转聚合cry1A.105、cry2Ab2和cp4epsps基因抗虫耐除草剂玉米的田间抗性评价
2019-06-11李国平刘冰黄建荣邱峰封洪强
李国平 刘冰 黄建荣 邱峰 封洪强
摘要为明确转聚合cry1A.105、cry2Ab2和cp4epsps基因的抗虫耐除草剂玉米的抗虫性和对除草剂的耐受性,本文通过田间人工接虫,以食叶级别、存活幼虫数、蛀孔数、隧道长度和雌穗被害级别等分别作为评价玉米心叶期和穗期的抗虫性标准,评价了转聚合cry1A.105、cry2Ab2和cp4epsps基因,具有抗虫耐除草剂复合性状的玉米品种‘ND6708YGRR和‘ND6515YGRR及同型非转基因‘ND6708、‘ND6515和当地‘郑单958品种对亚洲玉米螟、黏虫和棉铃虫的抗虫效果;通过喷施除草剂后调查玉米植株的存活率,评价上述品种对除草剂的耐受性。田间试验结果表明:‘ND6708YGRR和‘ND6515YGRR两个转基因玉米品种对亚洲玉米螟、黏虫及棉铃虫的抗性水平均为高抗,且在4叶期和8叶期喷施草甘膦后其受害率均为0,而同型对照品种和当地对照品种喷施同剂量草甘膦后受害率为100%。综上,聚合cry1A.105、cry2Ab2和cp4epsps基因,具有抗虫耐除草剂复合性状的玉米品种‘ND6708YGRR和‘ND6515YGRR对亚洲玉米螟、黏虫和棉铃虫具有较强的抗虫效果,对草甘膦具有较好的耐受性。此研究可为将来建立以转Bt基因玉米为主要手段的综合防治技术体系提供依据。
关键词亚洲玉米螟;黏虫;棉铃虫;转基因玉米;抗性
中图分类号:S 435.13, S 513
文献标识码:A
DOI:10.16688/j.zwbh.2018151
世界上第一個转苏云金杆菌Bacillus thuringiensis(简称Bt)cry1Ab基因的玉米于1996年在美国开始商业化种植,有效地控制了靶标害虫欧洲玉米螟Ostrinia nubilalis (Hübner)的为害[1]。到2016年,美国的转基因玉米种植面积达到3 500万hm2,占玉米总种植面积的92%,其中抗虫玉米占3%,耐除草剂玉米占13%,具有抗虫和耐除草剂复合性状的玉米占76%[2]。随着Bt玉米的大面积种植,美国的欧洲玉米螟种群得到了有效控制,其不仅在Bt作物上的发生明显减轻,且同一种植区内其他非Bt寄主作物上种群数量也显著降低[34]。在现有的玉米转化体构建中,cry1Ab和cry1F通常用于单基因和多基因的表达,而vip3A, cry2Ab2及cry1A.105只用于多基因的表达[5]。由于种植表达两个基因的转基因作物比种植表达单基因作物更能够有效地延缓抗性的发生[6], 种植转多个基因作物在美国呈流行的趋势。到目前为止,除美国外,巴西、阿根廷、加拿大、南非、乌干达、西班牙等17个国家批准了商业化种植转基因玉米,我国尚未商业化种植[2]。
