原始红松林退化演替后土壤氮矿化特征变化
2019-07-05张秀月付岩梅冯富娟
张秀月,付岩梅,刘 楠,冯富娟
东北林业大学生命科学学院, 哈尔滨 150040
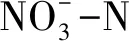
近些年,针对森林土壤氮矿化研究多集中于土壤氮转化速率、土壤氮矿化影响因子、温度敏感性Q10等方面[5- 6]。土壤氮矿化速率决定了土壤中用于植物生长的氮素的可利用性,是森林生态系统氮素循环最重要的过程之一[7]。影响土壤氮矿化速率因素众多,其中土壤的温度和含水量是影响土壤氮矿化的关键因素[8- 9],而且,弄清温度和水分因子对特定森林土壤氮矿化的影响及其交互效应,对维持土壤氮素供应以及植物生产力具有指导意义[10]。温度作为影响土壤氮矿化速率的主控因子,温度敏感性(Q10)已被广泛用作N循环模型中的一个重要生态参数,用以描述温度升高10℃时土壤氮矿化的响应[11]。以往众多的研究曾认定Q10是恒定值2,但Wetterstedt等人却认为,Q10在不同的生态条件下存在较大差异[12],而且得到了越来越多学者的肯定[13]。因此,了解Q10的空间格局和影响因子对于精确评估森林土壤有效氮具有重要意义[14]。
2017的《中国统计年鉴》表明,我国现有森林面积约208×104km2,其中68%为中幼龄次生林,显然,中幼龄为主的天然次生林将会发挥重要的生态功能。以红松(Pinuskoraiensis)为建群种的红松阔叶混交林是中国东北东部山区的地带性顶极植被,对于区域生态环境具有重要影响,还是环球北方森林的组成部分,该区气候寒冷、土壤潮湿、有岛状永久性冻土存在,属于全球气候变化敏感区域。但由于日伪时期的掠夺式采伐及建国初期生产建设的需要,原始红松林破坏严重,取而代之的是大面积的的中幼龄次生阔叶林。顶极群落向次生阔叶林的演变意味着人为干扰下的退化演替,森林生态功能必然发生变化[15]。顶极群落中,占优势的晚期演替树种往往对土壤有着深刻的改造与适应,次生林中缺少了建群种红松,必然会引起包括土壤氮营养条件在内的生境演变[16],但对此类问题的系统研究仍属空白。本文以原始红松林为对照,采用好气培养方法,研究其退化演替为次生阔叶林后土壤氮矿化水平的变化,分析两者的氮矿化潜力,揭示温度和含水量变化对土壤矿化速率的影响,进而科学评价原始红松林退化演替后中幼龄次生林后土壤肥力及初级生产力水平的变化。
