棉花PsbS基因对烟草光合特性的影响
2019-07-18曾晓燕赵瑞海李志博魏亦农
曾晓燕,赵瑞海,李志博,魏亦农
(石河子大学农学院/新疆生产建设兵团绿洲生态农业重点实验室,新疆石河子 832000)
0 引 言
【研究意义】高等植物具有光保护机制,可以减轻光带来的有害影响,非光化学猝灭(NPQ)是光保护机制之一[1],PsbS被认为仅存在于高等植物中,通过与天线蛋白的相互作用来激活NPQ[2],研究PsbS基因在光系统II中的功能对今后选育高光效棉花品种具有现实意义。【前人研究进展】有研究表明,通过表达调节QA氧化还原状态,影响气孔导度和叶片的水分利用效率[3];过量表达PePsbS1和PePsbS2的转基因拟南芥植物均显示出增强的光保护作用,而且PePsbS1和PePsbS2的表达还可以恢复拟南芥npq4突变体的NPQ[4]。已经对该基因在棉花中进行了克隆和功能初步分析[5],该基因能响应多种胁迫及不同的光照处理。【本研究切入点】该基因相关研究主要集中在荧光特性方面,而光合方面研究则相对较少。研究比较烟草中过量表达PsbS株系与野生型的表型差异。【拟解决关键问题】比较烟草中过量表达PsbS株系与野生型的光合相关参数,研究棉花PsbS基因在光保护中作用机制。对PsbS基因在棉花中的具体功能进行了进一步验证。
1 材料与方法
1.1 材 料
试验三个纯系(3个纯合株系L1、L2、L3)已克隆完成的过表达载体pGWB17-GhPsbS,及已转化完成的T2代过量表达PsbS烟草、普通野生型烟草(WT),种植在新疆兵团绿洲生态农业重点实验室人工气候室。
1.2 方 法
1.2.1 试验设计
选择三个烟草株系L1、L2、L3种子均匀点播在1/2 MS培养基中。25~30 d,采用盆栽试验,将幼苗移栽入花盆中,以基质和蛭石按3∶1比例混合装入直径为15 cm、高12 cm 的花盆,每盆移栽一株,转基因烟草(3个纯合株系L1、L2、L3)与野生型烟草各移种9盆,放入人工气候室生长,定期浇适量的水,人工气候室光照为240 μmol/(m2·s),温度为20℃。
生长期间不定期测量各株系的叶面积;当第7片叶完全展开时,取样测叶绿素含量;在光照培养箱进行不同光强处理:75(较低光强)、240 (正常光强)、450 μmol/(m2·s)(较高光强),4 h后采用LI-6400 型便携式光合系统仪(美国Li-cor公司)测量各株系间的光合特性。
1.2.2 转基因植株PCR鉴定
根据改良的CTAB法提取抗性植株的叶片DNA[6],设计PsbS特异引物F:5'-CACCATGGCTCAAACAATGCTGGTAAT-3',R:5'-GTCTTCTTCCGCTTCAT
CAGTAAC-3' (由上海捷瑞生物工程有限公司合成)。利用该对引物,以提取的烟草DNA为模板进行 PCR 扩增。其条件为 94℃预变性5 min;94℃变性30 s,57℃退火30 s,72℃延伸1 min,32个循环;72℃延伸10 min;16℃保存。用1%的琼脂糖凝胶电泳,在凝胶成像系统下检测PCR产物,验证为阳性植株的用于后续实验。
1.2.3 叶面积测定
不同生长时期用长宽法测定烟草叶片叶面积[7],计算叶面积(叶面积/cm2=0.707 5×长×宽),转基因3个株系与野生型各随意选取5株,每株选择最大叶长宽。
1.2.4 叶绿素( Chl) 及类胡萝卜素含量测定
参考张蜀秋等[8]的方法并有所改进,采用直接浸提法,避开叶脉取叶片圆片用80%的丙酮溶液提取(叶片1.1 cm打孔器4片称重+13 mL丙酮溶液),试管用黑布遮住置于黑暗室温条件下放置72 h,期间摇动数次至各绿色器官圆片呈白色时,取其上清液用U-5100 UV/VIS 型分光光度计(日本)于663、645和470 nm波长下比色,用80%丙酮作空白对照。每个株系及野生型烟草取5株的平均值为测定值。计算出提取液中叶绿素a和叶绿素b的含量,叶绿素含量计算公式如下(Lichtenthaler,1983):
叶绿素a的含量(mg/g)= (12.21D663-2.81D646)×V/W.
