外来入侵植物刺苍耳的繁殖生物学特性研究
2019-07-11顾威马淼
顾威,马淼
(石河子大学生命科学学院/新疆植物药资源利用教育部重点实验室,新疆 石河子 832003)
刺苍耳(XanthiumspinosumLinn.)为菊科苍耳属一年生草本植物,起源于南美洲,现广泛分布于欧洲和东亚地区[1]。植株具有微毒,雌雄同株,茎直立,高30~100 cm,浓密分枝,大量的三叉型尖锐黄色硬刺生长于叶柄基部两侧,刺入肉中会产生较长时间的强烈疼痛[2],因此牛羊均不采食。刺苍耳结实量大,且总苞表面密被倒钩刺,极易通过附着在人类和动物体表,或夹杂在干草和货物当中实现快速广泛的散播,从而迅速占据大面积的区域,抑制农作物、饲草和其他本地植物的生长和繁殖,因而被世界很多国家列为恶性入侵杂草[3]。刺苍耳的野生归化种群于1932年在我国的河南省周口市郸城县被首次发现,现已广泛分布于我国华北和西北的广大地区。截至目前,尽管国内外学者在刺苍耳的生态适应性和竞争能力等方面开展了部分研究,但有关其繁殖生态学特性的研究尚未见报道。
外来种在新栖息地种群建立的成功与否,其繁殖特性起到了决定性的作用[4]。研究[5,6]表明外来物种往往具有很强的繁殖能力,它们能够凭借迅速产生大量种子的策略,在抵达新栖息地后快速建立种群。因此,研究刺苍耳的繁殖生物学特性是揭示其成功入侵机理的有效途径。本文从开花物候、传粉媒介、花粉散播距离、花粉活力、柱头可授性、柱头表面积与花粉粒横截面积的比值、繁殖分配及交配系统等方面,系统地对刺苍耳的繁殖生物学特性进行了研究,为揭示该植物成功入侵的机理提供理论依据。
1 材料与方法
试验地概况
试验地点位于新疆玛纳斯县境内(476 m a.s.l.; 44.15°N,86.17°E)。试验区气候为北温带大陆性干旱气候,冬季寒冷,夏季炎热,日照时间长,蒸发量大,降水稀少。年均气温 7~ 8 ℃,年日照时间2318~2732 h,年降水量180~270 mm,年蒸发量1000~1500 mm[7]。刺苍耳种群为该群落的优势种,种群密度大。对野生刺苍耳种群进行定点观测记录,并开展与刺苍耳繁殖特性相关的实验研究。
1.2 方法
1.2.1 开花物候的观察记录
在刺苍耳种群中随机选取人为影响小、长势大小基本一致的30棵植株,进行标记,2017年6—10月,每隔5 d统计记录每株植株的开花雌花序和雄花序的数量,观察记录其开花物候。参照马文宝等[8]的研究方法,确定个体和群体水平上的始花日期、开花峰值期以及终花期。
1.2.2 传粉媒介的研究
1.2.2.1 访花昆虫的观察
选择晴朗无风天气,08∶00开始,随机选取处于盛花期的刺苍耳10株,观察至20∶00结束。连续30 d定点观察,确定是否有昆虫访花。如果有昆虫访花,则鉴别访花昆虫的种类,记录访花频率。
刺苍耳花粉随风散播的距离以及散播的花粉数量的测定方法为重力载玻片法。选取处于盛花期的刺苍耳植株,清除周围的其他苍耳属植物。以其为起点,在地面沿盛行风向布置涂满凡士林的载玻片。08∶00开始布设载玻片,每隔5 m布置1行,每行平行布设3张载玻片,载玻片上标记距离。布置总距离为50 m,共33张载玻片,用以接收散播于空气中的花粉粒。与此同时,用风速测量仪(AVM-01/03,Prova Instruments Inc.,中国台湾)测定全天风速,每隔10 min测定记录1次,取平均值。20∶00回收载玻片,带回实验室。在Olympus光学显微镜下观察,记录每个载玻片上承载的刺苍耳花粉粒数目[9,10]。