亚洲玉米螟Ostrinia furnacalis (Guenée)、黏虫Mythimna separata (Walker) 和棉铃虫Helicoverpa armigera (Hübner)是我国为害玉米的3种重要的鳞翅目害虫[78]。亚洲玉米螟是玉米常年发生的第一大类害虫,一般年份可造成春玉米减产10%左右,夏玉米减产20%~30%,大发生年份可致减产达50%以上[9]。黏虫是一种暴食性害虫,可以在小麦、玉米、水稻和其他作物上发生为害,暴发为害时植株叶片被大量取食,仅剩叶脉。2012年以来,黏虫已经对河北、河南、陕西、辽宁以及内蒙古等地区的作物,尤其是玉米造成了严重的危害[8,1011]。棉铃虫是棉花上的重要害虫,自20世纪80年代后期以来, 棉铃虫已成为我国夏玉米产区及辽宁南部春玉米穗期的主要害虫, 该虫主要取食玉米花丝、穗尖及籽粒,一般可造成减产5%~7%, 严重者减产10%以上,且常诱发玉米穗腐病的发生,影响品质[12]。
Bt玉米对靶标害虫欧洲玉米螟有很好的控制效果, 但对其他鳞翅目害虫的控制效果因所转基因的不同而异。比如,西班牙种植的转cry1Ab基因玉米对地中海蛀茎夜蛾Sesamia nonagrioides(Lefebvre)的控制效果在99%以上[13],而当地的一种黏虫Mythimna unipuncta在该Bt玉米上的存活率却高达80%以上[1415]。在我国,转cry1Ab基因玉米对亚洲玉米螟、黏虫和棉铃虫都具有较好的田间抗虫效果[9,1619]。目前有关转聚合cry1A.105和cry2Ab2基因玉米对亚洲玉米螟、黏虫和棉铃虫田间抗虫效果还未见报道,本文通过田间人工接虫试验研究了转聚合cry1A.105、cry2Ab2和cp4epsps玉米对这3种鳞翅目害虫的抗虫效果以及对草甘膦的耐受性,为将来建立以Bt玉米为主要手段的综合防治技术体系提供依据。
1材料与方法
1.1供试玉米品种
供试的转聚合cry1A.105、cry2Ab2和cp4epsps基因抗虫抗除草剂复合性状玉米品种‘ND6708YGRR和‘ND6515YGRR及同型非转基因玉米品种‘ND6708、‘ND6515均由孟山都公司提供,以‘郑单958作为当地对照品种。
1.2供试虫源
试验用亚洲玉米螟卵块由中国农业科学院植物保护研究所提供;黏虫卵由河北省农林科学院植物保护研究所提供;棉铃虫卵为室内用人工饲料饲养的采自河南新乡七里营的种群。
1.3试验小区设计
试验于2009年7月-10月在河南省新乡市七里营进行,试验地周围有500 m以上非玉米作物隔离带。抗虫性鉴定采用完全随机区组设计、3次重复,每个小区面积为30 m2(5 m×6 m),行距60 cm,株距25 cm,8行区。不同小区间设有2 m宽的隔离带,以避免害虫在不同小区间扩散。除必要的农事操作外,生长期不施用任何杀虫剂。
除草剂试验采用裂区试验设计,主区为两个处理,即喷除草剂与不喷除草剂,副区为随机区组设计,5个品种,重复3次,小区面积为12 m2(5 m×2.4 m),小区行长5 m,行距0.60 m,4行区。不同小区间设有2 m宽的隔离带。施用的除草剂为孟山都公司生产的41%草甘膦异丙胺盐(glyphosate isopropylamine salt)水剂,采用马来西亚进口恒压稳流喷雾器进行喷施。
1.4对亚洲玉米螟抗虫性鉴定