叶绿素b的含量(mg/g)= (20.13D646-5.03D663)×V/W.
类胡萝卜含量(mg/g)=(1 000D470 - 3.27C(Chla)- 104C(Chlb))/229.
式中D663、D645、D470 分别为相应波长下的光密度值,V为提取液体积,W为所取叶片鲜重。
1.2.5 光合特性测定
人工气候箱温度设为28℃,湿度为37%,75、240、450 μmol/(m2·s)三个光强下转基因株系各3株,野生型3株。仪器使用开放式气路,内置光源,测定转基因各植株净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)等指标。测定时选择植株相同叶片,选取大致相同的部位,最后取各株系的平均值与野生型进行比较。
1.3 数据处理
原始数据采用Microsoft Excel 2010 软件,分别计算平均值和标准误差,并绘制图表。采用统计软件SPSS 19.0 进行显著性分析,图中小写字母和“*”表示转基因与野生型烟草在P< 0.05水平上差异显著。
2 结果与分析
2.1 转基因植株的获得
分别提取野生型和抗性株系的 DNA,以PsbS基因引物进行PCR扩增,在转化株系中都分别扩增出预期目的条带,在野生型烟草植株中未扩增出目的条带,已验证植株为转基因的用于后续实验。图1

注:M:Marker;1~3:转PsbS基因植株;4:WT;5:阴性对照;6:阳性对照
Note: M:Marker; 1-3: transgenic PsbS gene plants; 4: WT; 5: negative control; 6: positive control
图1 转PsbS基因烟草鉴定
Fig. 1 Identification of transgenic PsbS tobacco
2.2 叶面积差异
研究表明,转PsbS基因烟草生长前期的叶片增长幅度明显比野生型烟草叶片增长的快。从移栽入土中开始,19~30 d,转基因3个株系与野生型均差异显著。第19 d时,转基因烟草的平均叶面积是普通野生型的2.36倍,22 d时,转基因烟草的叶面积是普通野生型的2.39倍,而到第30 d时,转基因烟草的叶面积是普通野生型的1.66倍。移栽前期转PsbS基因烟草明显较野生型生长较快,30 d时增长幅度已呈现逐渐下降的趋势。图2
与野生型相比,转PsbS基因的烟草前期叶片明显偏大,而到40 d左右差异逐渐减小。该基因在某种程度上促进了烟草叶片前期的快速生长。图3
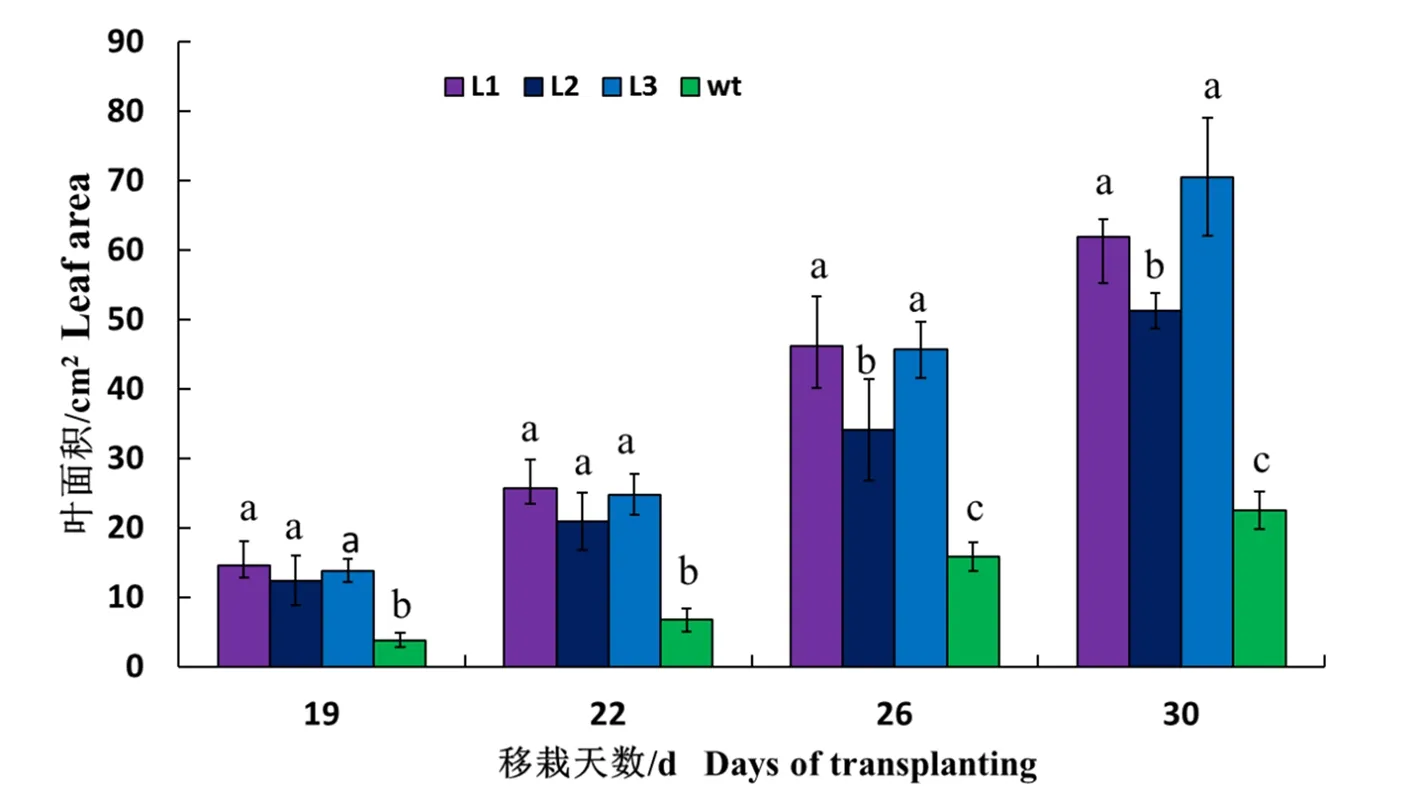
图2 转PsbS基因烟草与野生型烟草不同时期叶面积比较
Fig. 2 Comparison of Leaf Areas between TransgenicPsbSGene and Wild Type Tobacco in Different Periods

