生态恢复先锋植物苔藓的耐旱性评价及应用
2019-06-12高旭
高旭
(国营新宾满族自治县赵家林场,辽宁 抚顺 113213)
苔藓植物是植物界中由水生向陆生过渡的一大类群,其种类仅次于种子植物的高等植物群落[1], 有其特殊的适应机制,能在寒冷、干旱、温度过高和阴暗的环境中生长繁育,由于其特殊的生理适应性,其在森林生态系统、岩石和沙丘、草甸高山等生态环境中扮演着重要的生态恢复先锋植物角色[2]。本项试验以抚顺博大复垦基地矸石山排土场阴坡边坡地为研究对象,兼顾前期研究成果,选取4种植物组配置模式进行生态修复试验,综合分析土壤改良及植被恢复效果,探索其各参试植物生长发育状况,以及土壤理化性质,以期为生态恢复技术模式提供理论参考和试验依据,最终筛选出最优配比模式[3,4]。
1 研究方法
1.1 试验材料
以辽宁地区常见苔藓植物为试验材料,试验伴生树种选择耐旱性较强的紫穗槐(AmorphafruticosaLinn.),草本植物选择草木樨(MeliotusofficinalisL.)。苔藓种类及来源具体见表1。
1.2 试验设计
试验地选取抚顺博大复垦基地矸石山排土场阴坡边坡地。选择紫穗槐为造林优选树种,以真藓配子体和草木樨种子混合物为优选草本植物。共设计4种配置模式:模式一(M1),紫穗槐行间直播真藓配子体和草木樨种子混合物(真藓和草木樨质量比为0.5∶1);模式二(M2),紫穗槐树间隔栽植—行间直播草木樨;模式三(M3),紫穗槐行间隔栽植—行间直播真藓配子体和紫穗槐种子混合物(真藓和草木樨质量比为0.5∶1);模式四(M4),紫穗槐间隔栽植—行间直播紫穗槐。2017年5月中旬进行造林试验,每种模式以紫穗槐各50株为一个造林地块,株行距0.5 m×0.5 m,每种模式种植5个地块,4种模式共计20个地块。紫穗槐栽植均采用树穴内放客土15 cm造林措施。因边坡地位于各排土场平台之间,坡度较大,表层土壤、砂石松散易滑落,因此栽植前不进行整地,以尽可能保护边坡土壤本底状况,真藓以具有配子体的残体喷播施用,喷播量为500 g·m-2。

表1 苔藓种类及来源
1.3 测定项目
2017年依据4种模式进行人工造林,分别于造林完成时(A1),2017年8月(A2)和2018年8月(A3)进行株高、冠幅、成活率、地径、生物量和地表覆盖率等状况调查。
2 结果与分析
2.1 干旱胁迫下抗旱性主成分分析
由于各品种之间存在固定的差异性,为了减小其之间的误差,本试验所测定的各项数据指标都进行了人为抗旱干扰试验测定,对其各结果进行了主成分分析法作抗旱分析主要根据。由于主成分分析法相对于方差分析和绝对值法更能准确反映各个苔藓抗旱指标,本方法采用的是降维思想,直接反映了无法直观测量的隐性变量数据指标,对比其他方法更能准确地分析了各个苔藓品种间抗旱性指标数据的准确,综合性全方位的概括了多因素分析抗旱指标的有效途径。在本次抗旱指标的测定实验中,不同年度测定的抗旱数据经过加权平均分析后在利用主成分分析,所得的抗旱指标结果如表2所示。
