Pretreated Oenanthe Javanica extract increases antiinflammatory cytokines, attenuates gliosis, and protects hippocampal neurons following transient global cerebral ischemia in gerbils
2019-07-17JoonHaParkInHyeKimJiHyeonAhnYooHunNohSungSuKimTaeKyeongLeeJaeChulLeeBichNaShinTaeHeungSimHyunSamLeeJeongHwiChoInKooHwangIlJunKangJongDaiKimMooHoWon
Joon Ha Park , In Hye Kim , Ji Hyeon Ahn YooHun Noh Sung-Su Kim Tae-Kyeong Lee, Jae-Chul Lee, Bich-Na Shin,Tae Heung Sim, Hyun Sam Lee, Jeong Hwi Cho, In Koo Hwang, Il Jun Kang, Jong Dai Kim , Moo-Ho Won,
1 Department of Biomedical Science and Research Institute for Bioscience and Biotechnology, Hallym University, Chuncheon, Gangwon, Republic of Korea
2 Famenity Company, Gwacheon, Geyonggi, Republic of Korea
3 Department of Neurobiology, School of Medicine, Kangwon National University, Chuncheon, Gangwon, Republic of Korea
4 Danchunok Company, Chuncheon, Gangwon, Republic of Korea
5 Department of Histology, College of Veterinary Medicine, Chonbuk National University, Iksan, Jeollabuk-do, Republic of Korea
6 Department of Anatomy and Cell Biology, College of Veterinary Medicine, and Research Institute for Veterinary Science, Seoul National University, Seoul, Republic of Korea
7 Department of Food Science and Nutrition, Hallym University, Chuncheon, Gangwon, Republic of Korea
8 Division of Food Biotechnology, School of Biotechnology, Kangwon National University, Chuncheon, Gangwon, Republic of Korea
Abstract Recently, we have reported that Oenanthe javanica extract (OJE) displays strong neuroprotective effect against ischemic damage after transient global cerebral ischemia. However, neuroprotective mechanisms of OJE have not been fully identified. Thus, this study investigated the neuroprotection of OJE in the hippocampal CA1 area and its anti-inflammatory activity in gerbils subjected to 5 minutes of transient global cerebral ischemia. We treated the animals by intragastrical injection of OJE (100 and 200 mg/kg)once daily for 1 week prior to transient global cerebral ischemia. Neuroprotection of OJE was observed by immunohistochemistry for neuronal nuclear antigen and histofluorescence staining for Fluoro-Jade B. Immunohistochemistry of glial fibrillary acidic protein and ionized calcium-binding adapter molecule 1 was done for astrocytosis and microgliosis, respectively. To investigate the neuroprotective mechanisms of OJE,we performed immunohistochemistry of tumor necrosis factor-alpha and interleukin-2 for pro-inflammatory function and interleukin-4 and interleukin-13 for anti-inflammatory function. When we treated the animals by intragastrical administration of 200 mg/kg of OJE, hippocampal CA1 pyramidal neurons were protected from transient global cerebral ischemia and cerebral ischemia-induced gliosis was inhibited in the ischemic hippocampal CA1 area. We also found that interleukin-4 and -13 immunoreactivities were significantly increased in pyramidal neurons of the ischemic CA1 area after OJE pretreatment, and the increased immunoreactivities were sustained in the CA1 pyramidal neurons after transient global cerebral ischemia. However, OJE pretreatment did not increase interleukin-2 and tumor necrosis factor-alpha immunoreactivities in the CA1 pyramidal neurons. Our findings suggest that pretreatment with OJE can protect neurons and attenuate gliosis from transient global cerebral ischemia via increasing expressions of interleukin-4 and -13. The experimental plan of this study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) in Kangwon National University (approval No. KW-160802-1)on August 10, 2016.
Key Words: Oenanthe javanica extract; transient global cerebral ischemia; hippocampus; ischemic damage;cerebral ischemia; neuroprotection; glial activation; pro-inflammatory cytokines; anti-inflammatory cytokines;inflammation; neural regeneration
Introduction
A temporary disruption of blood circulation in the brain evokes transient global cerebral ischemia (tGCI), and tGCI leads to the death/loss of neurons in regions with high baseline metabolic activity (Kirino, 1982; Globus et al., 1991).Pyramidal neurons in the cornu ammonis (CA) 1 area of the hippocampus are extremely sensitive to tGCI, whereas pyramidal neurons of the CA3 area are more resistant, because CA1 area is characterized by low capillary density compared with the CA3 area (Cavaglia et al., 2001). The selective death/loss of CA1 pyramidal neurons occurs from several days after tGCI (Park et al., 2016; Lee et al., 2017). Many studies have proven the underlying mechanisms of neuronal death following tGCI, including glutamate-mediated excitotoxicity,oxidative stress by excessive reactive oxygen species production and neuroinflammatory response (Niizuma et al., 2009;Ashabi et al., 2015; Park et al., 2017a). Up to recently, based on those mechanisms, many researchers have tried to develop effective neuroprotective agents against cerebral ischemia(Zhang et al., 2017; Zhu et al., 2017).