图3 转基因烟草与野生型烟草生长状况
Fig. 3 Growth status of transgenic tobacco and wild type tobacco
2.3 转PsbS基因烟草株系叶绿素及类胡萝卜素含量
研究表明,转基因株系L3的叶绿素a、叶绿素b和类胡萝卜素含量与普通野生型差异都极显著,L3的叶绿素a和叶绿素b含量分别是普通野生型的1.39倍和1.62倍,且其类胡萝卜素含量比普通野生型高了37%,L1和L3的叶绿素a、叶绿素b和类胡萝卜素含量与普通野生型虽没有太大差异,但均高于普通野生型。而叶绿素a/b含量有所不同,普通野生型的含量显著高于转基因两个株系。转PsbS基因能够提高烟草的叶绿素a、叶绿素b和类胡萝卜素含量。图4

注:A:叶绿素a; B:叶绿素b; C:类胡萝卜素; D:叶绿素a/b
Note: A: chlorophyll a; B: chlorophyll b; C: carotenoid; D: chlorophyll a/b
图4 转基因烟草株系叶绿素和类胡萝卜素含量
Fig. 4 The strain of transgenic tobacco chlorophyll and Carotenoid content
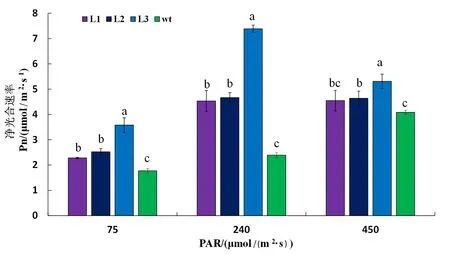
图5 转基因烟草各株系不同光强下的净光合速率
Fig. 5 Net photosynthetic rate of different lines of transgenic tobacco under different light intensities
2.4 转PsbS基因烟草的光合特性
2.4.1 净光合速率(Pn)的影响
研究表明,三个株系(L1、L2、L3)与野生型(普通野生型)间差异显著;在与生长光照相同的240 μmol/(m2·s) 光照处理后,三个株系均与普通野生型存在显著差异,株系L3的光合速率是普通野生型的3倍,而株系L1、L2的净光合速率则是普通野生型的2倍,同时转基因株系间也存在差异;低光强75 μmol/(m2·s)下光合速率虽然都有所下降,但转基因株系的净光合速率还是高于野生型的光合速率,较高光强450 μmol/(m2·s)下L1、L2转基因株系的光合速率比240 μmol/(m2·s)明显有所升高。
2.4.2 气孔导度(Gs)的影响
转基因株系L3在240 、450 μmol/(m2·s)光强下的气孔导度与野生型间差异显著,分别是普通野生型的2.26、1.46倍;虽然株系L1、L2在三个光强处理下与野生型间无差异,但转基因三个株系在不同光强下气孔导度均略高于野生型烟草的气孔导度。图6