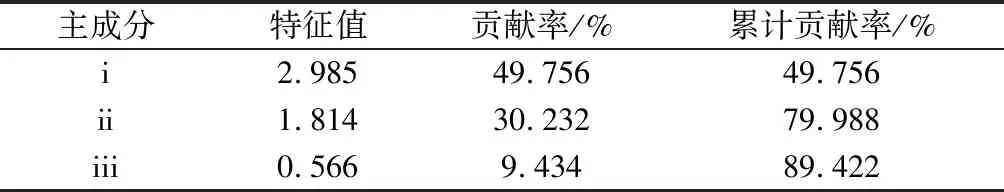
表2 2个主成分特征值及累计贡献率
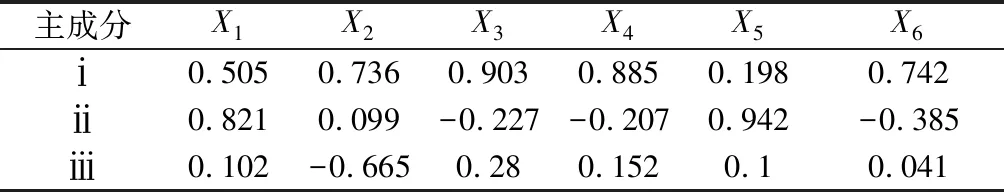
表3 各因子载荷矩阵

表4 各成分向量矩阵
各个苔藓抗旱指标经过主成分分析,其中各个抗旱指标数据中单独成立的6个抗旱主成分贡献率已接近其主体贡献率的100%,X1~X6之间数据可以完全代表了各个指标间大量数据信息,为其方便计算及形象直观展示,使用了排列前3的数据指标变异进行主成分分析法对其分析描述,依据其主成分分析数据的因子载荷量制作主成分散点图,更能形象直观展现了抗旱指标差异,通过图1可知,各个苔藓抗旱指标数据主成分分析载荷值所呈现的不同空间分布,不同测定指标均不落在原点和坐标轴上,有效的阐明了各个指标抗旱性和坐标轴所对应的因子都有相关性,并且相互作用非常明显。
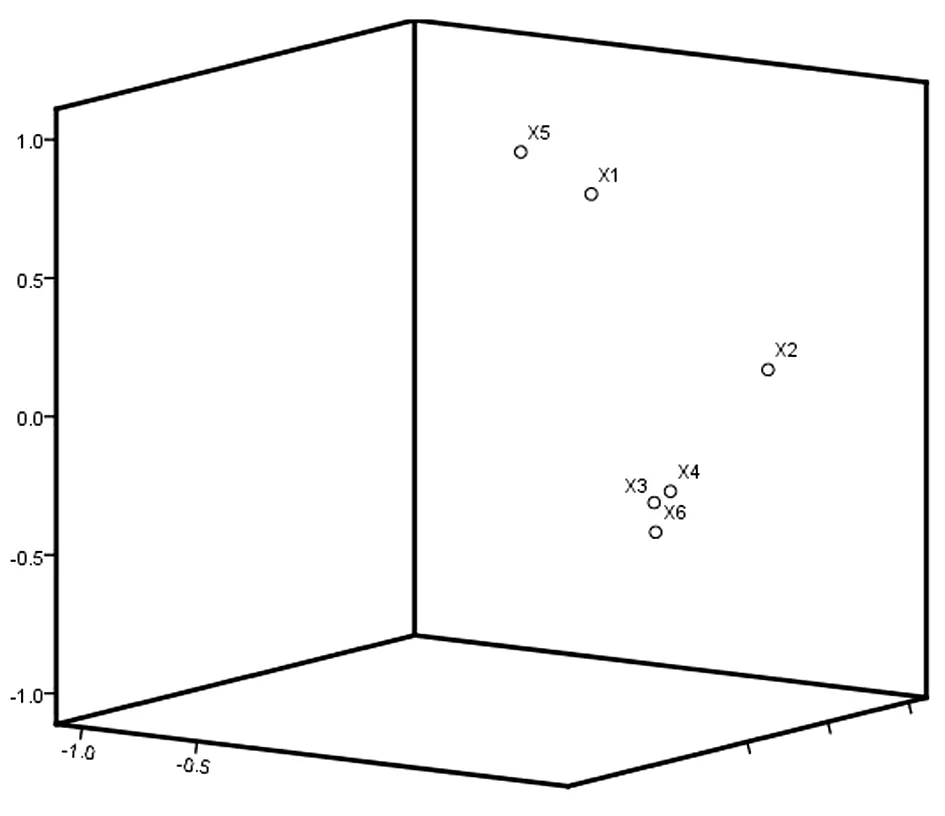
图1 干旱胁迫对苔藓抗旱性影响的主成分散点图
表3和表4分别为各成分的因子载荷矩阵和成分向量矩阵,据此可获得各因子得分公式,如下:
Y1=0.29×X1+0.43×X2+0.52×X3+0.51×X4+0.11×X5+0.43×X6
(1)
Y2=0.61×X1+0.07×X2-0.17×X3-0.15×X4+0.7×X5-0.29×X6
(2)
Y3=0.14×X1-0.88×X2+0.37×X3+0.2×X4+0.13×X5+0.05×X6
(3)
根据各品种Y1、Y2和Y3的值,各品种的综合得分公式Y为:
Y=0.476 74×Y1+0.276 25×Y2+0.124 57×Y3
(4)
通过矩阵分析后Y值可见,Y值随着指标的增大,其各个苔藓品种的抗旱性则越来越强。由此可知,在矸石山干旱地区的植被恢复中,各个苔藓品种之间的抗旱性依次顺序为:葫芦藓>泥炭藓>真藓>小墙藓>大灰藓>山地墙藓>仙鹤藓>狭叶小羽藓>刺叶墙藓>黑扭口藓>提灯藓>大帽藓>曲柄藓>小叶藓>凤尾藓。
2.2 基于SOM神经网络干旱胁迫苔藓抗旱性聚类分析
对15个苔藓种类在4个胁迫时间下测得的6个性状的相对值进行数据标准化后,利用数据挖掘软件MATLAB 2012a进行自组织映射(SOM)聚类分析,将标准化的数据作为网络的输入层,进行200 000次的训练后稳定,得到聚类分析结果如表5所示。
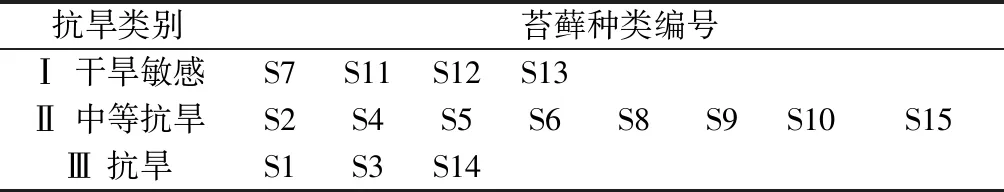
表5 基于SOM神经网络聚类分析结果