Diverse plants, which have been traditionally used for medicinal purposes, have gained high attention as attractive candidates to prevent and treat neurological disorders, including cerebral ischemia, because of their diverse biological functions (Dekanski et al., 2011; Raghavan and Shah, 2015).Oenanthe (O.) javanica (water dropwort), which is commonly cultivated in East Asia, is a perennial member of the Oenanthe genus belonging to the Apiaceae family and has been used to heal diverse ailments, such as hypertension, jaundice, fever, urinary difficulties and abdominal pain (Jiang et al., 2015; Ai et al., 2016). Indeed, many researchers have reported via in vitro and in vivo studies that O. javanica extract(OJE) possesses multiple therapeutic properties, including anti-arrhythmic, anti-diabetic and hepatoprotective effects(Ji et al., 1990; Yang et al., 2000, 2014). In particular, we have recently studied that OJE displays strong neuroprotective properties against ischemic damage following tGCI (Park et al., 2015).
Even though, as mentioned above, OJE exerts multiple beneficial effects against various diseases, the neuroprotection of OJE against brain ischemic injury and its mechanisms have been unclear. Therefore, this study was done to investigate the neuroprotective effect of OJE and changes in expressions of pro-inflammatory cytokines (interleukin-2 [IL-2]and tumor necrosis factor-alpha [TNF-α]) and anti-inflammatory cytokines (IL-4 and IL-13) which regulate neuronal survival and death after tGCI (Murakami et al., 2005; Yoo et al., 2016) in gerbils subjected to 5 minutes of tGCI, which have been used as a representative animal model to study the neuroprotective efficacies of diverse agents and their related mechanisms or function (Kaundal et al., 2009; Sasaki et al.,2016).
Materials and Methods
Animals
Male gerbils at 6 months of age (body weight, about 70 g,total n = 42) were purchased from the Experimental Animal Center in Kangwon National University, Chuncheon,Republic of Korea. Animal handling and care followed the guidelines of the current international laws and policies in Guide for the Care and Use of Laboratory Animals (The National Academies Press, 8thEd., 2011). The experimental plan of this study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) in Kangwon National University (approval No. KW-160802-1) on August 10, 2016.
Preparation and treatment of OJE
O. javanica was collected in April 2016 in Kangwon Province (Republic of Korea) by Jong Dai Kim and kept in a deep freezer (-70°C). To prepare OJE, O. javanica was extracted with 70% ethanol (10 vol (v/w)) at 70°C for 4 hours. We repeated the extraction three times, filtered the extract using Whatman Filter Paper (no. 2), concentrated the extract in a vacuum evaporator, and dried the extract in a freeze-drier.We confirmed that the extraction yield was 13.5%.
Experimental animals were randomly assigned to four groups: (1) vehicle-sham group was given 300 μL of saline(0.9% w/v NaCl) and underwent sham operation, (2) vehicle-ischemia group was given saline and subjected to tGCI,(3) OJE-sham group was pretreated with OJE (200 mg/kg)and subjected to sham operation, and (4) OJE-ischemia group was pretreated with OJE and subjected to tGCI. The dose of OJE (200 mg/kg) was selected according to our previous study which showed that pretreatment with 200 mg/kg OJE exhibited a strong neuroprotective potential against tGCI (Park et al., 2015). We dissolved OJE in saline and intragastrically administered it once daily for 1 week prior to tGCI.
Induction of tGCI
tGCI was induced by our published method (Park et al.,2017c). Shortly, gerbils used in this surgery were anesthetized via a mask using a gas mixture of 2.5% isoflurane(Hana Pharm Co., Ltd., Hwaseong, Republic of Korea), 32%oxygen, and 68% nitrous oxide. We occluded both common carotid arteries for 5 minutes, and confirmed the restoration(reperfusion) of arterial blood flow in the retina using an ophthalmoscope. Room temperature (37 ± 0.5°C) was maintained during and after tGCI. Sham gerbils were given the same surgery without the arterial ligation.