图6 转基因烟草各株系在不同光强下的气孔导度
Fig. 6 Stomatal conductance of transgenic tobacco lines under different light intensities
2.4.3 胞间CO2浓度(Ci)的影响
研究表明,不同光强处理后,野生型普通野生型的胞间CO2浓度明显比转基因株系的Ci高,但二者间的胞间CO2浓度无显著差异。当处理光强为75 μmol/(m2·s)时,只有转基因株系L2与普通野生型差异显著,且普通野生型的Ci是株系L2的1.3倍,分别比L1、L3高了15%、18%;而与生长光强一致的240 μmol/(m2·s)和较高光强(450 μmol/(m2·s))处理后,转基因各株系与普通野生型间虽无显著差异,普通野生型的Ci高于转基因株系。图7

图7 转基因烟草各株系在不同光强下的胞间CO2浓度
Fig. 7 Intercellular CO2concentration of transgenic tobacco lines under different light intensities
2.4.4 蒸腾速率(Tr)的影响
研究表明,尽管转基因三个株系的Tr均比野生型的Tr较高,但是只有株系L3的Tr在240、450 μmol/(m2·s)光强处理后与普通野生型存在明显差异,较低光强75 μmol/(m2·s)处理后,三个株系的蒸腾速率整体均比野生型略高,但之间差异不显著;而240 μmol/(m2·s)处理后,株系L3的蒸腾速率Tr是野生型烟草蒸腾速率的2.28倍,株系L1、L2与普通野生型无明显差异;450 μmol/(m2·s)处理后,株系L3的Tr是普通野生型的1.4倍,而株系L1、L2的Tr则分别比普通野生型高了5%、13%。图8

图8 转基因烟草各株系在不同光强下蒸腾速率
Fig. 8 Transpiration rate of each line of transgenic tobacco under different light intensities
2.4.5 水分利用效率(WUE)的影响
研究表明,240 μmol/(m2·s)处理下三个转基因株系与普通野生型相差较大,但只有株系L1与普通野生型存在显著差异;较低光强(75 μmol/(m2·s))下,L3的WUE与普通野生型差异显著;较高光强(450 μmol/(m2·s))处理下,株系L3则出现了水分利用效率比普通野生型 低的情况,且各株系与普通野生型的差异也逐渐缩小,水分利用效率WUE=Pn/Tr ,即单位叶面积上叶片的净光合速率与蒸腾速率之比,不同光强处理下,水分利用效率出现明显差异,正常光强(240 μmol/(m2·s))转基因株系与普通野生型的相对差异较大,而较低光强(75 μmol/(m2·s))和较高光强(450 μmol/(m2·s))下,转基因株系与普通野生型的相对差异较小。图9
