SMAD1基因的沉默和过表达及对秦川牛原代成肌细胞生肌的影响
2019-06-11宁越米雪陈星伊邵建航昝林森
宁越,米雪,陈星伊,邵建航,昝林森,2
SMAD1基因的沉默和过表达及对秦川牛原代成肌细胞生肌的影响
宁越1,米雪1,陈星伊1,邵建航1,昝林森1,2
(1西北农林科技大学动物科技学院,陕西杨凌 712100;2国家肉牛改良中心,陕西杨凌 712100)
【目的】为了探索秦川牛()SMAD1基因在成肌细胞分化过程中的分子作用机制,通过研究沉默、过表达该基因后对牛原代成肌细胞分化及成肌相关基因表达量的影响,以期为BMP信号通路在牛肌肉组织生长发育过程中的作用机制提供理论依据。【方法】根据Genbank牛(NM_001077107.2)已知序列设计并合成靶向沉默SMAD1基因的干扰序列si和阴性对照siRNA-NC。以秦川牛原代成肌细胞cDNA为模板,利用PCR技术克隆获得SMAD1基因CDS序列,构建腺病毒过表达穿梭载体pDC316-mCMV-EGFP-b。将腺病毒基因组质粒和腺病毒载体穿梭质粒共转染到HEK 293细胞中,通过Cre-loxp重组酶获得重组腺病毒。获得的携带目的基因的腺病毒标记为AD-b,重组质粒pDC316-EGFP标记为AD-NC,用做对照组病毒,利用LaSRT法测定腺病毒的滴度。将获得的干扰序列si和过表达腺病毒AD-b分别侵染秦川牛原代成肌细胞,采用实时荧光定量PCR检测SMAD1基因和成肌标志基因、、的mRNA水平,并观察对细胞分化融合情况的影响。【结果】成功获得了1条能有效沉默SMAD1基因siRNA,将其转染秦川牛原代成肌细胞后进行人工诱导分化,相比对照组,SMAD1基因分别下调了75.4%(诱导1d,<0.01)、66.7%(诱导3d,<0.01)、60.0%(诱导6d,<0.01)、54.7%(诱导9d,<0.01)。此外,还成功构建了秦川牛SMAD1基因的腺病毒过表达穿梭载体,获得了滴度为1×1010pfu/mL的过表达重组腺病毒AD-b。与对照组相比,高滴度过表达病毒AD-b侵染秦川牛原代成肌细胞0、1、3、6、9d后,与对照组相比,SMAD1基因的表达水平分别上调了2.10倍(诱导1d)、3.19倍(诱导3d)、105.3倍(诱导3d),<0.01)和144倍(诱导9d,<0.01);结合肌管形成过程的变化和实时荧光定量结果证明,SMAD1基因促进原代成肌细胞分化和肌管形成,并显著上调成肌标志基因、、的表达。【结论】获得了能有效沉默秦川牛原代成肌细胞表达的特异性的干扰序列si,以及可成功实现过表达SMAD1基因的腺病毒介导的体外表达载体,可正向调控成肌细胞的分化和肌管形成过程。
SMAD1基因;RNAi;腺病毒载体;原代成肌细胞;肌管形成;秦川牛
0 引言