分别在玉米心叶期(玉米植株6叶至8叶)和吐丝期接种亚洲玉米螟卵块进行抗虫鉴定。在玉米心叶期,将亚洲玉米螟黑头卵接入玉米心叶丛中,在吐丝期将黑头卵接入雌穗新鲜花丝上。每株接2块卵(约60粒),并挂牌标记以便于调查记载。每小区接虫30株。心叶期第1 次接虫5~7 d 后,补接1 次。心叶期接虫后,分别于第14天和21天逐株调查玉米心叶食叶级别,采用9级标准[20]。吐丝期接虫后,在收获前逐株调查幼虫存活数、蛀孔数及隧道长度。
1.5对黏虫抗虫性鉴定
在玉米心叶期将初孵黏虫幼虫接入玉米心叶丛中,每小区接虫30株,每株接种初孵幼虫30头。接虫3 d后,进行第2次补接,数量同第1次。在接虫14 d后,采用农业部953号公告10.1-2007的9级标准[21], 调查玉米叶片食叶级别和幼虫存活数。
1.6对棉铃虫抗虫性鉴定
在玉米吐丝期将棉铃虫初孵幼虫接到玉米花丝上,每小区人工接虫30株,每株接种初孵幼虫20~30头,接虫3 d 后,第2次补接,接虫数量同第1次。人工接虫后第14天和第21天,采用农业部953号公告10.1-2007的标准[21],逐株调查雌穗受害级别、每个雌穗存活幼虫数和雌穗被害长度 (cm)。
1.7耐除草剂鉴定
分别在玉米4叶期和8叶期喷施41%草甘膦异丙胺盐水剂各1次,施用浓度为1.68 kg/hm2,用药后1周、2周和4周调查和记录药害症状(选取药害症状最轻的5株),药害症状分级按5级标准执行[22],计算受害率。
1.8數据统计与处理
用单因素方差分析不同品种间的差异,百分数进行反正弦转换,多重比较采用Duncans新复极差法,采用统计软件SPSS 20.0 进行,显著性检验水平均为P<0.05。
2结果与分析
2.1转基因玉米对亚洲玉米螟的田间抗性
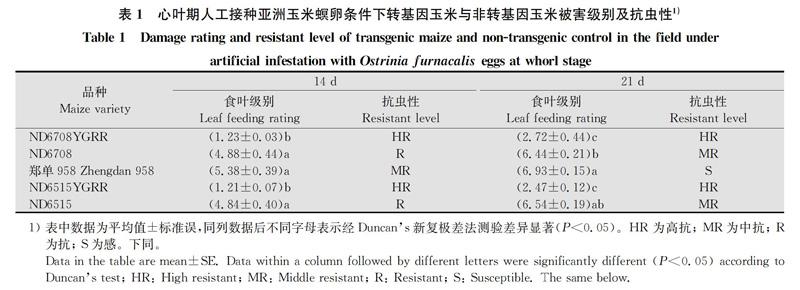
在玉米心叶期接种亚洲玉米螟卵块,14 d后,‘ND6708YGRR和‘ND6515YGRR 两个转基因品种食叶级别分别为1.23级和1.21级,达到高抗水平,而其同型非转基因对照品种‘ND6708和‘ND6515食叶级别分别为4.88级和4.84级,当地对照品种‘郑单958食叶级别为5.38级(表1)。品种间食叶级别差异显著(F4,14=12.768,P<0.000 1),‘ND6708YGRR和‘ND6515YGRR两个品种受害级别均小于其同型非转基因对照品种‘ND6708和‘ND6515及当地对照品种‘郑单958,且3个对照品种间差异不显著(表1)。
在玉米心叶期接种亚洲玉米螟卵块21 d后,‘ND6708YGRR和‘ND6515YGRR两个品种食叶级别为2.72级和2.47级,达到高抗水平,而其同型对照品种‘ND6708和‘ND6515的食叶级别则分别为6.44级和6.54级,‘郑单958为6.93级(表1)。品种间受害级别差异显著(F4,14 =244.852, P=0.000 1),‘ND6708YGRR和‘ND6515YGRR两个品种无显著差异,均轻于其同型对照品种‘ND6708和‘ND6515以及‘郑单958(表1)。
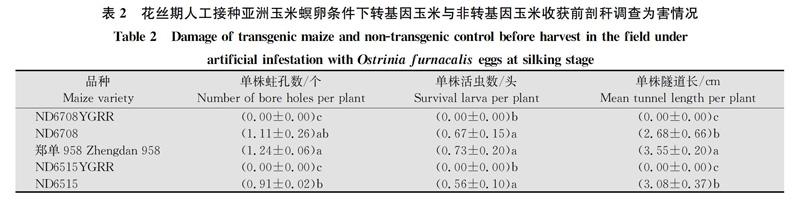
