盐胁迫对2种菠萝蜜属植物幼苗生长及光合荧光特性的影响
2019-07-06苏兰茜白亭玉鱼欢吴刚谭乐和
苏兰茜,白亭玉,鱼欢,吴刚,谭乐和
(中国热带农业科学院香料饮料研究所,海南万宁 571533)
0 引言
【研究意义】盐碱土约占陆地总面积的25%,是分布最广的土壤类型之一。我国滨海盐碱地面积较大,随着人口增长与耕地减少的矛盾日益突出,各类盐土资源,特别是我国海岸带盐土作为一种重要的土地后备资源,亟待开发和利用[1]。筛选耐盐经济植物,发展盐土农业,是利用盐碱地的有效途径[2-3]。然而滨海盐碱地区土壤条件苛刻,适宜栽培的植物种类相当有限,严重限制了我国滨海盐碱地的开发利用。例如,西沙各岛气候环境、土壤与内地差异很大,土壤全是珊瑚风化的细砂,土地贫瘠,淡水资源紧缺,驻岛官兵的粮食和饮用水都依靠海南岛运输上岛。因此培育和引种优良耐盐碱作物,对开发利用滨海土地资源具有重要意义。【前人研究进展】菠萝蜜和面包果都是桑科(Moraceae)菠萝蜜属(Artocarpus)特色木本粮食作物。菠萝蜜果实中含有丰富的糖分、蛋白质和脂肪,营养价值高,种子富含淀粉。面包果果肉及种子富含蛋白质、碳水化合物及丰富的膳食纤维,煮熟后可食,可代粮食。二者均是可开发利用的木本粮食作物资源。目前,国内外关于植物对盐胁迫的响应开展了大量的研究,如盐胁迫可抑制大豆、芦苇幼苗生长、SPAD值、光合荧光参数,对盐敏感品种影响程度大于耐盐品种[4-5],而低浓度盐胁迫对植物的影响不显著,高浓度盐胁迫对植物的抑制作用显著[6]。一般认为盐胁迫对植物的伤害主要表现在渗透胁迫、离子毒害和营养失衡 3个方面。渗透胁迫会抑制根系对水分吸收,进而降低光合作用和蒸腾作用效率;植物体内Na+和 Cl-的大量积累打破了细胞内的离子平衡,破坏细胞膜结构并致使代谢紊乱;盐离子与营养元素相互竞争则导致植物体内营养亏缺,从而影响植物的正常生长和发育[7]。【本研究切入点】过去对盐生植物的研究多集中在其分布、生理特征以及种子萌发等方面[8-14],而热带木本粮食作物的抗盐能力未见报道,盐胁迫下菠萝蜜和面包果叶片光合荧光特性的差异也不明确。【拟解决的关键问题】本研究以菠萝蜜和面包果幼苗为试验材料,揭示盐胁迫与光合作用及相关指标之间的关系,为热带盐碱地开发利用,发展岛礁特色农业(热带木本粮食作物)提供参考。
1 材料与方法
1.1 供试材料
1.1.1 供试海沙 供试海沙采自海南省万宁市兴隆镇石梅湾(18°39′43′N;110°15′47′E),该地区属热带海洋性季风气候,年均降水2 135 mm,母质为近代浅海沉积物[15],海沙pH 6.85,速效磷30 mg·kg-1,速效钾 12.68 mg·kg-1,碱解氮 8.01 mg·kg-1,有机质0.5 g·kg-1。
1.1.2 供试材料 供试海水为石梅湾海水,用电导盐度计测定海水盐度为34.0‰。供试肥料为牛粪有机肥(N,15.6 g·kg-1;P2O5,47.2 g·kg-1;K2O,12.9 g·kg-1),购自江苏禾喜生物科技有限公司。供试菠萝蜜苗为马来西亚1号嫁接苗,面包果苗为中国热带农业科学院香料饮料研究所自主选育的4号品种。