Preparation of histological sections
In each group, we used seven gerbils at 2 and 5 days after tGCI. In short, as described previously by Lee et al. (2017),the gerbils were fixed with 4% paraformaldehyde by transcardial perfusion. Their brains were removed and cryoprotected by soaking them in 30% sucrose solution. The frozen brain tissues were serially sectioned into 30-μm thick coronal sections in a cryostat (Leica, Wetzlar, Germany).
Fluoro-Jade B histofluorescence staining
In brief, as described by Lee et al. (2017), we carried out Fluoro-Jade B (F-J B) (a marker for degenerating cells) (Histochem, Jefferson, AR, USA) histofluorescence staining. The brain sections were immersed in 1% sodium hydroxide solution, incubated in 0.06% potassium permanganate solution,and reacted in 0.0004% F-J B solution. Sections stained with F-J B were examined using epifluorescent microscope of Carl Zeiss (Germany).
Immunohistochemical staining
Primary antibodies were mouse anti-neuron-specific soluble nuclear antigen (NeuN) (1:1000, Chemicon, Temecula,CA, USA) to stain neuronal cells, mouse anti-glial fibrillary acidic protein (GFAP) (1:800, Chemicon) to stain astrocytes,rabbit anti-ionized calcium-binding adaptor molecule 1(Iba-1) (1:800, Wako, Tokyo, Japan) to stain microglial cells,rabbit anti-IL-2 (1:200, Santa Cruz Biotechnology, Santa Cruz, CA, USA) and rabbit anti-TNF-α (1:1000, Abcam)to stain pro-inflammatory cytokines, and rabbit anti-IL-4(1:250, Santa Cruz Biotechnology) and rabbit anti-IL-13(1:250, Santa Cruz Biotechnology) to stain anti-inflammatory cytokines. We carried out immunohistochemistry according to a published method by Park et al. (2014). In short, we incubated the sections in the above-mentioned primary antibodies for 12 hours at 4°C and reacted them to biotinylated goat anti-mouse or donkey anti-rabbit IgG (1:200, Vector,Burlingame, CA, USA) for 2 hours at room temperature, and streptavidin peroxidase complex (1:200, Vector, Burlingame,CA, USA) for 1 hour at room temperature. Lastly, each immunoreaction in the reacted tissue was shown using 3,3′-diaminobenzidine (DAB) solution (in 0.1 M Tris HCl buffer).
Data analysis
To analyze the protective effect of OJE against tGCI, neurons stained with NeuN (NeuN+neurons) and cells positive to F-J B (F-J B+cells) in the CA1 area were counted according to a published method by Park et al. (2017c). In short, digital images were captured from five sections per gerbil with a light microscope (BX53, Olympus, Hamburg, Germany) equipped with camera of DP72 (Olympus) connected to a PC. We counted NeuN+neurons and F-J B+cells in a 250 × 250 μm2,which includes the pyramidal cell layer at the middle of the CA1 area, using Optimas 6.5 image analysis software package (CyberMetrics, Scottsdale, AZ, USA).
GFAP, Iba-1, IL-2, TNF-α, IL-4 and IL-13 immunoreactivities were quantitatively analyzed according to a published method by Park et al. (2017b). Briefly, their images were captured in 140 × 140 μm2area at 40× original magnification)and calibrated into an array of 512 × 512 pixels. Each immunoreactivity was measured by a 0-255 gray scale system to show as a ratio of the relative optical density (ROD) for GFAP and Iba-1 and relative immunoreactivity for TNF-α,IL-2, IL-4 and IL-13. ROD was adjusted as % using a program of Adobe Photoshop version 8.0 (Adobe Systems Inc.,San Jose, CA, USA), and relative immunoreactivity was adjusted using a program of a NIH Image 1.59 software (NIH,Bethesda, MD, USA). Ratios of the ROD and relative immunoreactivity were compared with the vehicle-sham gerbils,which was designated as 100 percent.
Statistical analysis
The data obtained in the current study are expressed as the mean ± SEM and were analyzed using GraphPad Prism version 5.0 (GraphPad Software, La Jolla, CA, USA). Statistical differences of the means between the groups were analyzed by two-way analysis of variance (ANOVA) with Bonferroni post-hoc test, which is a less strict correction for multiple comparisons, to elucidate the neuroprotective effect of OJE.P < 0.05 was considered to be statistically significant.
Results
Neuroprotection of tGCI-induced CA1 pyramidal cell death by OJE
Intact NeuN+neurons were distributed in the stratum pyramidale (SP) of the hippocampal CA1 area in the vehicle-sham and OJE-sham gerbils (Figure 1A, B, D, E, and M), and,in the gerbils, F-J B+cells were not detected in the CA1 area(Figure 1C and F).