通过本试验对各个苔藓抗旱指标数据进行分析处理后,我们选定的各个品种大致可以分为3种类型,依据主成分分析法对其数据分析可知,从第Ⅰ类到第Ⅲ类基本遵循抗旱性由弱到强排列。其中,真藓(S1)、葫芦藓(S3)和山地墙藓(S14)对干旱胁迫不敏感,抗旱能力较强,可用于排土场生态修复中。
2.3 不同模式下参试灌木和草种生长发育影响
不同模式下草木樨和紫穗槐的生长状况可知,除边坡地下部紫穗槐外,真藓配子体可显著提高先锋作物的保存率。混有真藓配子体的草木樨保存率达到了96.4%、89.6%和84.3%,而未直接播种的草木樨种子的最高发芽率仅为在边坡地上部的80.2%。真藓配子体对紫穗槐影响较大,具有促进萌发、提高保存率的作用,但是在边坡地下部,是否混有真藓配子体的紫穗槐保存率间差异不显著,这可能是由于边坡地不稳定,下部植物易受中上部坍塌或者滚石的影响,M4模式下的一部分紫穗槐可能因砾石花落或坍塌等而出现非正常死亡。真藓配子体提高种子发芽率的原因可能是苔藓植物具有保水作用,在干旱条件下,降低了土壤水分的散失速度,或者是因为真藓配子体中的代谢产物对种子萌发有促进作用,这仍需进一步研究。
不同模式下,草木樨的单株生物产量差异不大。M1模式的草木樨单株生物量上部、中部和下部分别为20.9 g、22.3 g和21.5 g,M2模式的单株生物量上部、中部和下部19.8 g、22.4 g和21.6 g,不同模式间差异不显著,且上部、中部和下部边坡区域间草木樨单株生物量差异不显著。紫穗槐的单株生物量与草木樨具有类似的特点,不同边坡区域和不同模式间差异较小。
M2模式下,不同边坡区域内草木樨的群体生物产量均高于M1模式。M2模式下,边坡上部区域的草木樨群生物量为152.49 g,是M1模式的111.3%,增加了11.3%;下部区域,M1模式的草木樨群体生物量最低,仅有108.4 g;但相同模式内,中部区域与下部区域间群体生物量差异不显著。M3和M4模式下,紫穗槐群体生物量间差异明显,M4模式下,上部区域的紫穗槐生物量为195.4 g,高于M3模式9.5%,中部和下部区域紫穗槐生物产量分别高于M3模式18.6和16.5%。

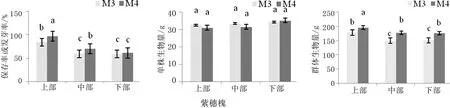
图2 不同模式下紫穗槐和草木樨生长状况
3 结论
葫芦藓、泥炭藓、真藓3种苔藓可以通过提高周边土壤的水分含量,有效促进了灌木、草本萌发生长,提高了单株株高和生物产量等,但对群体生物产量影响较小;利用主成分分析各个苔藓抗旱指标,结果表明苔藓种类抗旱性由大到小排列为:葫芦藓>泥炭藓>真藓>小墙藓>大灰藓>山地墙藓>仙鹤藓>狭叶小羽藓>刺叶墙藓>黑扭口藓>提灯藓>大帽藓>曲柄藓>小叶藓>凤尾藓,即葫芦藓、泥炭藓、真藓等3种苔藓可作为排土场生态恢复的先锋植物。