1.2 试验设计
盆栽试验于2018年3—6月在中国热带农业科学院香料饮料研究所温室进行。试验用土过筛(2 mm)后装入塑料盆(直径18 cm,高30 cm)中,每盆装土6 kg,按照1%的比例均匀拌入牛粪有机肥60 g,移栽1株三叶一心的供试菠萝蜜苗/面包果苗,每天浇不同盐浓度的海水200 mL。盐浓度共设5个水平:T0(不添加海水的自来水,自来水盐度为0.05‰,此处忽略不计)、T1(盐浓度为3.4 g·kg-1,900 g水和100 g海水混合,后续浓度配置同理)、T2(盐浓度为 10.2 g·kg-1)、T3(盐浓度为 17.0 g·kg-1)、T4(盐浓度为23.8 g·kg-1)。为防止菠萝蜜和面包果产生盐激反应,本试验采用每d增加3.4 g·kg-1渐增的方式达到预设浓度。整个试验过程,通过称重法将土壤水分维持在田间持水量的75%左右,每天傍晚18:00,用五点法采集土样进行含水量测定,及时补水。15 d后用自来水淋洗3—5次,按照之前的方式再加入不同盐浓度的海水,以保持土壤盐分浓度的相对稳定。每个水平设置3个重复,每个重复4株,总计60株。整个试验培养30 d后测定相关指标。
1.3 项目测定与方法
1.3.1 生长及生理指标测定 种植 4个月后开始测定各个指标,每个处理随机选取6株长势较一致的菠萝蜜苗和面包果苗测定生长指标,苗高为茎基部到叶片最高点的高度;倒三叶的叶面积采用长宽系数法测定(叶面积=长×宽×0.75,未展开叶系数 0.5)[16];茎粗用游标卡尺测定,统一测定距茎基部2 cm处。叶绿素含量用SPAD-502便携式叶绿素速测仪测定。植物样品收获后,用去离子水将植株冲洗3次,分成地上部和地下部,置于105℃烘箱中杀青30 min,转至65℃烘至恒重,并称取其干质量。根冠比=根干重/地上部干重。
1.3.2 光合指标测定 采用LI-6400光合测定仪,选择向阳、生长均匀一致的倒三叶测定叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)等光合指标。气孔限制值(Ls)算式:Ls=1-Ci/Ca(式中:Ci为胞间 CO2浓度;Ca为大气CO2浓度)[17]。测定时间为天气晴朗的上午 9:00—11:00,CO2浓度为(385±5)μmol·mol-1,测定光强为 1 500 μmol·m-2·s-1,叶片温度为(25.5±2)℃,空气相对湿度维持在45%左右。采用标准叶室,开放式气路,流速均设定为500 μmol·s-1。每个处理随机选取6株长势较一致的幼苗,选取相同部位的叶片,每叶重复测定3次。
1.3.3 叶绿素荧光参数的测定 采用超便携式调制荧光仪 Mini-Image-PAM 测定,每个处理随机选取 6株叶片完整的幼苗,选取倒三叶测定,每叶重复测定3次。叶片经暗处理30 min后,测定初始荧光(Fo)、最大荧光(Fm)以及最大光量子效率(Fv/Fm);自然光下活化30 min后,测定光适应下的最小荧光Fo′、最大荧光Fm′、稳态荧光Fs。