In the gerbils with ischemia, neurons stained with NeuN were rarely found (9 ± 1.7 per section) in the SP 5 days after tGCI, and cells stained with F-J B were numerous (67 ± 3.9 per section) in the SP 5 days after tGCI (Figure 1G-I, M,and N). However, many NeuN+cells (75 ± 7.2 per section)were found in the SP in the OJE-ischemia gerbils (Figure 1J, K, and M), and, in this group, F-J B+cells in the SP were significantly decreased (3 ± 1.5 per section, P < 0.05) (Figure 1L and N).
Attenuation of tGCI-induced gliosis by OJE Astrocytes
In the vehicle-sham gerbils, typical GFAP+cells as resting form of astrocytes were mainly scattered in strata radiatum and oriens in the CA1 area, and they were small in cell body and had thin and long processes (Figure 2A). GFAP+cells in the vehicle-ischemia gerbils showed reactive form (hypertrophic with bulky cytoplasm and thickened processes) (Figure 2B and C). In these gerbils, the ROD of GFAP+structure was significantly increased from 2 days after tGCI and the ROD was very high (about 190% of the vehicle-sham group, P <0.05) 5 days following tGCI (Figure 2G).
We did not get significant differences in the structure and ROD of GFAP+cells in the OJE-sham gerbils, compared with the gerbils with sham tGCI (Figure 2D and G). The hypertrophy of GFAP+cells in the OJE-ischemia gerbils was distinctively decreased (Figure 2E and F), and their ROD was significantly lower (139% of the vehicle-sham gerbils, P <0.05) at 5 days after tGCI, compared to the gerbils with tGCI(Figure 2G).
Microglia
Intact Iba-1+cells, as microglia, had a round cell body and thin processes in the gerbils with sham tGCI, and the Iba-1+cells were scattered throughout the hippocampal CA1 area(Figure 2H). In the gerbils with tGCI, the morphology of Iba-1+microglia was strikingly changed with time after tGCI(Figure 2I and J). Concretely, Iba-1+microglia 2 days after tGCI showed hypertrophied bodies with thickened processes as active form (Figure 2I). Five days following tGCI, Iba-1+ cells were concentrated near or in the SP, and they were more activated (Figure 2J). At this time point, significantly increased ROD (about 238% of the vehicle-sham gerbils, P <0.05) was observed in Iba-1+cells (Figure 2N).
In the OJE-sham gerbils, Iba-1+cells were similar in their structure and ROD to those in the gerbils with sham tGCI(Figure 2K and N). tGCI-induced activation of Iba-1+cells in the OJE-ischemia gerbils was distinctly decreased (Figure 2L and M), and their ROD was about 136% of the gerbils with sham tGCI at 5 days following tGCI, compared with the vehicle-ischemia gerbils (Figure 2N).
Immunoreactivities of IL-2 and TNF-α IL-2
Immunoreactivity of IL-2 in the gerbils with sham tGCI was observed in the SP of the hippocampal CA1 area (Figure 3A). IL-2 immunoreactivity in the SP began to significantly decrease (46% of the gerbils with sham tGCI) at 2 days after tGCI, and its immunoreactivity was significantly decreased(28% of the gerbils with sham tGCI) at 5 days following tGCI(Figure 3B, C, and G).
Immunoreactivity of IL-2 in the SP of the sham gerbils treated with OJE was not different from that in the gerbils with sham tGCI (Figure 3D and G). Immunoreactivity of IL-2 in the SP of the ischemic gerbils treated with OJE was not altered until 5 days following tGCI (Figure 3E, F, and G).
TNF-α
The gerbils with sham tGCI showed TNF-α immunoreactivity in the SP (Figure 3H). In the gerbils with tGCI, TNF-α immunoreactivity was not altered at 2 days following tGCI,however, 5 days after tGCI, TNF-α immunoreactivity was significantly reduced (19% of the vehicle-sham gerbils) (Figure 3I, J, and N).
Immunoreactivity of TNF-α in the SP of the OJE-sham gerbils was similar to that in the gerbils with sham tGCI(Figure 3K and N). Immunoreactivity of TNF-α in the ischemic gerbils treated with OJE was not significantly altered until 5 days after tGCI (Figure 3L, M, and N).
Immunoreactivities of IL-4 and IL-13 IL-4
Moderate IL-4 immunoreactivity was observed in the SP of the gerbils with sham tGCI (Figure 4A). A significant decrease (45% of the gerbils with sham tGCI) in IL-4 immunoreactivity was observed at 2 days following tGCI, and its immunoreactivity was very low (36% of the gerbils with sham tGCI) at 5 days following tGCI (Figure 4B, C, and G).