1 材料与方法
1.1 试验地概括
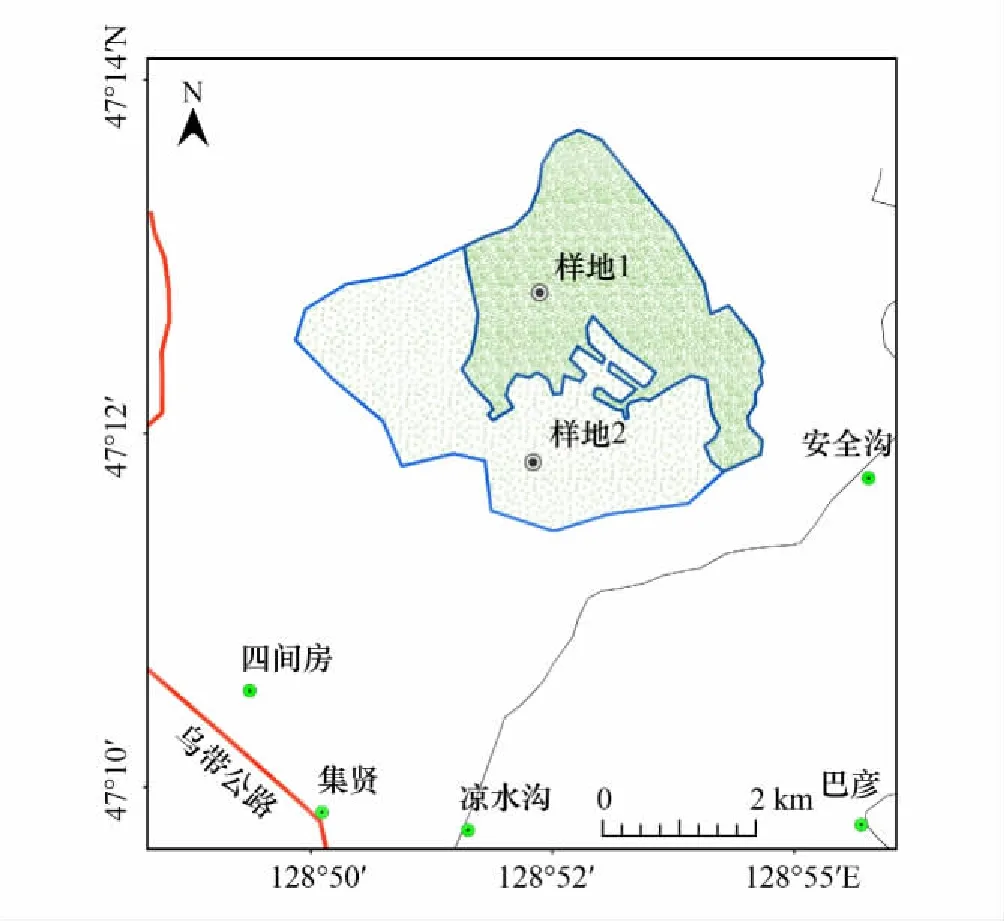
图1 样地的分布情况Fig.1 Distribution of sampling sites样地1:原始红松林;样地2:次生阔叶林
试验地设置在隶属于黑龙江省伊春市带岭区东北林业大学的凉水国家级自然保护区内,保护区地处小兴安岭山脉的东南部-带岭支脉的东坡,地理位置为47°07′39″N—47°14′22″N、128°48′30″E—128°55′50″E,凉水国家级自然保护区属温带大陆性季风气候,气候特点是春季大风天多,降水较少;因该区纬度较高,年均气温约4℃,年均最低气温-4.6℃,年均最高气温8.6℃,年均降水日数120—150天,年均降水量676mm,积雪期130—180天,年均相对湿度78%。该区是保存着最为典型和完整的原始红松混交林分布区之一,既有从未采伐过的原始林相,也有经皆伐和火烧后发生的次生林相,代表了森林发生、演替的各个阶段,样地分布情况如下。
本研究选取的样地1和2在20世纪60年代之前为连续分布的原始红松林,1961年样地2遭到一次皆伐,地上的乔木和灌木几乎都被伐,经过近60年的演替形成了现在的次生阔叶林(图1)。样地1面积约11.7 hm2,样地2面积约9.3 hm2,2块样地相毗邻,海拔、坡向、土壤母质等生态因子一致,样地树种组成概况如下所示(表1)。
1.2 样地设置及样品采集
2017年7月中旬在原始红松林、次生阔叶林两个大样地分别设立3个重复10 m×10 m正方形样方,用内径5 cm的土钻在样地按照“S”型曲线随机取10个样点,取0—20 cm土样,去除地表凋落物层及土壤中可见植物根系和动植物残体,用无菌袋将取出的土芯封袋保存,放入有冰袋的泡沫箱中带回实验室。将各重复样方内重复样品充分混合成一个混合土样,过2 mm土筛。经过预处理的部分土样风干,部分于-4℃条件下冷藏。

表1 试验地概况
1.3 室内培养实验
