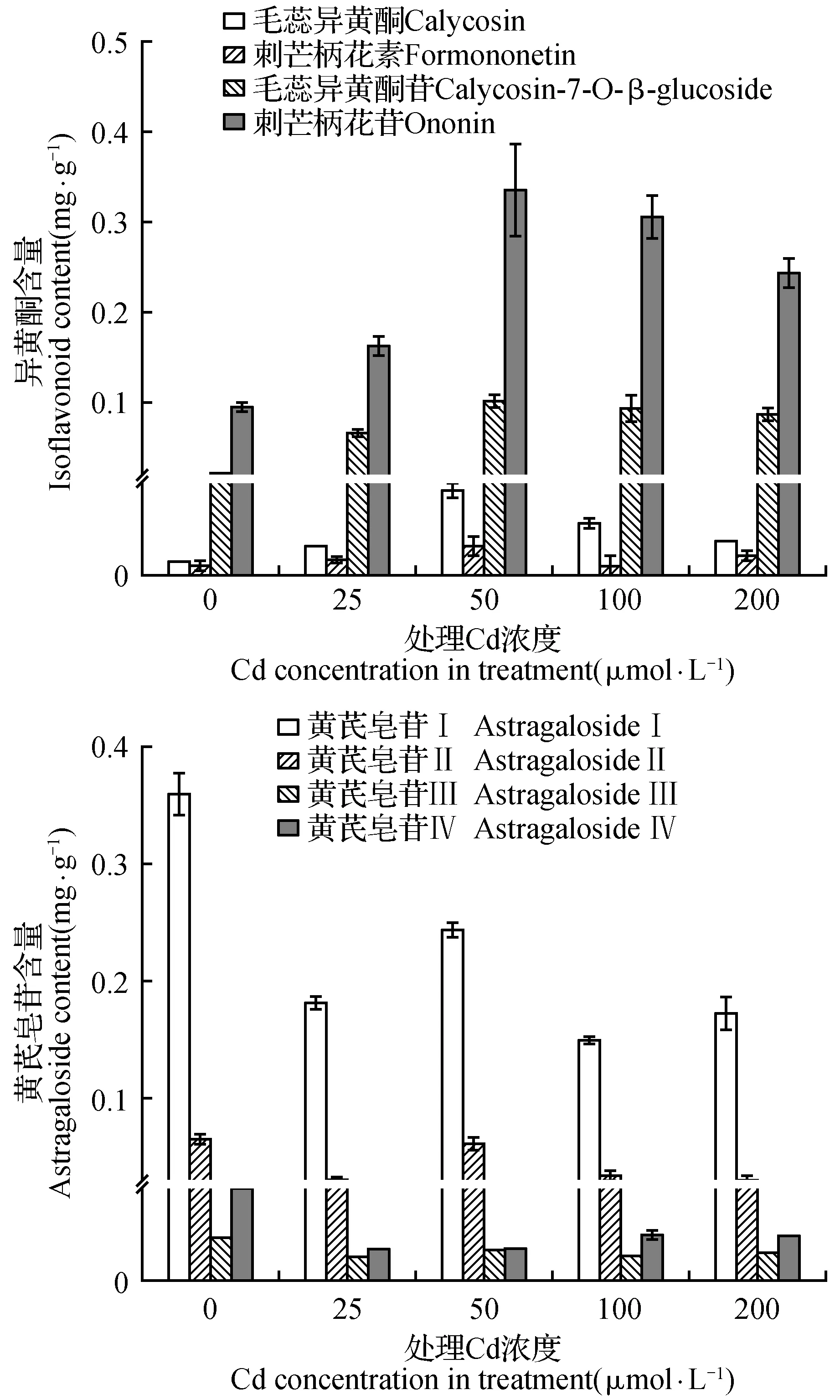
图4 Cd对黄芪幼苗根系次生代谢的影响Fig.4 The effects of Cd treatments on secondary metabolism in roots of A.membranaceus seedlings
3 讨论
作为毒性最大的重金属元素之一,Cd在较低浓度时就可以引起植物的中毒反应,最为基本的可见症状为生长速率的降低以及随之而来的叶片失绿等[18]。在本研究中,25 μmol·L-1Cd处理7 d后,黄芪幼苗根系伸长生长就受到了显著抑制,随着Cd处理浓度的升高,株高以及根系和地上部生物量积累也都明显降低。此外,Cd处理还降低了黄芪幼苗新生叶片光合色素的含量,导致新生叶片失绿变黄。但是,即使在较高浓度Cd处理下,中下部叶片特别是基部老叶未发现失绿现象,暗示Cd处理主要抑制了叶绿素的合成而非促进了原有叶绿素的降解。与本结果相一致,有报道认为Cd引起叶绿素下降的原因可能是影响了叶绿素合成途径的关键酶的活性[19]。同时,我们发现Cd处理下地上部顶端Fe和Mg等与叶绿素合成和活性密切相关的元素含量明显下降,这可能也是引起叶片变黄的因素之一。
Cd诱导的植物膜系统伤害及其透性增加是其重要的毒害机理之一[20]。我们发现,Cd处理引起了黄芪幼苗根系的褐变以及MDA含量的明显增加,显示Cd处理增加了根系质膜的过氧化程度。随着膜脂过氧化程度的加剧,多种位于膜上的重要酶的空间结构也随之遭到破坏,并最终引起膜的空隙和透性增大[21]。干扰植物对矿质营养元素的吸收和转运也是造成Cd毒害的原因之一[22],本研究中Cd处理显著降低了黄芪根系中K、Ca、Zn、Mn以及地上部几乎全部种类元素的含量。Cd毒害引起的根系细胞膜系统透性增加导致小分子物质外流,削弱了植物细胞主动吸收矿质元素的作用[25]是其扰乱营养元素吸收的重要机理之一。其它机理则包括Cd2+与矿质营养元素竞争根系质外体阳离子吸收位点以及转运体蛋白结合位点等[23~24]。