Immunoreactivity of IL-4 was significantly enhanced(139% of the gerbils with sham tGCI) in the sham gerbils treated with OJE (Figure 4D and G). Increased immunoreactivity of IL-4 was found in the ischemic gerbils treated with OJE, and the increased immunoreactivity was sustained until 5 days following tGCI (Figure 4E, F, and G).
IL-13
IL-13 immunoreactivity in the gerbils with sham tGCI was weakly observed in the SP (Figure 4H). IL-13 immunoreactivity in the vehicle-ischemia gerbils was reduced in a time-dependent manner following tGCI, and IL-13 immunoreactivity was very low (33% of the gerbils with sham tGCI) at 5 days following tGCI (Figure 4I, J, and N).
In the OJE-sham gerbils, a significant increase (150% of the vehicle-sham gerbils) in IL-13 immunoreactivity was found (Figure 4K and N). In the OJE-ischemia gerbils, the increased immunoreactivity of IL-13 was not changed until 5 days following tGCI (Figure 4L, M, and N).
Discussion
With accumulating evidences regarding therapeutic effects of medical plants on neurological disorders, some studies have shown the neuroprotective potential of components of O. javanica. For example, Ma et al. (2010) reported that persicarin isolated from O. javanica protected cortical neurons from neurotoxicity induced by glutamate in an in vitro system. Additionally, it was demonstrated that hyperoxide and isorhamnetin, known as components of O. javanica,showed strong neuroprotective properties in many models of ischemia (Liu et al., 2012; Zhao et al., 2016). Furthermore,we recently demonstrated that OJE protected pyramidal cells in the hippocampal CA1 region from tGCI-induced damage in gerbils using cresyl violet staining (Park et al., 2015). To investigate the neuroprotective effects of OJE in more detail against ischemic damage induced by tGCI, we used immunohistochemistry for NeuN and histofluorescence staining for F-J B in this research, because NeuN and F-J B are known as good markers for examining neuronal death/loss in tissues of the central nervous system (Wang and Tsirka, 2005;Huttner et al., 2014). We found that pretreatment with OJE effectively protected CA1 pyramidal cells at 5 days following tGCI. Thus, our present results indicate that OJE exhibits a powerful neuroprotective property against tGCI.
Neuroinflammation is a major in the progression of neuronal death following brain ischemic insults (Ceulemans et al., 2010; Kim et al., 2016). It is well accepted that brain ischemic insults result in activations of resident astrocytes and microglia in ischemic brain areas and the activated cells attend inflammatory response through releases of a variety of neurotoxic molecules including pro-inflammatory cytokines,which would mediate neuronal death/loss (Uno et al., 1997;Amantea et al., 2014). Thus, the inhibition of resident astrocytes and microglia activations following cerebral ischemia has been thought as a part of neuroprotective factors against brain ischemic insults (Kuang et al., 2014; Chen et al., 2015).In that regard, in this research, we show that tGCI-induced activations of glial cells (microglia and astrocytes) in the gerbil hippocampal CA1 area were significantly inhibited by pretreatment with OJE. Based on our present results together with the findings of above-mentioned studies, we suggest that pretreatment with OJE can suppress glial activation in the ischemic CA1 area and the suppression might be associated with the protective potential of OJE against tGCI.

Figure 1 Cells stained with NeuN and F-J B in the CA1 area of the gerbils.(A-L) Representative microscopic images of cells stained with NeuN and F-J B in the CA1 area of the gerbils with vehicle-sham-tGCI (A-C), OJE-sham-tGCI (D-F), vehicle-tGCI (G-I) and OJE-tGCI (J-L) gerbils 5 days following tGCI. In all sham gerbils, cells positive to NeuN are easily observed in the pyramidal layer (SP), and cells positive to F-J B are not found. Rare NeuN+ cells are shown, but many F-J B+ cells are found in the SP (asterisks,green) in the gerbils with tGCI. However, many NeuN+ and few F-J B+ cells are observed in the SP of the ischemic gerbils after OJE treatment. Scale bars: 400 μm in A, D, G, J and 40 μm in B, C, E, F, H, I, K, L. Graphs of numbers of NeuN+ neurons (M) and F-J B+ cells (N) in the CA1 area (n = 7 in each group). *P < 0.05, vs. vehicle-sham gerbils; #P < 0.05, vs. vehicle-ischemia gerbils. Bars indicate the means ± SEM. DG: Dentate gyrus; SO: oriens layer; SR: radiatum layer; NeuN: neuronal nuclei; F-J B: Fluoro-Jade B; tGCI: transient global cerebral ischemia; OJE: Oenanthe javanica extract.