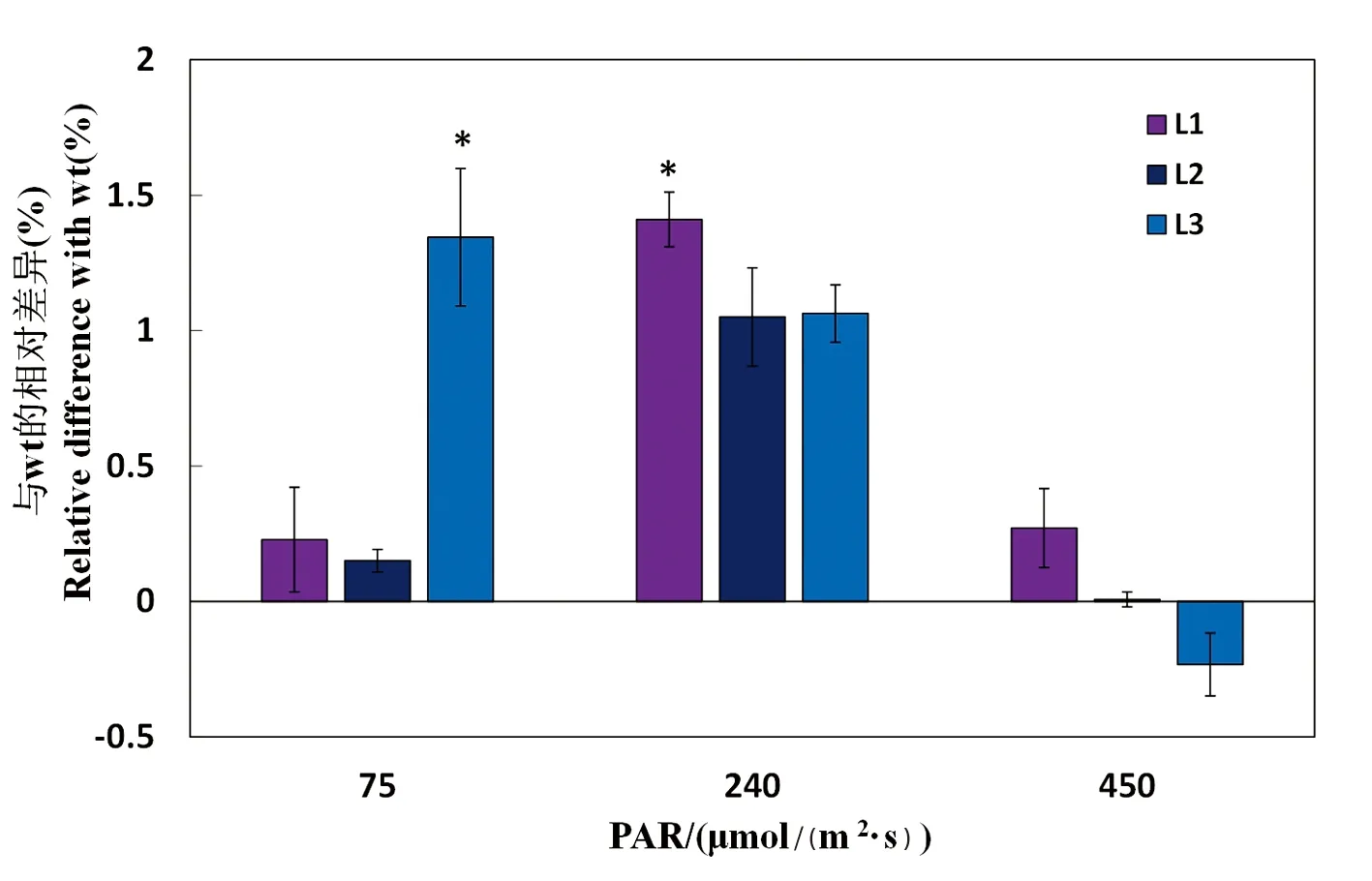
图9 转基因烟草各株系与普通野生型的水分利用效率相对差异
Fig. 9 Relative difference in water use efficiency between transgenic tobacco lines and wild-type
3 讨 论
3.1 转PsbS基因对烟草叶绿素含量的影响
有研究表明[9]植物会通过改变形态特征或生物量分配来提高生存适合度与竞争力,叶片作为烟草光合作用的基础,其较大的生物量有利于烟草捕获光能,维持光合作用的高速运转,从而避免因光能过剩形成的光抑制[10]。研究中,生长前期转基因烟草的叶片比野生型增长较快,与水稻中PsbS缺陷的植株在苗期显示出生长迟缓,并且在生殖阶段的适应性降低[11]等结果相一致,这样使转基因植株更快速的捕获光能,导致了后期对叶片叶绿素含量及光合产生了影响。
反应中吸收利用光能的主要色素-叶绿素与光合速率之间有着密切的关系[12],除少数特殊状态下的叶绿素a分子作为作用中心色素外,其余的叶绿素a和叶绿素b均为聚光色素[13]。在一定范围内,叶绿素含量的增加可以增强叶绿体对光能的吸收与转化,进而增强光合速率。因此,叶片中叶绿素含量的高低是反映植物光合能力的一个重要指标[14]。研究结果发现,转基因株系叶绿素a、b含量与野生型烟草间差异显著,PsbS基因导入烟草后在叶片中得到了表达使转基因烟草株系的叶绿素含量增加,延缓了功能叶片的衰老。
3.2 不同光强对转基因烟草光合的影响
PsbS蛋白是光系统II处结合叶绿素结合蛋白[15,16]。PsbS蛋白之所以被认为在能量耗散中起着关键性的作用[17],主要是强光照导致类囊体膜的酸化[18];正是因为PsbS感受了类囊体膜腔的酸度变化[19,20]进而引起qE以及叶黄素循环来起到光保护的作用。作为能态淬灭产生的关键蛋白之一,它是光系统II超级复合体的蛋白组分之一,更是耗散过剩光能不可或缺的部分,在叶绿体荧光的非光化学淬灭(non-photochemical quenching, NPQ)中发挥着重要作用[17]。光合速率是表示植物在单位面积在单位时间内的 CO2吸收量,或者是 O2释放量,通常情况下植物光合速率值越高,说明植物的生理状态越好[21],实验对转PsbS基因的烟草植株进行了光合速率的测定,发现不同光强下转基因烟草的净光合速率普遍都比野生型烟草的高,结果表明,转基因的烟草促进了叶片的生长,与 Zhang M等[22]的研究结果一致,在东南景天中,超表达SaPsbS基因能够促进烟草的生长,并且能增加烟草的Fv/Fm值,相反的,拟南芥突变体npq4由于缺失PsbS基因而导致植物生长受抑制[23],由于PsbS在高等植物中的作用是普遍的,因此,这种操作应该对所有作物都有效,可能为提高作物产量和品质提供了巨大潜力[3]。