1.3.4 土壤理化性质测定 土壤样品参照《土壤农化分析》[18]进行理化性质测定。
1.4 数据分析
在SPSS 19.0中,使用Kolmogorov-Smirnov检验和Levene's检验对所有数据进行正态分析和方差齐性检验。采用单因素方差分析(ANOVA)进行数据比较,利用Duncan新复极差法检验处理间差异的显著性水平(P<0.05)。使用 R软件(3.3.2)里的corrplot程序包进行相关性分析,BioEnv程序挑选对菠萝蜜和面包果生物量变化具有显著影响的光合参数,再通过vegan程序包进行方差分解分析(variance partitioning analysis,VPA)来研究盐胁迫与光合参数以及它们共同对菠萝蜜和面包果生物量变化的贡献。
2 结果
2.1 盐胁迫对菠萝蜜和面包果幼苗生长的影响
从表1可以看出,与对照相比,T1处理显著抑制了菠萝蜜幼苗的株高、茎粗和叶面积生长,抑制率分别达 23.75%,18.23%和 229.31%,T1与其他盐浓度处理差异不显著。T2处理显著抑制菠萝蜜叶片的叶绿素合成,抑制率达45.63%,T2与T3和T4处理无显著性差异。T1处理对菠萝蜜幼苗生物量的积累有显著抑制,对地上部干重、地下部干重和总干重的抑制率分别达32.09%,25.66%和30.02%,T4处理的幼苗生物量最低。与对照相比,T2—T4处理显著增加菠萝蜜幼苗的根冠比,增加幅度达44.68%,与T3和T4处理无显著性差异。
盐胁迫下面包果地上部生长表现明显不同于菠萝蜜,除T4处理的叶面积外,随着盐胁迫程度的增加,面包果地上部生长指标有下降趋势,但均与对照差异不显著。盐胁迫对面包果叶片叶绿素含量无显著影响。与对照相比,T2处理显著减少面包果地上部干重、地下部干重和总干重,减少比例分别达37.57%,27.54%和35.03%,T3处理的生物量最低。T1处理与对照相比显著减少地下部干重,减少比例为15.61%,对地上部干重和总干重的影响不显著。与对照相比,T4处理显著增加幼苗根冠比,比例达36.36%,其他盐浓度处理与对照无显著性差异。
2.2 盐胁迫对菠萝蜜和面包果幼苗光合特性的影响
2.2.1 对净光合速率的影响 随着盐胁迫程度的增加,菠萝蜜叶片的净光合速率急剧下降。与对照相比,T2处理对净光合速率的抑制率达 84.41%,T2与T3和T4处理无显著性差异。T1处理较对照降低净光合速率的比例为9.56%(P<0.05)。面包果叶片的净光合速率随盐胁迫程度增加呈逐渐下降的趋势,T1—T4处理下降的比例分别为 21.79%、39.01%、61.02%和 69.19%,其中 T1和 T2盐浓度处理间无显著性差异,T3和T4处理间差异不显著(图1-a)。

表1 盐胁迫下菠萝蜜和面包果幼苗生长指标变化Table 1 Growth indexes of the seedlings of jackfruit and breadfruit under salt stress