【研究意义】秦川牛()是我国著名的地方黄牛品种,具有耐粗饲、抗逆性(病)好、肉质好的优点,但仍存在生长慢、产肉量低且脂肪沉积欠佳等缺点[1]。随着人们生活水平的不断提高,牛肉供应紧缺,提高牛肉的产量、质量成为当务之急[2-4]。【前人研究进展】研究表明,骨骼肌生长和肌内脂肪含量对畜禽的产肉性能和肉质有着至关重要的影响。TGF-β(transforming growth factor-β)信号通路主要参与调节细胞生长、增殖、分化、迁移和凋亡,可介导组织与器官的正常生长和发育(胚胎发育、骨骼器官形成)、机体的免疫反应等生物过程,其在抑制脂肪细胞和成肌细胞分化中的作用十分重要[5-6]。SMAD蛋白家族作为细胞内重要的信号转导蛋白,直接参加与TGF-β超家族成员TGF-β1、TGF-β2、TGF-β3以及骨形态发生蛋白(bone morphogenetic protein, BMP)、活化素等多个成员的信号转导[7]。目前,共有8种SMAD成员被报道(SMAD1-8),按照结构和功能可分为3类[8-10]。第一类,特异性SMADs(亦称receptor- activated SMAD,R-SMADs),包括SMAD1、2、3、5、8,其中SMAD1、5、8能被BMP-Ⅰ型受体(ALK1、2、3、6)磷酸化而激活,参与BMP信号通路;而SMAD2、3能被活化素或者TGFβ-Ⅰ型受体(ALK4、5)和孤儿Ⅰ型受体(ALK7)激活,作为TGF-β信号通路的直接作用底物,可对成肌因子及成肌分化过程起关键作用。第二类,共有型SMADs(common SMAD, C-SMAD),即SMAD4,它能与激活的R-SMADs形成异源的多聚复合物,转录到细胞核内调控下游基因。第三类,抑制型SMADs(Ⅰ-SMADs),包括SMAD6、7,它们能抑制R-SMADs和C-SMAD复合物的活性。【本研究切入点】BMP信号通路作为调控骨/软骨形态发生的重要通路而被熟知。近年来,研究发现SMAD1/5/8通过参与BMP信号通路对调控肌肉量有着十分重要的作用[11]。试验发现,在鼠的骨骼肌细胞细胞核中定位到SMAD1、SMAD5、SMAD8蛋白,并且通过成年在鼠骨骼肌细胞中敲除SMAD1/5/8基因两周后,观察到其肌肉量减少了20%。此外,还有试验证实,BMP-SMAD1/5/8通路可通过与activin/ myostatin–SMAD2/3通路竞争SMAD4从而促进肌肉量及成肌分化过程[12]。【拟解决的关键问题】鉴于SMAD1基因的重要功能,本试验以秦川牛背最长肌组织为试验材料,克隆秦川牛SMAD1基因,设计合成了RNA干扰序列、构建了腺病毒过表达载体,获得了具有干扰能力的RNA序列si和对有过表达能力的腺病毒AD-b,研究其在秦川牛成肌细胞分化过程中的调控作用,为进一步研究该基因和BMP信号通路在秦川牛肌肉组织生长发育过程中发挥的作用奠定基础。
1 材料与方法
本研究于2016年9月至2018年4月在西北农林科技大学国家肉牛改良中心完成。
1.1 试验材料
秦川牛第1代原代成肌细胞[13]、背最长肌组织(第12—13肋骨间)均冻存于西北农林科技大学国家肉牛改良中心种质资源库。