玉米收获前剖秆调查显示:‘ND6708YGRR和‘ND6515YGRR两个品种平均单株蛀孔数为0个,平均活虫数为0头,平均隧道长度为0 cm;与其相对应的非转基因对照品种‘ND6708、‘ND6515以及‘郑单958的平均单株蛀孔数分别为1.11、0.91和1.24个,平均单株活虫数分别为0.67、0.56和0.73头,平均单株隧道长度为2.68、3.08和3.55 cm(表2)。蛀孔数、活虫数、隧道长度在品种间差异显著(F4,14=242.266,P=0.000 1;F4,14=262.362,P=0.000 8;F4,14=218.245,P=0.000 1),转基因品种显著低于非转基因品种(表2)。表明‘ND6708YGRR和‘ND6515YGRR在玉米的营养生长和生殖生长阶段对亚洲玉米螟的抗性均显著高于其同型非转基因对照和当地对照品种。
2.2转基因玉米对黏虫的田间抗性
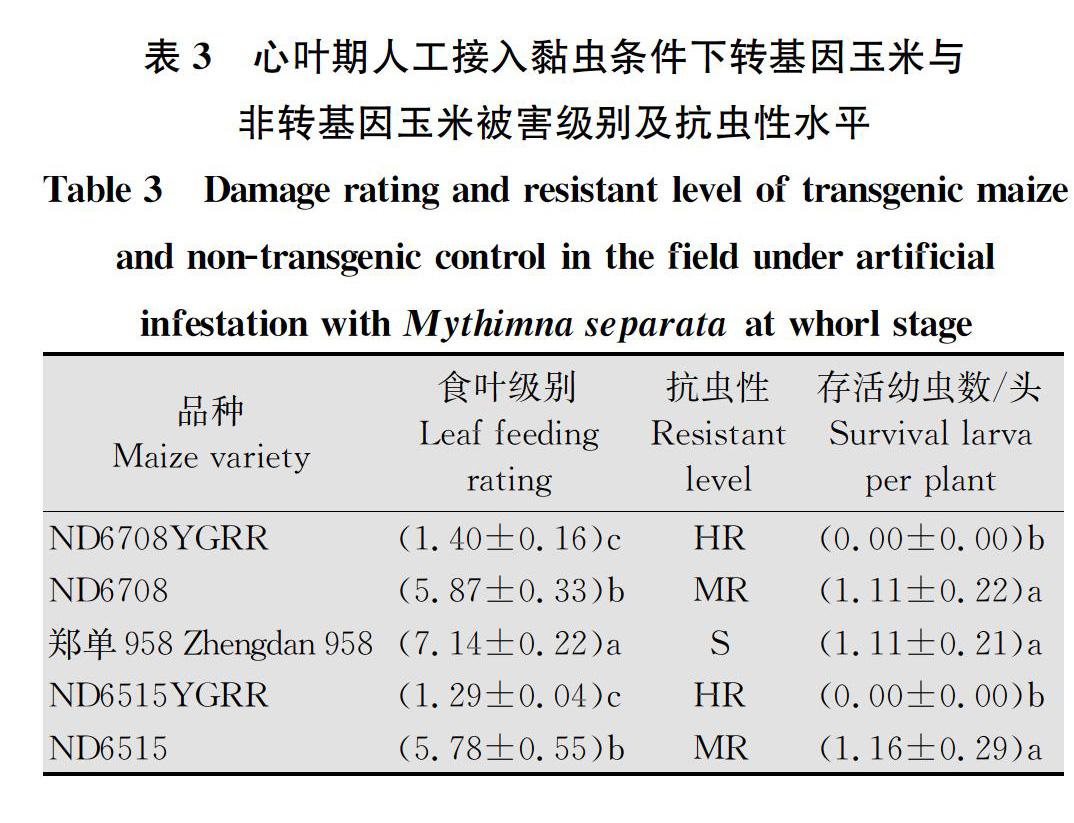
在玉米心叶期接入初孵黏虫,14 d后,‘ND6708YGRR和‘ND6515YGRR的叶片食叶级别分别为1.40级和1.29级,达到高抗水平;而其同型非转基因对照品种‘ND6708和‘ND6515的叶片食叶级别分别为5.87级和5.78级,‘郑单958食叶级别为7.14级(表3)。转基因玉米食叶级别显著低于其同型非转基因对照和‘郑单958品种(F4,14=32.294 0,P=0.000 1) (表3)。
‘ND6708YGRR和‘ND6515YGRR植株上的平均单株活虫数均为0头;而其同型非转基因对照品种‘ND6708和‘ND6515的平均单株活虫数分别为1.11头和1.16头,当地对照品种‘郑单958的平均单株活虫数为1.11头(表3)。转基因玉米上的平均存活数目显著低于其同型非转基因对照和当地对照品种(F4,14 =142.576, P =0.000 1)。
由以上分析可以得知,转基因抗虫抗除草剂复合性状玉米‘ND6708YGRR和‘ND6515YGRR对黏虫的抗性显著高于其同型非转基因对照和‘郑单958品种。
2.3转基因玉米对棉铃虫的田间抗性
在玉米吐丝期接入棉铃虫初孵幼虫,14 d后,‘ND6708YGRR和‘ND6515YGRR的雌穗受害级别分别为0.11级和0.04级,幼虫存活数均为0头,雌穗被害长度均为0 cm。而其同型非转基因对照品种‘ND6708和‘ND6515雌穗受害级别分别为0.61级和0.58级,幼虫存活数分别为0.17头和0.19头,雌穗被害长度分别为0.22 cm和0.24 cm;当地对照品种‘郑单958雌穗受害级别为0.64级,幼虫存活数为0.30头,雌穗被害长度为0.31 cm,转基因品种显著低于非转基因品种(雌穗受害级别:F4,14=110.569,P=0.000 1;存活幼虫数:F4,14=16.858,P=0.000 1;雌穗被害长度:F4,14=34.034,P=0.000 1)(表4)。