1.2.3 花粉活力的测定
用TTC(triphenyl tetrazolium chloride)染色法测定刺苍耳的花粉活力在1 d内的变化情况。08∶00开始,从刺苍耳植株上随机选取正处于盛花期正值散粉的10个雄花序,用载玻片收集花粉。迅速用0.5%TTC染液对花粉进行染色,并在37 ℃条件下避光静置30 min,然后在Olympus光学显微镜下进行观察,每个载玻片随机观察3个视野,统计记录每个视野中的花粉粒总数目和红色花粉粒数目。花粉活力=红色花粉粒数目/花粉粒总数目×100%。取平均值。每隔2 h检测1次,直至24∶00结束[11]。
1.2.4 柱头可授性的测定
非饱和粉煤灰抗剪强度的时间效应及其预测···················王思聪 孙德安 (1,108)
刺苍耳柱头可授性的测定采用联苯胺-过氧化氢法。于盛花期的每日12∶00,随机选取10株刺苍耳,每株植株随机选取8朵雌花,在柱头伸出之前套袋。从柱头伸出雌花序的第1天算起,每日观察柱头的生长状况,并标记天数,依次获得生长1~12 d的完整柱头,设置3组重复。截取柱头后立即置于载玻片上,然后滴加1~2滴联苯胺-过氧化氢溶液。静置1~2 min后,在Olympus光学显微镜下观察。具有可授性的柱头呈蓝色,同时柱头周围有大量气泡[11,12]。
1.2.5 柱头表面积与花粉粒横截面积的比值的测
定与计算
柱头表面积与花粉粒横截面积的比值(S/C)表示柱头可以承载的最大花粉量。标记选取开放1周左右的雌花序10个,用于测定柱头表面积。选取处于盛花期的刺苍耳植株10株,每株随机选取花粉粒3粒,用于测定花粉粒横截面积。将柱头和花粉粒分别放置于刻度载玻片上,使用光学显微镜(BX61,Olympus,日本)测量单个柱头的长度和直径及单个花粉粒的直径。
柱头表面积=(柱头长度×柱头直径×π)+(柱头直径/2)2×π
花粉粒横截面积=(花粉粒直径/2)2×π。
1.2.6 繁殖分配
随机选取果实完全成熟的植株10株,每株完整收集、编号标记,带回实验室。抖净泥土和杂质,将根系部分与地上部分分离,地上部分分为繁殖器官(包括果实和花序)和营养器官(包括茎、叶和刺),在记录刺苍耳和苍耳种群的花期时,将散粉完毕并枯萎的所有雄花序逐株分别进行收集。将繁殖器官和其余部分分装,干燥至恒重,再用电子天平(0.1 mg)测量全株和繁殖器官(果实和雄花序)的干重,记录每株的果实数量,计算平均值。繁殖分配用繁殖器官的干重与全株干重的比值表示,比值越大,说明植株分配于繁殖的物质与能量越多[13]。
1.2.7 交配系统
采用套袋实验方法,在盛花期,从刺苍耳种群中随机选取30株植株测定其交配系统。选取柱头尚未伸出的雌花序,进行以下处理(每个处理至少包括30个花序)。(1)对照:对雌花序只进行标记,不进行任何处理。(2)自花授粉:在雌花序伸出柱头前,连同与其相邻的尚未散粉的雄花序一并套袋(羊皮纸袋)。(3)异花授粉:选取柱头尚未伸出的雌花序,去除与其相邻的雄花序后套袋,待柱头成熟并具有较强可授性时,采集异株同种植物的花粉对其进行人工辅助授粉。(4)无融合生殖:选取柱头尚未伸出的雌花序,将与雌花序相邻的雄花序去雄后直接套袋[11,14],观察对照组和各处理组的结实情况。