培养实验开始前,先采用烘干法测定土壤含水量,经过含水量换算后,再称取相当于干土质量土样30 g装入150 mL锥形瓶,加蒸馏水分别调节至饱和持水量(WHC)的20%、40%、60%、80%、100%。将样品分别放入4℃、12℃、20℃、28℃和36℃恒温恒湿培养箱培养30d,每个处理3次重复,共计150个样品。测定培养前后土壤铵态氮、硝态氮含量。在培养过程中,每隔2—3 d采用称重法给土壤样品补水1次。
1.4 土壤理化测定
土壤总有机质、土壤全碳、全氮测定采用Multi N/C®3100 (AnalytikJena, Germany);全磷用硫酸-高氯酸-钼锑抗比色法;速效磷用碳酸氢钠-钼锑抗比色法;速效钾用乙酸铵-火焰光度法;含水量用105℃,24 h烘干;pH值用pH计法(水∶土=10∶1);容重用环刀法;铵态氮和硝态氮分别用靛酚蓝比色法和酚二磺酸法。
1.5 计算方法
(1)
(2)
(3)
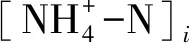
土壤氮矿化速率与温度之间的关系采用指数模型拟合:
Rmin=AeBT
(4)
土壤氮矿化的温度敏感性(Q10)计算方法如下:
Q10=exp10B
(5)
Rmin净氮矿化速率,T为培养温度(℃),A为基质质量,B为温度反应系数。
气候要素的趋势变化一般采用一元线性回归模型描述,即:
y=a+bx
(6)
式中,y为气候要素序列,x为时间序列(本文中为1976—2016年),b为线性趋势项,10b即为气候要素每10年的气候倾向率[17],用于定量分析气候要素变化。此次分析数据来自伊春市气象局。
1.6 数据处理与分析
采用单因素方差分析(One-way ANOVA)检验不同森林类型间土壤理化特征、基质质量(A)及Q10显著性差异;采用多因素方差模型(Multiple-way ANOVA)分析培养温度、培养湿度、林型及其交互作用对土壤氮转化速率的影响。数据分析采用SPSS19.0,制图使用R studio和origin软件,显著性水平设为P<0.05。
2 结果与分析
2.1 退化演替后土壤理化性质的变化
除土壤容重外,原始红松林和次生阔叶林0—20 cm表层土的有机质、总碳、总氮、铵态氮、硝态氮、C/N、总磷、速效磷、速效钾、pH值均差异显著(P<0.05);原始红松林的铵态氮高于次生阔叶林,包括硝态氮在内的其余指标则均低于次生阔叶林(表2)。

表2 土壤理化特性(n=6)