2.2.2 对叶片气孔导度的影响 随着盐胁迫程度的增加,菠萝蜜叶片气孔导度变化趋势与净光合速率趋势相似。T2处理对气孔导度的抑制作用最强,较对照抑制率达96.80%,T2与T3和T4处理无显著性差异。T1处理较对照降低气孔导度的比例为 12.29%(P<0.05)。面包果叶片的气孔导度随盐胁迫程度增加呈逐渐下降的趋势,T1处理的气孔导度较对照下降的比例为45.43%(P<0.05),T1处理显著高于T3和T4处理(图1-b)。
2.2.3 对胞间CO2浓度的影响 菠萝蜜和面包果叶片胞间CO2浓度随盐胁迫程度的增加均呈先下降后上升的趋势,其中菠萝蜜苗在 T2处理时达最低值,面包果苗在T3处理时达最低值。T1处理的菠萝蜜叶片胞间CO2浓度与对照无显著性差异,面包果T1处理叶片胞间 CO2浓度较对照下降的比例为 15.38%(P<0.05,图1-c)。
2.2.4 对叶片蒸腾速率的影响 随着盐胁迫程度的增加,菠萝蜜叶片蒸腾速率变化趋势与净光合速率趋势相似。与对照相比,T2处理对蒸腾速率的抑制率达93.58%,T2与T3和T4处理无显著性差异。T1处理较对照降低蒸腾速率的比例为 6.97%(P<0.05)。面包果叶片的蒸腾速率随盐胁迫程度增加呈逐渐下降的趋势,T1处理的蒸腾速率较对照下降的比例为 41.73%(P<0.05),显著高于 T3和 T4处理(图1-d)。
2.2.5 对叶片气孔限制值的影响 T2—T4处理的菠萝蜜叶片气孔限制值显著高于对照和 T1处理。面包果的变化幅度较菠萝蜜小,随着盐胁迫程度的增加,各处理的气孔限制值呈先增加后降低的趋势,均显著高于对照,T3处理的气孔限制值显著高于T1处理(图1-e)。
2.3 盐胁迫对菠萝蜜和面包果幼苗叶绿素荧光特性的影响
表2结果表明,菠萝蜜叶片光化学猝灭系数(qP)、潜在最大光能转换效率(Fv/Fm)、实际光能转换效率(ΦPSII)和相对电子传递效率(rETR)在T0—T3处理之间无显著性差异,在 T4处理中显著低于对照,减少的比例分别为 7.37%、12.66%、23.61%和23.40%。与对照相比,T4处理对叶片光系统Ⅱ活性(Fv/Fo)的抑制率达36.63%。盐胁迫对叶片的非光化学淬灭系数(NPQ)无显著抑制作用。低浓度盐胁迫对面包果叶片的叶绿素荧光特性无显著影响。T4处理面包果叶片的光化学猝灭系数、非光化学淬灭系数、实际光能转换效率和相对电子传递效率较对照显著降低,减少的比例分别为4.26%、55.56%、5.63%和5.90%。
2.4 盐胁迫下菠萝蜜和面包果幼苗生物量与光合参数相关性分析
图2表明,菠萝蜜和面包果幼苗总干重与叶绿素含量、净光合速率、气孔导度和蒸腾速率呈显著正相关关系,根冠比与叶绿素含量、净光合速率、气孔导度和蒸腾速率呈显著负相关关系;叶绿素含量、净光合速率、气孔导度、胞间CO2浓度和蒸腾速率两两之间呈显著正相关关系。

图1 盐胁迫下菠萝蜜和面包果幼苗叶片的光合参数Fig. 1 Photosynthetic parameters in leaves of jackfruit and breadfruit seedlings under salt stress (mean±SE)