1.2.8 数据处理
用Excel2010和SPSS19.0软件对实验数据进行处理,并进行统计分析,P<0.05则表明差异显著。
2 结果与分析
2.1 开花物候
刺苍耳单个花序的开花持续时长(花粉散播持续时间)为4~7 d。个体水平上,刺苍耳的雄花花期持续时长平均为80 d (6月中下旬到9月上旬),刺苍耳的雌花花期持续时长平均为85 d (6月下旬到9月中旬)。种群水平上,刺苍耳种群的雄花花期持续时长平均为91 d (6月中旬到9月中旬),刺苍耳种群的雌花花期持续时长平均为100 d(6月中下旬到9月下旬)(表1)。
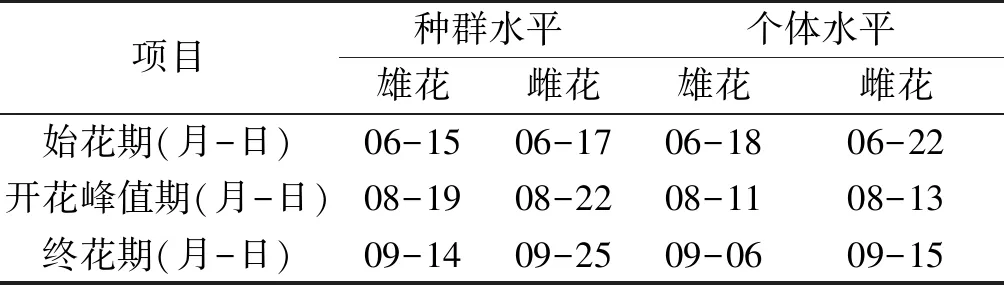
表1 刺苍耳种群水平和个体水平上的开花物候Tab.1 Flowering phenology for X.spinosum at the population and individual level
2.2 传粉媒介与风媒传粉距离和散粉量
2.2.1 访花昆虫的观察
通过定位观察,没有发现刺苍耳种群中有任何昆虫访花。因此,判断刺苍耳为非虫媒传粉植物。
2.2.2 花粉风媒散播距离以及散播的花粉数量
研究发现:刺苍耳的花粉散播呈单峰曲线模式,花粉散播的数量随着距离的增加先增加而后减少。在平均风速为0.89 m/s的微风条件下,刺苍耳花粉借风散播的最远距离可达34 m。但刺苍耳散播的花粉主要集中于距刺苍耳植株0~10 m的范围内,散播数量占其花粉总量的87%(图1)。
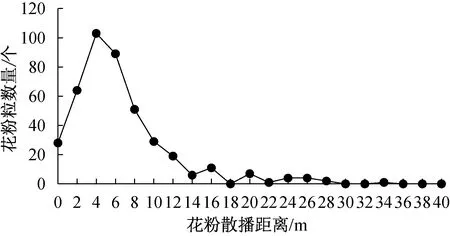
图1 刺苍耳花粉的散播距离Fig.1 Pollen-dispersal distance of X. spinosum
2.3 花粉活力和柱头可授性
刺苍耳的花粉活力在08∶00—10∶00,随着温度的升高而升高;10∶00—14∶00,几乎保持在最高水平;14∶00—16∶00,随着气温的升高开始显著下降;16∶00后,温度逐渐降低,花粉活力又开始升高,并在18∶00达到第2个高峰,此后,又开始降低;但在20∶00之前,刺苍耳的花粉活力均维持在75%以上;至23∶00,花粉活力降至0(图2)。
如表2所示,刺苍耳的柱头具有可授性的持续时间为10 d。刺苍耳雌蕊的第1个柱头伸出当天,由于雌花序还很小,尚处于生长阶段,因而柱头也特别细小,仅为1 mm左右,因而不具有可授性。随着柱头的生长,第2天柱头开始具有可授性,并一直持续到第11天。从第12天开始,柱头开始弯曲、变褐,丧失活力。
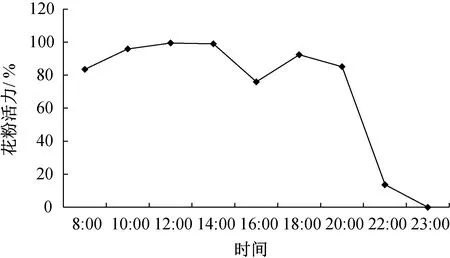
图2 刺苍耳1天内花粉活力的变化Fig.2 Changes of pollen viability of X. spinosum in one day

表2 刺苍耳的柱头可授性Tab.2 Stigma receptivity of X.spinosum
注:“+”表示柱头具有可授性;“-”表示柱头不具有可授性。
2.4 柱头表面积与花粉粒横截面积的比值(S/C)
柱头表面积与花粉粒横截面积的比值(S/C)的数据结果显示:刺苍耳的S/C值为742.8,表明刺苍耳单个柱头上可以容纳的花粉数量约为742粒。(表3)。