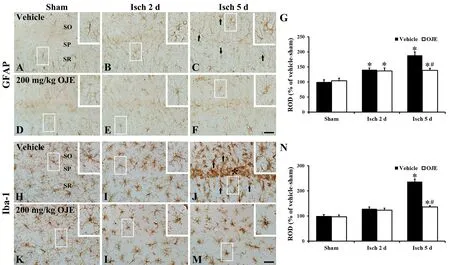
Figure 2 Cells stained with GFAP and Iba-1 in the CA1 area of the gerbils.Representative microscopic images of cells stained with GFAP (A-F) and Iba-1 (H-M) in the CA1 area of the gerbils with vehicle-sham-tGCI (A,H), vehicle-tGCI (B, C, I, J), OJE-sham-tGCI (D, K) and OJE-tGCI (E, F, L, M) at two (B, E, I, L) and five days (C, F, J, M) following tGCI. Marked differences are not found in morphology of GFAP+ and Iba-1+ cells between the vehicle- and OJE-sham-gerbils. In the gerbils with tGCI, glial cells are activated (arrows), especially, many Iba-1+ cells are concentrated and activated in the pyramidal layer (SP, asterisk) at five days following tGCI.In the gerbils with tGCI, the enhanced activation of GFAP+ and Iba-1+ cells is shown. High magnification images are shown in white boxes. Scale bars: 40 μm. ROD as percent of GFAP+ (G) and Iba-1+ structures (N) (n = 7 in each group). *P < 0.05, vs. vehicle-sham gerbils; #P < 0.05, vs. vehicle-ischemia gerbils. Bars indicate the means ± SEM. SO: Oriens layer; SR: radiatum layer; GFAP: glial fibrillary acidic protein; Iba-1: ionized calcium-binding adapter molecule 1; tGCI: transient global cerebral ischemia; OJE: Oenanthe javanica extract; ROD: relative optical density.
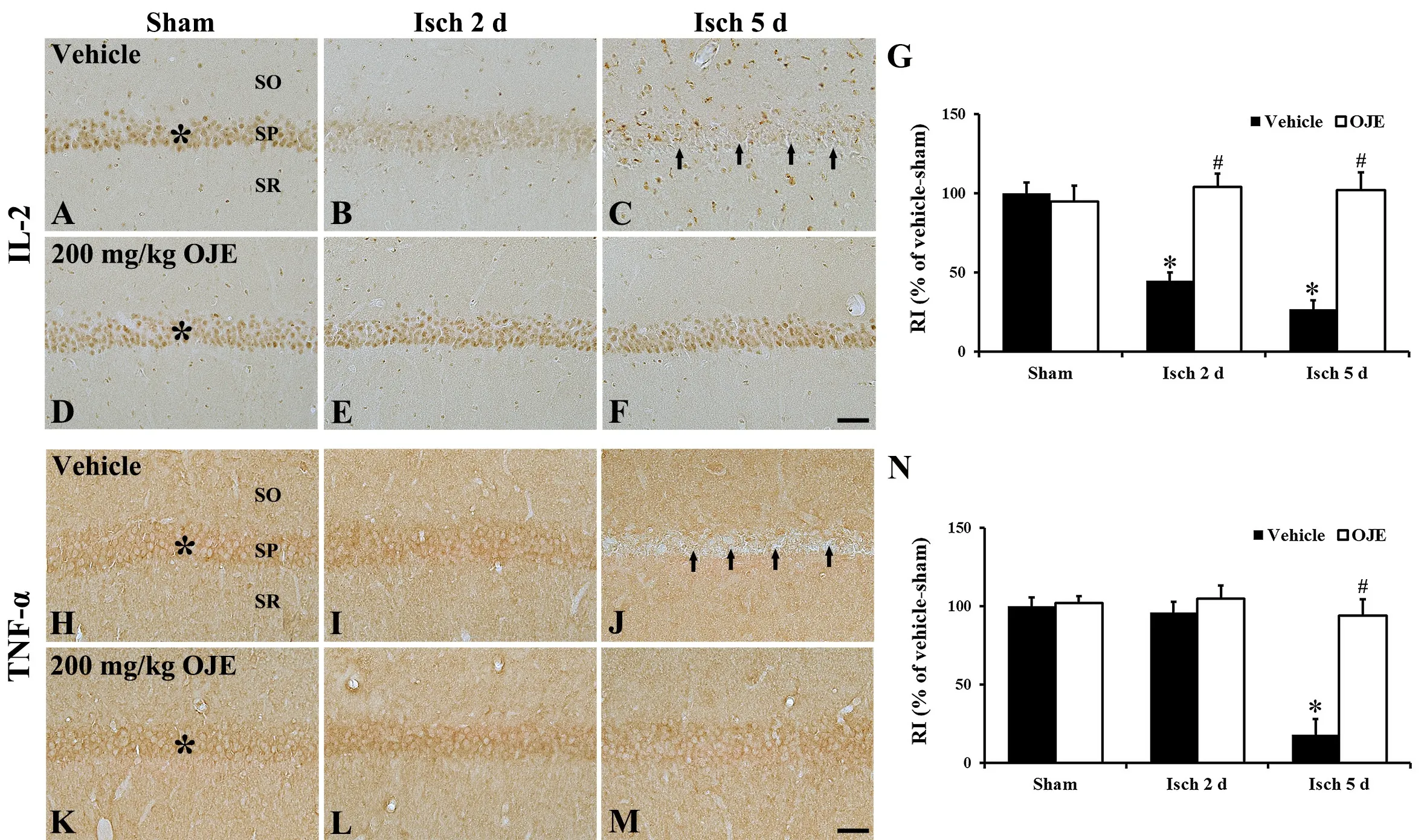
Figure 3 IL-2 and TNF-α immunoreactivities in the CA1 area of the gerbils.Representative microscopic images of IL-2 (A-F) and TNF-α (H-M) immunoreactivities in the CA1 area of the gerbils with vehicle-sham-tGCI (A,H), vehicle-tGCI (B, C, I, J), OJE-sham-tGCI (D, K) and OJE-tGCI (E, F, L, M) at two (B, E, I, L) and 5 days (C, F, J, M) following tGCI. IL-2 and TNF-α immunoreactivities in the pyramidal layer (SP, asterisks) are similar in all sham gerbils. In the gerbils with tGCI, IL-2 and TNF-α immunoreactivities in the