2.5 盐胁迫、叶绿素含量、光合参数对菠萝蜜和面包果幼苗生物量变化的贡献
以菠萝蜜和面包果幼苗生物量为基础,探究盐胁迫、叶绿素含量和光合参数对幼苗生物量的影响。使用 Bioenv挑选出对幼苗生物量起关键性作用的光合参数为:净光合速率和叶绿素含量。变量分解分析(VPA)的结果如图3所示,盐胁迫、净光合速率和叶绿素含量以及它们之间交互作用解释幼苗生物量变化的比例为 35.6%。对于单个因子来说,盐胁迫、净光合速率和叶绿素含量可以解释菠萝蜜苗生物量变化的比例分别为-2.1%(P=0.706)、-1.3%(P=0.5)和2.5%(P=0.173);盐胁迫与净光合速率共解释幼苗生物量变化的比例为 6.3%(P=0.002),净光合速率与叶绿素含量共解释幼苗生物量变化的比例为 10.3%(P=0.001),盐胁迫与叶绿素含量共解释幼苗生物量变化的比例为-0.3%(P=0.001),3个因子共解释幼苗生物量变化的比例为20.2%(P=0.001)。

表2 盐胁迫下菠萝蜜和面包果幼苗叶片的叶绿素荧光特性Table 2 Chlorophyll fluorescence in leaves of jackfruit and breadfruit seedlings under salt stress
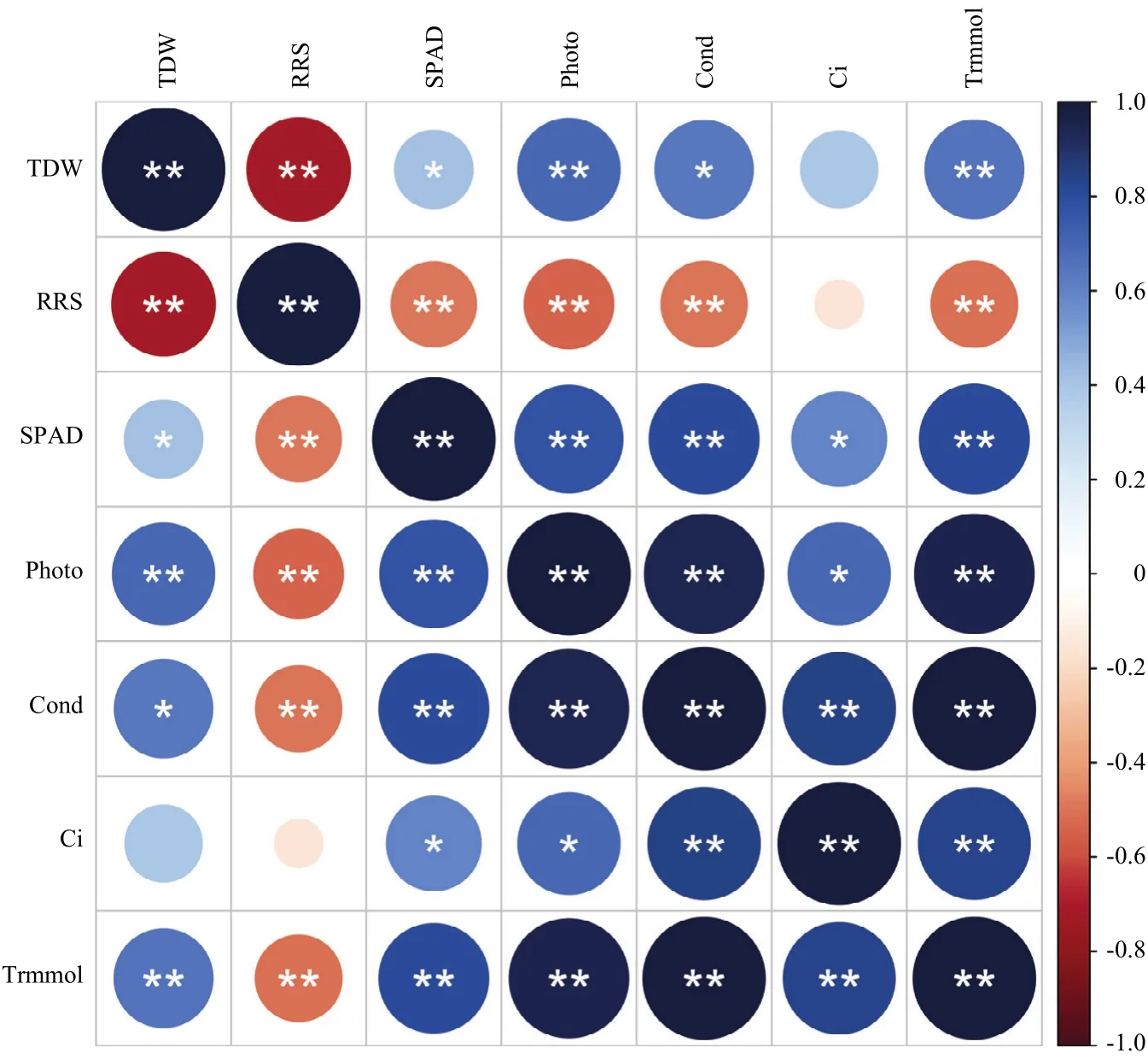
图2 盐胁迫下菠萝蜜和面包果幼苗生物量与光合参数相关性分析Fig. 2 Correlation between the biomass and photosynthetic parameters of jackfruit and breadfruit seedlings under salt stress
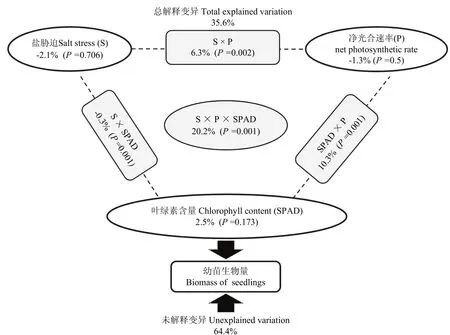
图3 菠萝蜜和面包果幼苗生物量变化的驱动力分析Fig. 3 Variation partitioning analysis (VPA) was used to quantify the effects of salt stress (S), net photosynthetic rate (P), and chlorophyll content (SPAD), and all of them on jackfruit and breadfruit seedlings biomass
3 讨论
作物对盐胁迫响应最敏感的生理现象是影响生长[19],本研究结果表明盐胁迫显著抑制菠萝蜜地上部的生长,而对面包果的抑制作用不显著。谢德意等[20]和杨淑萍等[19]对棉花的研究发现盐胁迫对棉花幼苗生长产生抑制作用。对于盐生植物来说,盐分浓度在没有超过生长阈值时,同化作用不会被抑制,甚至在较低时还会被促进[21],如木麻黄[22]、胡杨[23]等很多树种也表现出类似的特性。本试验发现,菠萝蜜对盐胁迫敏感,所有加盐处理的菠萝蜜生长指标及生物量均显著低于对照,但面包果的生长指标与对照相比有下降趋势,但差异不显著。面包果地上部和整株干重在T1处理中与对照无显著差异,而后随盐浓度的增加显著降低,地下部干重对盐胁迫较敏感,这可能说明盐分对菠萝蜜和面包果的作用方式不同。从本试验的结果来看,面包果较菠萝蜜耐盐性更强,T1盐浓度为面包果生长的最适浓度,而 T4盐浓度可能为菠萝蜜和面包果的最大耐盐阈值。