表3 刺苍耳的柱头表面积与花粉粒横截面积的比值(S/C)Tab.3 Ratio of the superficial area of stigma (S) to the maximum cross-sectional area of pollen grain (C) of X.spinosum
注:**P<0.05。
2.5 繁殖分配
刺苍耳繁殖器官的生物量占单株总生物量的平均比例为53%。刺苍耳单株的结实量平均为7508个。
2.6 交配系统
套袋试验结果表明,对照组(即自然授粉处理组)的30个雌花序的结实率为100%;异花授粉处理组的结实率为80%;自花授粉处理组的结实率可达96.67%;无融合生殖处理组的结实率为0(表4)。

表4 刺苍耳不同处理的结实率Tab.4 Average seed set rate of different treatments of X. spinosum
3 讨论
许多研究表明长花期有利于提升外来种的入侵能力,吴磊[15]研究发现外来入侵植物大多为花期较长的阳性植物。Goodwin等[16]研究报道加拿大的许多原产于欧洲的入侵植物的花期显著长于非入侵性的同属植物。Lloret等[17]指出在地中海的克里特岛、马耳他岛等5个岛屿上,长花期的外来植物的种群密度一般都比较高。本研究结果显示无论在个体水平,还是种群水平,无论是雄花花期的持续时长,还是雌花花期的持续时长,外来植物刺苍耳的花期持续时间(80~100 d)都显著地长于本地植物苍耳的花期时长,后者雄花与雌花花期持续时长分别为53 d和46 d[18]。长花期显著地提高了刺苍耳雄性和雌性繁殖器官的适合度,其成功授粉受精的可能性也将会大大增加[19]。此外,花期持续时间与座果数存在显著正相关关系[8]。长花期有利于提高传粉效率,从而提高座果的机会和结实量。因此,刺苍耳较长的花期将极有可能提高其结实量,而这一推测也被刺苍耳单株的平均结实量所证实:其单株的结实量平均约为7508个。而种子生产能力高的外来物种其入侵性往往也很强。因此,就结实量而言,刺苍耳具有强入侵性,必须予以高度重视。
传统观念一般认为:虫媒传粉具有高效性、普遍性和多样性等特点和优势[20],而水媒和风媒传粉则传粉效率较低,在自然系统中往往处于劣势[21,22]。然而,研究[23]发现,风媒传粉并不如传统观念认为的那样低效,其效率甚至能够超过虫媒传粉,并且,目前至少已经发现有多达65次被子植物从动物媒传粉转变为风媒传粉的实例[24,25]。植物从虫媒转变为风媒只需要进化出相对简单的花部特征[26]。因此,风媒传粉极有可能在某些方面具有明显的优势,成为一些被子植物传粉模式进化的方向。本研究结果表明刺苍耳为风媒传粉植物,在风速为0.89 m/s的微风条件下其花粉扩散距离便可达34 m;且在距离植株10 m的范围内花粉散播量占到其散播花粉总量的87%。由于刺苍耳植株在群落中往往呈高密度聚集性分布,因此其散播出的大量花粉将显著地增加其异交成功的可能性,有助于产生更多的异交种子,提高其后代的环境适应性[27]。