所用试剂有:pDC316-mCMV- EGFP载体质粒、JM109感受态细胞、Polyfectine转染试剂均购自百恩维生物科技有限公司(中国,深圳);DNA Polymerase、质粒小提试剂盒、Trans2K Plus II DNA Marker购自全式金生物技术有限公司(中国,北京);琼脂糖凝胶回收试剂盒、Endo-free Plasmid Kit购自Omega Bio-tek股份有限公司(美国);T4 DNA 连接酶、限制性内切酶I、I购自NEB有限公司(中国,北京);DMEM培养基、胎牛血清(FBS)、马血清(HS)、胰蛋白酶Trypsin 0.25% EDTA、Lipofectamine® 3000购自Invitrogen公司(美国);青链霉素双抗购自Gibco(美国);腺病毒纯化试剂盒ViraBind购于Cell Biolabs公司(美国);PrimeScriptTMRT Reagent Kit with DNA Eraser反转录试剂盒、SYBR® Primix ExTMⅡ试剂盒购自宝日医生物技术有限公司(中国,北京)。
1.2 生物信息学分析
登录NCBI数据库(https://www.ncbi.nlm.nih.gov/)下载包括牛()等9种动物的SMAD1氨基酸序列(表1)。利用DNAMan 6.0软件进行蛋白序列多重比对。
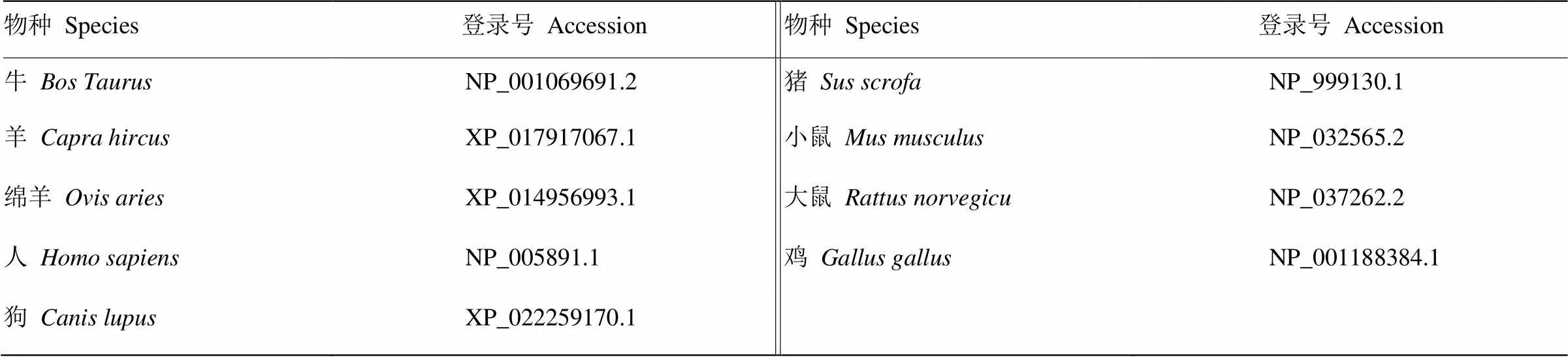
表1 9种物种SMAD1氨基酸序列信息
1.3 诱导分化秦川牛原代成肌细胞
复苏第1代的秦川牛原代成肌细胞,差速贴壁两次后进行传代培养至第3代,将细胞按2×105个/孔接种于6孔培养板中,使用未诱导分化培养基(20%FBS+DMEM/F12)培养。待细胞融合率达到80%后用含2%HS的分化培养基进行诱导分化培养,隔日换液。
1.4 沉默秦川牛SMAD1基因
1.4.1 设计并合成siRNA 按照siRNA设计原则,以Genbank上牛SMAD1基因mRNA序列(NM_ 001077107.2),应用Invitrogen公司在线设计工具(https://rnaidesigner.thermofisher.com/rnaiexpress/)设计1条靶向沉默SMAD1基因的siRNA前体寡核苷酸序列和1条阴性对照序列(表2)。

表2 siRNA序列
1.5 过表达牛SMAD1基因
1.5.1 构建秦川牛SMAD1基因腺病毒过表达载体 用Primer 5.0软件根据牛SMAD1基因mRNA序列设计特异性引物(表3),并在引物中引入酶切位点I和I。以原代成肌细胞cDNA为模板扩增CDS区序列,产物长1 452 bp。PCR体系为:含模板的DNA 100ng、正反向引物各4µL、dNTP Mix(2.5mmol·L-1)4µL、polymerase 1µL、5×buffer 10µL,用灭菌蒸馏水补足50µL。PCR扩增条件:95℃ 3 min;95℃ 20 s,55℃ 30 s,72℃ 2 min,30 个循环;72℃ 5 min。将PCR产物经双酶切(I和I)回收后与pDC316-mCMV-EGFP真核表达载体大片段进行连接,22℃反应2h。反应体系为:目的片段6µL、pDC316-mCMV-EGFP大片段2µL、10×DNA Ligase Buffer 1µL、T4 DNA Ligase 1µL。