注:n为样本数,表中数据为平均值±标准差,同列不同小写字母表示差异显著(P<0.05)
2.2 退化演替后土壤氮转化速率的变化
土壤氨化和硝化过程是土壤氮素转化的核心[18],一般采用氮素转化速率(净矿化速率、净氨化速率、净硝化速率)表示。本研究比较了不同温度及湿度下原始红松林和次生阔叶林的土壤氮转化速率,两者的净氨化速率、净硝化速率、净矿化速率并没有表现出一致的变化趋势。总体来说,原始红松林净氨化速率高于次生阔叶林(图2),而净硝化速率和净矿化速率均低于次生阔叶林(图3、图4)。表3中,原始红松林和次生阔叶林的土壤氮转化速率差异显著,培养温度和湿度及两者的交互作用均对土壤氮转化速率影响极显著(P<0.001,表3)。

图2 不同林型、不同培养温度和培养湿度土壤净氨化速率Fig.2 Soil net ammonification rates under different forest types and incubation conditions (temperature and moisture)
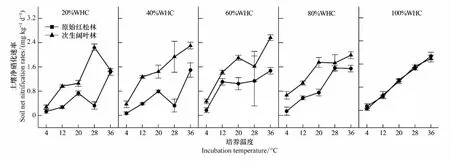
图3 不同林型、不同培养温度和培养湿度土壤净硝化速率变化Fig.3 Soil net nitrification rates under different forest types and cultivation conditions (temperature and moisture)
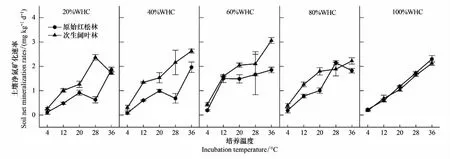
图4 不同林型、不同培养温度和培养湿度土壤净矿化速率变化Fig.4 Soil net mineralization rates under different forest types and incubation conditions (temperature and moisture)WHC:饱和持水量(saturated water holding capacity)
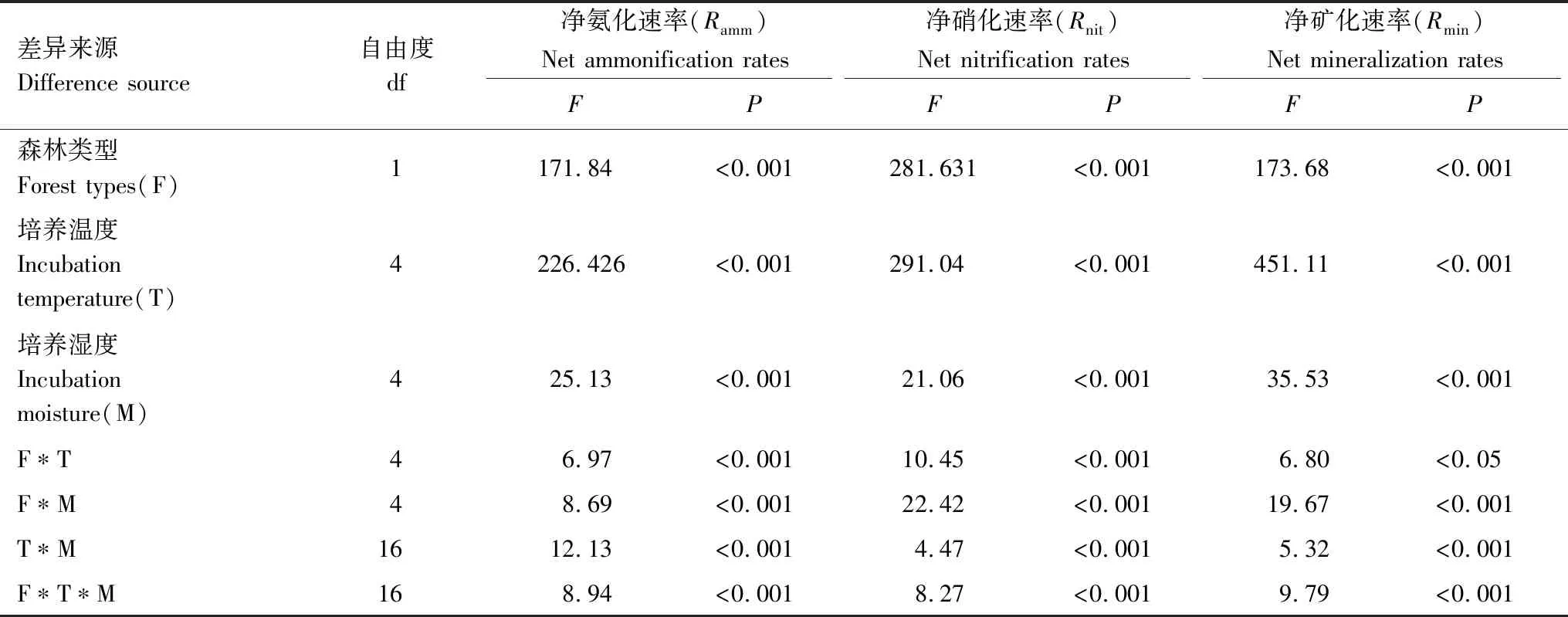