SP (arrows) are distinctively reduced at 5 days after tGCI. In the ischemic gerbils treated with OJE, immunoreactivities of IL-2 and TNF-α are not altered compared to the sham gerbils treated with OJE. Scale bars: 40 μm. RI as % of IL-2 (G) and TNF-α (N) immunoreactivities in the CA1 pyramidal cells (n = 7 in each group). *P < 0.05, vs. vehicle-sham gerbils; #P < 0.05, vs. vehicle-ischemia gerbils. Bars indicate the mean ± SEM. SO: Oriens layer; SR: radiatum layer; IL-2: interleukin-2; TNF-α: tumor necrosis factor-alpha; tGCI: transient global cerebral ischemia; OJE: Oenanthe javanica extract; RI: relative immunoreactivity.

Figure 4 IL-4 and IL-13 immunoreactivities in the CA1 area of the gerbils.Representative microscopic images of IL-4 (A-F) and IL-13 (H-M) immunoreactivities in the CA1 area of the gerbils with vehicle-sham-tGCI(A, H), vehicle-tGCI (B, C, I, J), OJE-sham-tGCI (D, K) and OJE-tGCI (E, F, L, M) at 2 (B, E, I, L) and 5 days (C, F, J, M) following tGCI. Immunoreactivities of IL-4 and IL-13 are weakly shown in the pyramidal layer (SP) in the sham gerbils, and these immunoreactivities are gradually reduced with time following tGCI. In the OJE-sham gerbils, immunoreactivities of IL-4 and IL-13 are significantly enhanced in the SP (asterisks) compared to the gerbils with vehicle-sham-tGCI, and these immunoreactivities are maintained in the SP (asterisks) following tGCI. Scale bars: 40 μm. RI as % of IL-4 (G) and 13 (N) immunoreactivities in the CA1 pyramidal cells (n = 7 in each group). *P < 0.05, vs. vehicle-sham gerbils; #P < 0.05, vs. vehicle-sham- or ischemia gerbils. Bars indicate the mean ± SEM. SO: Oriens layer; SR: radiatum layer; IL-4: interleukin-4;IL-13: interleukin-13; tGCI: transient global cerebral ischemia; Isch: ischemia; OJE: Oenanthe javanica extract; RI: relative immunoreactivity.