21 d后,‘ND6708YGRR和‘ND6515YGRR的雌穗受害级别分别为0.11级和0.08级,幼虫存活数均为0头,雌穗被害长度均为0 cm。而其同型非转基因对照品种‘ND6708和‘ND6515雌穗受害级别分别为0.64级和0.67级,幼虫存活数分别为0.22头和0.16头,雌穗被害长度分别为0.29 cm和0.25 cm;当地对照品种‘郑单958雌穗受害级别为0.62级,幼虫存活数为0.13头,雌穗被害长度为0.28 cm。转基因品种显著低于非转基因品种(雌穗被害率:F4,14=34.136,P=0.000 1;存活幼虫数:F4,14=14.144,P=0.000 1;雌穗被害长度:F4,14=20.420,P=0.000 1)(表4)。
2.4转基因玉米对草甘膦的耐受性
在4叶期喷施草甘膦1周后,非转基因对照品种显示出了明显的受害情况。其主要症状为玉米植株停止生长,叶片发黄枯萎,植株死亡。而抗虫耐除草剂复合性状玉米品种‘ND6708YGRR和‘ND6515YGRR则无此现象发生。2周后,非转基因对照品种受害率达到100%,植株全部死亡;而抗虫耐除草剂复合性状玉米品种‘ND6708YGRR和‘ND6515YGRR的植株受害率为0(表5)。
在8叶期喷施草甘膦2周和4周后,抗虫耐除草剂复合性状玉米品种‘ND6708YGRR和‘ND6515YGRR的植株受害率均为0,非转基因对照品种受害率均达到100%。
3结论与讨论
轉Bt基因玉米通过在生长季内持续表达杀虫蛋白,害虫的初孵幼虫通过取食其组织而被杀死在低龄状态,从而达到田间控制作用[23]。因此,评价转Bt基因玉米的田间抗虫效果通常是采用其初孵幼虫进行抗性鉴定。本文通过人工接虫,对聚合cry1A.105、cry2Ab2和cp4epsps基因抗虫耐除草剂玉米的抗虫性和对除草剂的耐受性进行了田间抗性评价,结果表明‘ND6708YGRR和‘ND6515YGRR两个转基因品种对河南新乡玉米常发钻蛀性害虫玉米螟具有很好的抗虫效果,对穗期棉铃虫的抗性也较高,对偶发性害虫黏虫也表现出了极高的抗性,同时‘ND6708YGRR和‘ND6515YGRR对草甘膦也表现出了很好的耐受性。表现出与转cry1A.105+cry2Ab2基因的MON89034相似的抗虫效果[24]。
玉米在整个生长季常常遭受多种害虫的为害,因此评价转Bt基因玉米对多种害虫的田间抗性效果, 能够更全面地了解玉米转化体的抗性谱,对于其适生区的种植具有很好的田间指导作用。在我国,亚洲玉米螟是玉米上的第一大害虫,在全国玉米产区广泛分布,而棉铃虫在黄淮海棉区、长江棉区等常年为害玉米[12]。黏虫是一种典型的迁飞性偶发害虫,在特殊年份会大发生[11]。因此,应根据当地种植作物类型及其害虫种类,在同一地区合理使用抗虫作物。种植表达Cry1A.105和Cry2Ab2蛋白的转基因玉米与我国目前已经商业化种植的转Bt基因棉花不同,Bt棉大多表达单一的Cry1Ac蛋白,而Cry1A.105和Cry2Ab2蛋白与其杀虫机制不同,由于作用位点的增多,害虫同时对两种杀虫基因产生抗性的几率比单个基因低,可以延缓抗性的发展。因此,即使在黄淮海地区种植此类型的转基因品种,其引起棉铃虫对Bt棉产生交互抗性的风险也相对较小。