黄芪幼苗在实验浓度Cd处理下显现出的受害现象说明其对Cd胁迫的耐受性远低于十字花科的多种Cd超量积累植物,但是当与其近缘物种苜蓿相比时,它又表现出较高的Cd耐性[26]。限制植物体对Cd的吸收或将进入植物体的Cd区域化地积累在液泡或代谢活性不旺盛的老旧组织中是植物提高其Cd耐性的重要机制[20,27]。在黄芪幼苗中,黄芪吸收的Cd主要积累在根部,向地上部运输的能力很低,其TF值最高仅为0.157。此外,黄芪地上部老叶中Cd的含量最高,而新生组织中含量最低。以上结果说明黄芪通过调控Cd的空间分布,最大限度地保护了地上部新生组织,以提高其在Cd处理下存活的能力。
Ca是植物的必需元素,它与Cd具有强烈的位点竞争效应并在维持膜结构稳定性方面具有重要作用[28]。Cd处理降低了黄芪幼苗根系以及地上部中下部位的Ca含量,却提高了地上部顶端的Ca含量,推测此反应可能在保护黄芪地上部新生组织免受更大的Cd2+伤害方面具有积极作用。此外,在植物中Mg2+和Cd2+之间具有很强的拮抗作用,Mg营养的缺乏会大大提高Cd对植物组织的毒害水平[29]。我们发现,Cd处理下Mg在黄芪地上部的基部发生了特异性的积累上升,这与Cd在地上部的主要积累部位相一致,我们猜测在老叶中高含量的Mg通过与Cd的拮抗作用一定程度上降低了Cd对老叶的毒害作用,最大限度地维持老叶的基本生理功能。
异黄酮和黄芪皂苷类物质是黄芪药用中最主要的两类次生代谢活性物质。研究资料显示,豆科植物细胞在面对不同环境刺激时会选择性地大量积累异黄酮或皂苷类物质,并在其防御反应中起着重要作用[30]。在我们的研究结果中,黄芪面对Cd胁迫时根系中积累了大量的异黄酮类物质,而黄芪皂苷含量则明显下降,表明黄芪选择了增加异黄酮而非皂苷类物质的合成来应对所面临的Cd胁迫。同时,异黄酮类化合物自身还具有有较高的抗氧化活性[31],所以Cd处理下异黄酮的增加还可能在降低Cd引起的氧化胁迫中发挥了积极作用。
综上所述,Cd胁迫可以通过抑制黄芪幼苗根系和地上部生长、阻碍光合色素合成、破坏膜系统稳定性以及干扰矿质营养成分吸收和平衡影响黄芪幼苗的生长发育,而黄芪幼苗则通过将进入体内的Cd进行区域化、调控拮抗性矿质元素定向累积、提高异黄酮物质产量等途径积极响应Cd胁迫,以降低其毒害水平。