植物顺利完成传粉受精需要经过一系列复杂的过程[28],而这一过程受到柱头可授性、花粉活力等因素的影响。花粉活力是指花粉具有存活、生长、萌发的能力[29],直接影响植物的授粉受精和结实率[30]。不同物种,其花粉具有活力的时长也不同,水稻花粉散播出大约10 min后就几乎全部失去了萌发能力;菊花花粉的寿命则为1~2 d[31]。不同物种,其新鲜花粉的活力也不相同。偃麦草在散粉期的花粉活力只有10%;而新鲜节瓜散粉期的花粉活力则能够高达90%[32]。而柱头可授性是指:柱头接受有活性的花粉和维持亲和花粉萌发的性能[33]。柱头可授性的强弱以及持续时长对于植物成功受精结实具有十分重要的影响。研究[34]证实,在柱头的最佳可授期(可授性最强时期)授粉,植物的结实率或结子率都明显高于其他时期。本研究结果表明:刺苍耳从08∶00—20∶00的12 h里,其花粉活力均可维持在75%以上,最高可达99%。其柱头具有可授性的时间则达到了10 d,显著地高于同属另一外来入侵物种意大利苍耳的柱头可授性时长(7 d)[35]。刺苍耳不仅可以在长达12 h里维持75%以上的花粉活力,而且还能维持较长时间的柱头可授期,从而大大提高了其成功繁殖的机会,显著提高了该植物的入侵力[4]。此外,前人对平榛和平欧杂种榛柱头可授性的研究[36]表明,当柱头的长度达到最大值时,其表面附着的花粉最多,显著提高了其繁殖成功的概率。刺苍耳的单个柱头上可以容纳的花粉数量约为742粒,极大地增加了柱头成功捕获空气中花粉的机会,大大提高了其雌蕊成功受精的概率。
繁殖分配是植物繁殖生物学研究的核心内容之一。繁殖分配是指植物分配到繁殖器官的生物量占总生物量的比例[37]。对被子植物而言,较高的繁殖分配就意味着母体优先投资更多资源用于繁殖,可使入侵植物产生更多的种子,从而快速地在新生境中定殖成功[38,39]。外来植物的繁殖能力和入侵力之间往往呈正相关关系,而入侵植物往往比本地植物具有更高的繁殖分配[40],这与本研究的结果是一致的。刺苍耳的繁殖分配达到了53%,说明刺苍耳将超过半数的资源投资用于繁殖产生后代,单株的果实数量平均约为7508粒,使其在入侵区域中能够更快更容易地建立种群并快速扩散[4,41]。
自交亲和是许多外来植物繁育系统的共同特征,自交亲和者,在其入侵新栖息地的初期阶段,即使传入的个体数量很少,也能够在短时间内产生足够多的种子,有利于其在新生境中的种群建立。根据调查,入侵我国的菊科植物中,有将近2/3的物种能够进行自交结实[42]。而异交亲和则可以显著地增强植物后代的生活力,主要表现为:相对于亲本,后代的繁殖力、生长势等性状明显增强、总生物量明显提高,这也被称为杂种优势[43]。本研究结果表明:刺苍耳的自花授粉处理的结实率显著地高于异花授粉处理的结实率,这表明刺苍耳趋于自交,而自交能够降低外部环境对外来种繁殖的干扰,从而达到迅速成功繁殖的目的[44];并且,刺苍耳也可以进行异交,异交结实率达到了80%,这对刺苍耳避免自交衰退,改善其后代的适合度具有积极意义。而在去雄套袋处理中没有结出任何种子,则说明刺苍耳不具有无融合生殖的特性[11](表4)。自交和异交相结合的繁殖策略大大提高了刺苍耳的繁殖能力,也提高了其入侵性[4]。
外来入侵植物刺苍耳在开花物候、传粉媒介、花粉散播距离、柱头可授性、花粉活力、柱头表面积与花粉粒横截面积的比值、繁殖分配、交配系统等繁殖生物学特性指标上都表明了:刺苍耳具有很强的有性繁殖能力,这可能是其成功入侵的重要原因之一。