本文仅评价了转聚合cry1A.105、cry2Ab2和cp4epsps基因玉米对亚洲玉米螟、棉铃虫和黏虫的抗性。但随着农业产业结构的调整、玉米耕作模式的改变以及气候变化等多种因素的影响,为害玉米的害虫种类也在不断发生变化。目前,桃蛀螟Conogethes punctiferalis (Guenée)在玉米上为害逐年加重[25],尤其在黄淮海夏玉米区,严重时在玉米果穗上的种群数量和为害程度甚至超过亚洲玉米螟,成为玉米穗期的主要害虫[26]。在今后进行转基因抗虫玉米的田间抗性鉴定时,需要进一步研究其对桃蛀螟的抗性。
参考文献
[1]MASON C E, RICE M E, DIFONZO C, et al. European corn borerecology and management and association with other corn pests [M]. US: Iowa State University,2018.
[2]JAMES C. Global status of commercialized biotech/GM crops[R]. ISAAA Briefs: Brief 52, Ithaca, NY, 2016.
[3]HUTCHISON W D, BURKNESS E C, MITCHELL P D, et al. Areawide suppression of European corn borer with Bt maize reaps savings to nonBt maize growers [J]. Science, 2010, 330(6001): 222225.
[4]DIVELY G P, VENUGOPAL P D, BEAN D, et al. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers [J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(13):33203325.
[5]USEPA (United States Environmental Protection Agency), Current and previously section 3 PIP registrations [R]. http:∥www. epa. gov/pesticides/biopesticides/pips/piplist. htm, 2009.
[6]ZHAO Jianzhou, CAO Jun, COLLINS H L, et al. Concurrent use of transgenic plants expressing a single and two Bacillus thuringiensis genes speeds insect adaptation to pyramided plants [J]. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102: 84268430.
[7]王振營,鲁新,何康来,等.我国研究亚洲玉米螟历史、现状与展望[J].沈阳农业大学学报,2000,31(5):402412.