差异来源Difference source自由度df净氨化速率(Ramm)Net ammonification rates净硝化速率(Rnit)Net nitrification rates净矿化速率(Rmin)Net mineralization ratesFPFPFP森林类型Forest types(F)1171.84<0.001281.631<0.001173.68<0.001培养温度Incubation temperature(T)4226.426<0.001291.04<0.001451.11<0.001培养湿度Incubation moisture(M)425.13<0.00121.06<0.00135.53<0.001F∗T46.97<0.00110.45<0.0016.80<0.05F∗M48.69<0.00122.42<0.00119.67<0.001T∗M1612.13<0.0014.47<0.0015.32<0.001F∗T∗M168.94<0.0018.27<0.0019.79<0.001
2.3 土壤净氮矿化速率对温度和湿度变化的响应
在不同培养湿度下,原始红松林与次生阔叶林净矿化速率对温度变化的响应趋势存在一定差异,但两者的净矿化速率多随培养温度升高而升高,且在28—36℃之间达到最大值。在不同培养温度下,原始红松林与次生阔叶林净矿化速率对湿度变化的响应趋势则基本相似,土壤净矿化速率均随着培养湿度升高而升高,多在培养湿度60%WHC时达到高点后出现下降(图4)。
2.4 退化演替后土壤净氮矿化速率温度敏感性Q10及与土壤理化的相关性
利用指数模型Rmin=AeBT,计算了5个培养湿度下原始红松林和次生阔叶林的土壤净氮矿化速率的温度敏感性Q10,其均值分别为2.08和1.80,且两者差异显著。指数模型参数A 可以作为衡量基质质量的指标,被称为基质质量指数[19],原始红松林的A值也显著低于次生阔叶林(表4)。

表4 土壤净氮矿化速率指数模型参数(n=30)
n为样本数,表中数据为平均值±标准差,A为基质质量指数,B为温度反应系数,R2为指数方程拟合优度,Q10土壤氮矿化温度敏感性指数;同列不同小写字母表示差异显著(P<0.05)
土壤净氮矿化速率温度敏感性Q10在很大程度上受控于底物数量和质量,即土壤基质质量,而土壤的各项理化指标均能够直接或间接影响土壤基质质量[20]。Pearson′s相关性分析表明,Q10与所测的各项土壤理化指标(除铵态氮外)均呈负相关,其中与土壤有机质的相关性达到极显著水平(图5)。