叶绿素含量与植物叶片光合能力密切相关,是衡量植物抗逆性的重要生理指标之一[24]。盐胁迫能增强叶绿素酶活性,加剧叶绿体的降解和抑制其合成,因此叶绿素含量变化能反映盐胁迫对植物的伤害程度[25-27]。本研究结果表明,轻微的盐胁迫能使叶绿素含量略有增加,且面包果的增幅较大;随着盐胁迫程度增加,菠萝蜜的叶绿素含量显著下降,该结果与前人在大豆[28]和高粱[29]上的研究结果类似。而面包果叶绿素含量变化不大,对盐胁迫不敏感。根冠比的值反映了植物在逆境条件下对生物量的分配策略,植物不同部位对盐分的敏感程度不同,导致根冠比值发生变化[30]。表1结果表明,盐胁迫下,菠萝蜜和面包果的根冠比呈增加趋势,高浓度盐胁迫下的根冠比显著高于对照。可见高盐胁迫导致根系养分吸收能力及叶片同化产物的分配明显改变[31]。根冠比越大,表明根系分配到的同化产物更多[32],有利于根系的快速扩张。本试验结果表明,随着盐浓度增加,根冠比呈增加趋势,且菠萝蜜的根冠比值大于面包果,表明盐胁迫下,菠萝蜜根系扩张的速度更快,因此竞争的光合产物较地上部多,影响了地上部的生长速度。这种分配模式有利于增加根系对水分和营养的获取,从而增强植物体的生长能力,稀释细胞内的盐分[33]。
盐胁迫显著抑制菠萝蜜和面包果叶片净光合速率、气孔导度和蒸腾速率,这与前人对大麦[34]和高粱[29]的研究结果一致。并且盐胁迫下,面包果光合参数下降幅度低于菠萝蜜,说明面包果的耐盐能力相对较强。盐胁迫对植物生长和代谢的影响是多方面的,其中光合作用的变化最明显。影响植物光合作用的因子主要分为气孔因子和非气孔因子[35]。大量研究表明,盐胁迫导致植物叶片细胞程序性死亡、输导组织受损、光合速率降低、气孔阻力升高,认为气孔限制是盐胁迫下植物光合速率下降的主要原因[36],而有些研究表明,非气孔限制是盐胁迫后光合速率下降的主要原因,盐胁迫下RuBP羧化酶效率降低和光合作用表观量子效率下降导致光合速率降低[37]。当光合速率下降,若胞间CO2浓度降低,而气孔阻力升高,表明光合速率降低主要原因是气孔限制,否则为非气孔限制[38]。因此本试验中,盐胁迫下菠萝蜜和面包果叶片净光合速率下降,气孔导度降低,限制了CO2向叶绿体的输送。当盐浓度在T1—T2时,叶片胞间CO2浓度 Ci随气孔导度Gs的减小而降低,气孔限制值Ls增加,净光合速率 Pn降低,这可能是由于盐胁迫刺激引起渗透胁迫,导致气孔关闭,叶肉细胞间隙中CO2的消耗速率减慢,从而导致Ci降低。由此表明,当盐浓度为T2时,菠萝蜜和面包果叶片Pn下降的主要原因为气孔限制,这与李学孚等[6]研究葡萄的结果一致。两种植物的 Gs值随着盐浓度的增加呈现缓慢下降的趋势,但Ci却呈上升趋势,Ls逐渐降低,这可能是高盐胁迫下细胞呼吸作用短暂加强,而细胞中积累了大量盐离子,破坏了叶绿体结构和叶片光合器官,造成叶绿素含量降低、叶肉细胞的光合活性下降,说明此时光合速率降低的主要原因是非气孔限制。因此,菠萝蜜和面包果随着盐浓度的增加,光合作用的限制均由初期的气孔限制主导而逐渐转变为非气孔限制主导,但限制两者光合作用主导因子的转换盐浓度不同,菠萝蜜为T2盐浓度,而面包果则为T3盐浓度,这可能再次证实菠萝蜜和面包果对盐分的耐受程度不同。