Cytokines, which encompass diverse groups of inflammatory mediators, are known to act as signaling molecules to coordinate inflammation, which exert positive or negative effects on neuronal survival (Becher et al., 2017). In general,we accept that pro-inflammatory cytokines are involved in the amplification of neuroinflammatory response and contribute to pathogenesis of neurological diseases (Vila et al.,2000; Somera-Molina et al., 2009), while anti-inflammatory cytokines are critically implicated in resolving neuroinflammation via down-regulation of the production of pro-inflammatory cytokines (Ledeboer et al., 2000; Yang et al.,2002). In this regard, we found in this study that immunoreactivities of IL-2 and TNF-α were significantly changed in CA1 pyramidal cells of the gerbils with tGCI, compared with the gerbils with sham tGCI. However, their immunoreactivities were not altered in the OJE-sham-gerbils and OJE-ischemia-gerbils. It has been studied that exogenous TNF-α results in exacerbation of ischemic injury in rat models of transient and permanent focal brain ischemia (Barone et al.,1997) and that dysregulated expressions of IL-2 and TNF-α in ischemic brain areas following transient global and/or focal brain ischemia are involved in developing ischemic neuronal death/loss (Yin et al., 2003; Hwang et al., 2006). Thus,our current findings indicate that pretreatment with OJE would prevent abnormal expressions of IL-2 and TNF-α in CA1 pyramidal cells following tGCI, which might contribute to neuroprotective effects of OJE.
On the other hand, in this research, significant decreases in immunoreactivities of IL-4 an IL-13 were observed in CA1 pyramidal cells following tGCI. However, unlike the immunoreactivities of TNF-α and IL-2, IL-4 and IL-13 immunoreactivities in the sham gerbils treated with OJE were significantly enhanced, compared to those in the sham gerbils, and the increased IL-4 and IL-13 immunoreactivities were consistently sustained in the OJE-ischemia gerbils.Xiong et al. (2011) reported that loss of IL-4 signaling in knock out mice exacerbates subsequent ischemic damage following transient focal cerebral ischemia with increases of macrophage/microglia and leukocyte infiltration. In addition, some researchers demonstrated that upregulations of IL-4 and IL-13 following treatment with some natural materials contributed to neuroprotection in a gerbil model of tGCI (Park et al., 2014; Yoo et al., 2016). Based on these reports, our current findings suggest that increased and sustained IL-4 and IL-13 expressions in pyramidal neurons following pretreatment with OJE would participate in the neuroprotection of OJE against tGCI.
In brief, taken together our overall results, pretreatment with OJE inhibited pyramidal neuronal death and attenuated activations of glial cells in the CA1 area following tGCI. In addition, pretreatment with OJE elevated IL-4 and IL-13 expressions in CA1 pyramidal cells following tGCI. Therefore,OJE can display neuroprotective property of tGCI via reducing glial activation, and enhancing and/or sustaining expressions of anti-inflammatory cytokines following pretreatment with OJE, although the limitations of this study include the lack of data on molecular experiments, such as expressions of genes, which should be addressed in future studies. In this regard, we insist that O. javanica can be utilized as a powerful candidate to prevent tGCI-induced neuronal loss/death in vulnerable brain regions to ischemic insults.
Author contributions:All authors participated in the design, implementation, and evaluation of the study, and approved the final version of this paper for publication.
Conflicts of interest:The authors declare no conflicts of interest relating to the material presented in this article. Its contents, including any opinions and/or conclusions expressed, are solely those of the authors.
Financial support:This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D-1A1B03033271, to JDK), and by the Bio-Synergy Research Project (NRF-2018M3A9C4076478, to IJK) of the Ministry of Science, ICT and Future Planning through the National Research Foundation.
Institutional review board statement:Animal handling and care followed the guidelines of the current international laws and policies in Guide for the Care and Use of Laboratory Animals (The National Academies Press, 8th Ed., 2011). The experimental plan of this study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) in Kangwon National University (approval No. KW-160802-1) on August 10, 2016.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement: This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers:Eduardo Puelles, Institute of Neurosciences,UMH-CSIC, Spain; Nobuyuki Ishibashi, Children's National Health System, USA; Murat Sahin, University of Amasya, Faculty of Medicine,Turkey.
Additional file:Open peer review reports 1 and 2.
杂志排行

中国神经再生研究(英文版)的其它文章
- Novel miRNA, miR-sc14, promotes Schwann cell proliferation and migration
- Role of behavioral training in reducing functional impairments after stroke
- Remodeling dendritic spines for treatment of traumatic brain injury
- Acute drivers of neuroinflammation in traumatic brain injury
- More than anti-malarial agents: therapeutic potential of artemisinins in neurodegeneration
- Why microglia kill neurons after neural disorders?The friendly fire hypothesis