试验中在光强度一定的情况下,随着光照时间的延长,在适宜植株生长的4 h时达到光合最高,这印证了高志民等[24]的毛竹PsbS基因在41℃下诱导4 h的表达效果最好,诱导时间明显影响PePsbS基因的表达,随后降低,趋于平衡。同时通过比较不同光强处理下的转PsbS基因烟草光合性能指标表明,在较低光条件下生长的烟草,净光合速率(Pn)较低,胞间CO2浓度(Ci)较高,气孔导度(Gs)较低,说明烟草叶片对CO2同化不能与光能吸收相协调,在光能捕获不足的情况下最终引起Pn下降;在光合速率较低的情况下通过减少水分的散失从而来适应弱光环境[14],试验中转基因株系在较低光强下(75 μmol/(m2·s))与正常生长条件(240 μmol/(m2·s))的水分利用效率相比,表现出明显的下降趋势,较高光强(450 μmol/(m2·s))与之相比,同样出现明显下降趋势,可以猜想,随着光照强度的增加(>450 μmol/(m2·s)),转基因株系的水分利用效率会逐渐升高,与普通野生型的相对差异会逐渐呈现反向增长状态,具体情况有待进一步研究。而蒸腾速率(Tr)的下降可能与气孔的闭合有关,转基因烟草株系比野生型烟草有较高的气孔导度和蒸腾速率,使其能维持较高的光合速率和积累更多的光合作用产物为结实提供需要。
4 结 论
转GhPsbS基因的烟草苗期叶面积的增长比野生型烟草快,移栽后19、22、26和30 d转基因株系的平均叶面积分别是普通野生型的叶面积的3.57、3.54、2.64、2.72倍,再次验证了该基因能促进植株的生长,且叶绿素a、b含量间也存在显著差异,普通野生型的叶绿素a、b和类胡萝卜素含量分别为0.417、0.1743、0.708,而转基因株系的叶绿素a、b和类胡萝卜素含量分别为0.689、0.223、0.845,较普通野生型分别增长了65%、56%、19%;光合特性方面,转GhPsbS基因的烟草表现出了增强的光合特性,不同光照强度下,转GhPsbS基因株系的净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)均高于野生型烟草。在较高光强450 μmol/(m2·s)条件下,转基因株系的Pn较普通野生型分别高了11.5%、13.7%、30.1%,转基因株系的Tr分别为1.37、1.47、1.82,普通野生型的Tr为1.30,同时转基因株系的Gs分别为0.047、0.054、0.064,比普通野生型分别高了6.8%、22.7%、45.5%。综上说明转GhPsbS基因烟草较野生型具有较强的光合能力。实验的结论与田间试验存在明显差异,验证转入该基因的棉花的具体功能时,选择田间试验更具有可靠性。