[8]江幸福,张蕾,程云霞,等.我国粘虫发生危害新特点及趋势分析[J].应用昆虫学报,2014,51(4):14441449.
[9]中国农业科学院植物保护研究所,中国植物保护学会.中国农作物病虫害上册[M].北京:中国农业出版社,2014:673682.
[10]曾娟,姜玉英,刘杰.2012年黏虫暴发特点分析与监测预警建议[J].植物保护,2013,39(2):117121.
[11]程登发,赵中华.我国部分地区粘虫暴发原因分析与对策建议[J].种子科技,2016(10):8990.
[12]王振营,何康来,文丽萍,等.第四代棉铃虫卵在华北夏玉米田的时空分布[J].中国农业科学,2001,34(2):153156.
[13]FAROMS G P, ANDREADIS S S, DE LA POZA M, et al. Comparative assessment of the fieldsusceptibility of Sesamia nonagrioides to the Cry1Ab toxin in areas with different adoption rates of Bt maize and in Btfree areas [J]. Crop Protection, 2011, 30: 902906.
[14]PREZHEDO M, REITER D, LPEZ C, et al. Processing of the maize Bt toxin in the gut of Mythimna unipuncta caterpillars [J]. Entomologia Experimentalis et Applicata, 2013, 148: 5664.
[15]GONZLEZCABRERA J, GARCA M, HERNNDEZCRESPO P, et al. Resistance to Bt maize in Mythimna unipuncta (Lepidoptera: Noctuidae) is mediated by alteration in Cry1Ab protein activation [J]. Insect Biochemistry and Molecular Biology, 2013, 43: 635643.
[16]HE Kanglai, WANG Zhenying, ZHOU Dayong, et al. Evaluation of transgenic Bt corn for resistance to the Asian corn borer (Lepidoptera: Pyralidae)[J]. Journal of Economic Entomology, 2003, 96(3):935940.
[17]王振营,王冬妍,何康来,等.转Bt基因玉米对粘虫的室内杀虫效果评价[J].植物保护学报,2005,32(2):153157.
[18]常雪艳,何康来,王振营,等.转Bt基因玉米对棉铃虫的抗性评价[J].植物保护学报,2006,33(4):374378.
[19]常雪,常雪艳,何康来,等.转cry1Ab基因玉米对粘虫的抗性评价[J].植物保护学报,2007,34(3):225228.
[20]中华人民共和国农业行业标准.NT/T 1248.52006:玉米抗病虫性鉴定技术规范第5部分:玉米抗玉米螟鉴定技术规范[S].北京:中国农业出版社,2006.
[21]中华人民共和国农业部.农业部953号公告-10.1-2007:转基因植物及其产品环境安全检测:抗虫玉米第1部分:抗虫性[S].北京:中国农业出版社,2007.
[22]中华人民共和国国家标准.GB/T 17980.422000:田间药效试验准则(一)除草剂防治玉米地杂草[S].北京:中国标准出版社,2005.
[23]KOZIEL M G, BELAND G L, BOWMAN C, et al. Field performance of elite transgenic maize plants expressing an insecticidal protein derived from Bacillus thuringiensis [J]. BioTechnology, 1993, 11(2): 194200.
[24]SIEBERT M W, NOLTING S P, HENDRIX W, et al. Evaluation of corn hybrids expressing Cry1F, Cry1A. 105, Cry2Ab2, Cry34Ab1/Cry35Ab1, and Cry3Bb1 against southern United States insect pests [J]. Journal of Economic Entomology, 2012, 105(5): 18251834.
[25]王振营,何康来,石洁,等.桃蛀螟在玉米上危害加重原因与控制对策[J].植物保护,2006,32(2):6769.
[26]石洁,王振营,何康来.黄淮海地区夏玉米病虫害发生趋势与原因分析[J].植物保护,2005,31(5):6365.
(责任编辑:杨明丽)