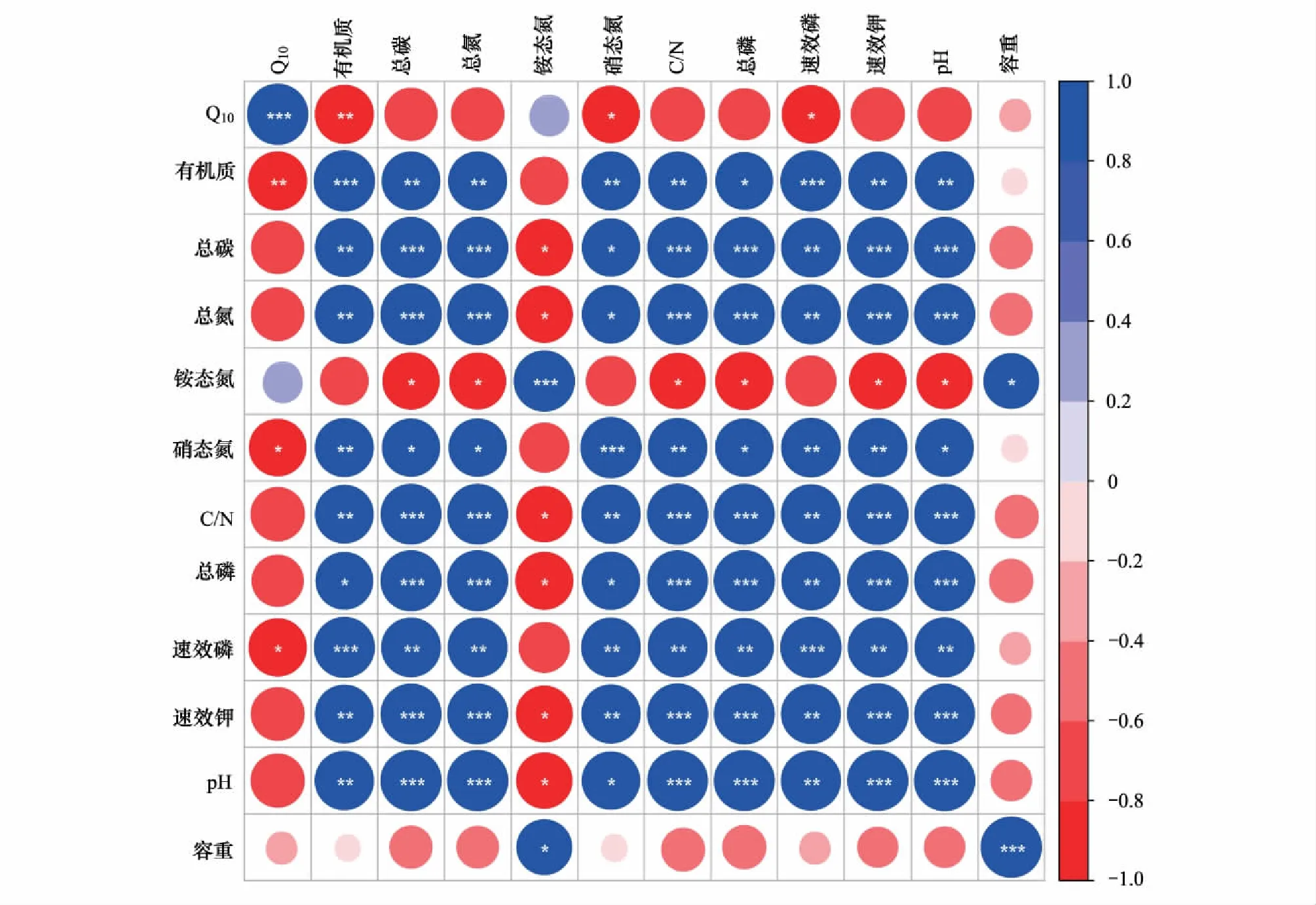
图5 土壤净氮矿化速率温度敏感性Q10与土壤理化相关性Fig.5 The correlation between soil physical and soil nitrogen mineralization Q10 value
3 讨论
3.1 退化演替后土壤净氮矿化速率变化
氮矿化作用是土壤有效氮库的主要来源,净氮矿化速率是衡量氮有效性的重要指标[21]。分析了不同温度及湿度下的所获的净矿化速率,次生阔叶林显著高于原始红松林,说明原始红松林退化演替为中幼龄次生林时,土壤对植物氮素的供应能力是升高的,这将会有助于中幼龄次生林地上植物生长,较高的净初级生产力水平能促进其植被恢复。事实上,森林不同演替阶段的净初级生产力的差异与林龄密切相关的,多数学者在研究森林净初级生产力与林龄关系时发现,森林净初级生产力通常在中幼林龄达到最大值后开始下降[22-23];黄兴召等在对杉木林的研究中得出了不同的结论,认为森林净初级生产与林龄呈显著负相关[24],但都一致认为顶级群落的初级生产力水平是偏低的。此外,原始红松林的针叶凋落物分解后会向中土壤中释放易挥发的单萜类物质,可能对土壤微生物氮矿化作用产生抑制[25]。
已经得到共识的是,土壤氮素转化是微生物主导的生物化学过程,因此,不同森林类型土壤净氮素矿化速率存在差异性主要原因是土壤微生物群落结构的异质性[26]。事实上,本课题组成员此前发现,原始红松林和次生阔叶林土壤微生物群落结构及功能均存在显著的差异[27]。
3.2 退化演替后土壤净氨化速率和净硝化速率变化
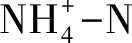
3.3 土壤净氮矿化速率温度敏感性
Liu等[33]通过Meta分析发现,中国森林生态系统的净氮矿化Q10值平均为1.94,原始红松林和次生阔叶林Q10值为2.08和1.80,分别高于和低于平均水平。原始红松林Q10值显著高于次生阔叶林,而基质质量(A)显著低于后者,验证了土壤的基质质量越差,氮矿化温度敏感性就越高,Q10值与基质质量指数呈负相关的结论[34]。