光合速率下降必然会影响植物对光能的吸收、传递和转化,尤以光化学活性下降最为突出[39]。研究表明,叶绿体光合机构在高浓度盐胁迫下易被破坏,导致光系统Ⅱ原初光能转换效率、PSⅡ潜在活性降低[19,29]。本试验也发现,在低浓度胁迫下,菠萝蜜叶片光化学猝灭系数(qP)、光系统Ⅱ活性(Fv/Fo)、潜在最大光能转换效率(Fv/Fm)、实际光能转换效率(ΦPSⅡ)和相对电子传递效率(rETR)在处理间和对照均无显著性差异,在高浓度胁迫下,除NPQ与其他处理无显著差异外,其他荧光参数均有所下降,表明菠萝蜜叶片发生了光抑制[40],说明高盐胁迫影响了菠萝蜜叶片光系统Ⅱ的激发能分配方式,通过提高热耗散消耗过多激发能来适应盐胁迫环境。而面包果的Fv/Fo和Fv/Fm变化不显著,其他荧光参数在高盐胁迫下均有所下降,说明面包果在盐胁迫下能够有效保持光系统Ⅱ活性和潜在最大光能转换效率,使光合机构趋于稳定,防止净光合速率过度下降,这可能是面包果对盐胁迫环境适应的一种保护机制。
叶绿素是影响光合作用速率的主要因素[41]。本研究结果显示叶绿素含量与光合参数呈显著正相关关系,这与冯国郡等[41]在甜高粱上的研究结果基本吻合。光合作用对植物干物质积累具有重要作用[42]。本试验结果显示,菠萝蜜和面包果幼苗总干重与叶绿素含量及光合参数呈显著正相关关系,也证实较强的光合作用和叶绿素含量更能促进作物总干物质的积累。盐胁迫对菠萝蜜和面包果幼苗叶绿素含量、净光合速率和生物量有显著影响。VPA分析表明,盐胁迫、叶绿素含量和净光合速率两两间及三者交互作用解释幼苗生物量变化的比例均达显著性。其中三者交互作用解释幼苗生物量变化的比例最大。因此,菠萝蜜、面包果生物量随着盐胁迫程度的增加而降低,与叶片叶绿素含量等光合参数的下降有密切联系。
4 结论
盐胁迫显著抑制菠萝蜜幼苗生长、叶绿素含量和生物量积累,而面包果的生物量在高浓度盐胁迫下较敏感。盐胁迫降低了菠萝蜜、面包果叶片的光合参数,且菠萝蜜的下降幅度显著大于面包果。导致光合参数降低的原因可能是,菠萝蜜和面包果随着盐浓度的增加,光合作用的限制均由初期的以气孔限制为主导因子而逐渐转变为以非气孔限制为主导因子。高浓度盐胁迫下,菠萝蜜幼苗的qP、Fv/Fo、Fv/Fm、ΦPSⅡ和rETR均有所下降,而NPQ则有所上升,说明高盐胁迫会抑制菠萝蜜幼苗光系统Ⅱ活性和光能转换效率,增加光系统Ⅱ非辐射能量的耗散来适应盐胁迫环境。而面包果的光合系统对盐胁迫的防卫能力明显优于菠萝蜜。盐胁迫、叶绿素含量和净光合速率以及它们之间交互作用解释幼苗生物量变化的比例为 35.6%。光合荧光参数可准确、快速地判断菠萝蜜和面包果幼苗对盐分的耐受性。