土壤氮矿化是一个耗能的过程,主要通过微生物介导的酶促反应来实现,基质越差,土壤矿化时所需的酶促反应步骤越多,需要的活化能也越高,对温度升高的响应就更加剧烈[33]。研究中发现,土壤总有机质与Q10呈极显著负相关,说明土壤总有机质是底物水平中影响Q10的最主要的因素。土壤有机质不仅是土壤酶促反应底物的主要供源,而且还可以作为土壤微生物、酶和矿物质的有机载体,作为土壤固相中最复杂的系统,是土壤肥力的主要物质基础[35]。事实上,人们早就发现土壤总有机质含量是影响N循环的的最主要因素之一[36-37]。
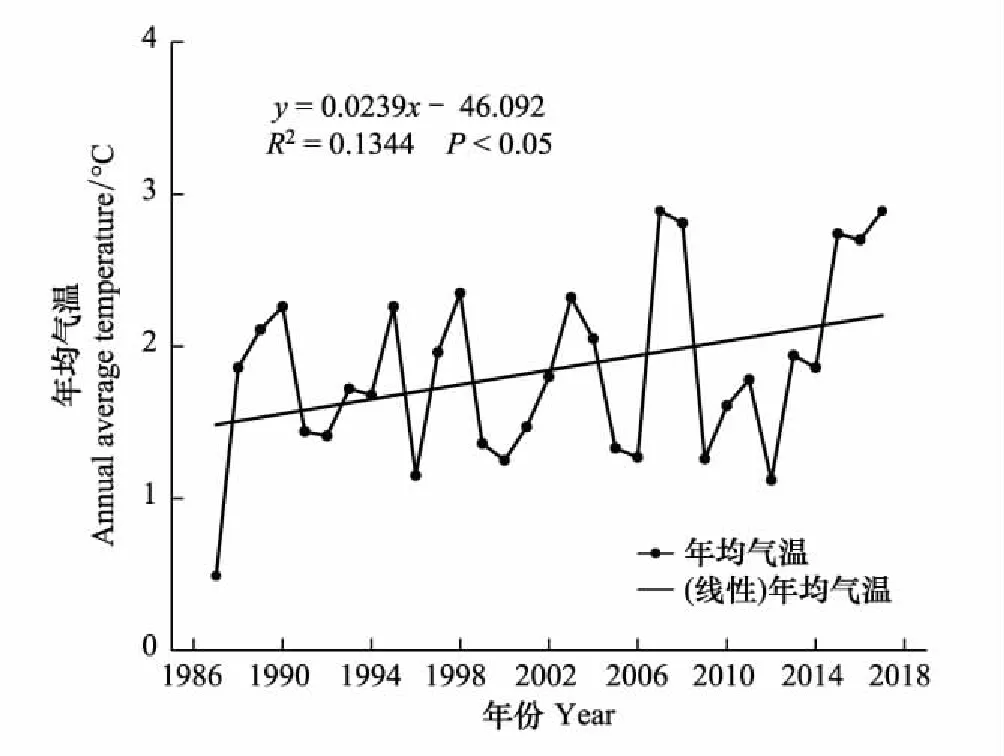
图6 带岭区近30年年平均气温变化 Fig.6 Changes of annual average temperature in Dailing Region in recent 30 years
Q10不仅可以表征不同基质土壤的温度敏感性,还是衡量土壤氮素矿化对未来温度变化响应的一个重要参数[38]。IPCC(2014)指出到21世纪末,全球气温将至少上升1.5℃,平均以0.15—0.3℃/10a的速度上升。通过对伊春市气象局获得的近30年的年平均气温的数据分析发现,此地区气温也呈现出相同的变暖趋势,年均气温以0.24℃/10a(P<0.05)的速率上升,和全球的平均升高速率相比,处于较高水平(图5)。显然,在这种全球气候变暖的大背景下,如果温度继续升高,和次生林相比,原始红松林土壤氮矿化速率的升高将会更为剧烈。已有研究表明,包括原始红松林在内的温带森林生态系统普遍具有氮素周转较慢,土壤氮素相对匮乏的现象[39-40],因此,推测未来气温的升高在短期内能有效缓解红松林的氮限制,促进其初级生产力的提高,但从长远来看,可能会造成红松林土壤氮素的加速流失,最终降低土壤可利用性氮素量,从而对植物生长和生态系统结构和功能形成负反馈[41]。
4 论
(1)原始红松林遭到皆伐退化演替为中幼龄次生阔叶林,其净氮矿化速率显著升高,因此,对植物氮素的供应能力是显著增强的。
(2)培养温度和湿度及两者的交互作用均对土壤氮转化速率影响显著,原始红松林和次生阔叶林净矿化速率最适温度和湿度是28℃—36℃和60%饱和持水力。
(3)原始红松林Q10显著高于次生阔叶林,Q10值与基质质量指数(A)呈负相关,与土壤有机质呈极显著负相关。