将上述连接产物取10µL与100µL JM109感受态细胞均匀混合,冰浴30min后42℃热激45s,再立即冰浴2min,加入预热至室温的400µL LB培养基后37℃摇床培养1h,以4 000r/min离心1min后弃去400µL培养上清,再用移液器将剩余100µL吹打混匀后涂布于LB平板上(氨苄抗性含100µg·mL-1),37℃恒温培养箱倒置培养过夜。次日挑取2个单克隆菌落并接种到5ml含氨苄抗性的LB培养液(氨苄浓度为100µg·mL-1)中,250r/min,37℃恒温摇床培养过夜,用小量质粒抽提试剂盒抽提质粒,质粒经I和I双酶切鉴定无误后,送上海生工生物工程股份有限公司进行测序。

表3 PCR引物序列
1.5.2 pDC316-mCMV-EGFP-b和对照腺病毒包装 将293细胞接种在6孔板中培养,细胞汇合度达到70%—80%,用转染试剂Polyfectine转染质粒,具体步骤参照说明书进行。转染后观察出毒迹象,即细胞收缩变圆,伴有细胞脱落。在大部分细胞病变并从培养瓶壁脱落后,作为病毒母液P1进行收毒。
扩增病毒AD-b。扩增培养HEK 293细胞,当细胞达到80%—90%汇合度时加入P1代病毒AD-b。在病毒侵染48h后收集细胞,在-80℃/37℃下冷冻/解冻3次,离心(10 000×g,10min)收集病毒上清,标记为P2。用P2代病毒以相同方法扩增病毒至P3代,再通过腺病毒纯化试剂盒对P3代病毒纯化。
PCR鉴定目的基因。收集P2代病毒,在沸水孵育10分钟后立即置于冰上。短暂离心后,取上清液作模板进行PCR反应。PCR鉴定条件与扩增目的基因相同。
1.5.3 病毒生物学滴度的测定 在滴度测定前一天将HEK 293细胞接种到48孔板中。用DMEM完全培养基10倍梯度稀释病毒。具体做法如表4:在96孔板每孔中加入90µL培养基,向第一横排孔中加入待测病毒原液各10µL,混匀后取10µL混合液转至第二横排孔,如此稀释至第八横排孔。取稀释好的病毒混合液10µL加入相应的48孔板中。侵染48h后,通过荧光显微镜计数荧光细胞情况。在最高稀梯度m的孔数出N个带荧光的细胞,则病毒滴度为N×10m/mL(m为稀释梯度),若N>10,则需继续稀释。

表4 病毒浓度梯度
1.5.4 测定腺病毒侵染细胞最佳MOI值 复苏实验室保存的第1代秦川牛原代成肌细胞,传代培养后,将细胞以1×104个/孔铺板至两个24孔培养板中。分别加入不同剂量的腺病毒(MOI分别为25、50、75、100、125和150),每个处理设置3个重复,48 h后根据细胞的生长情况及绿色荧光的表达情况来确定最佳的侵染浓度。
1.5.5 腺病毒侵染秦川牛原代成肌细胞复苏第1代秦川牛原代成肌细胞,传代培养到第3代时,将细胞以2×105个/ 孔铺至6孔培养板中,使用增殖培养基(20%FBS+ DMEM/F12)培养。试验共设计2个处理组,AD-b,AD-NC。当原代成肌细胞汇合度达到90%时,根据测定的MOI值加毒,12h后换成成肌细胞诱导分化培养基;侵染48h后,替换成分化培养基并隔天换液。在培养的第0、1、3、6、9天时收集细胞,提取总RNA并观察细胞分化融合情况。
俄罗斯一党制向多党制的过渡始于1991年叶利钦上台执政,其颁布的使俄罗斯政府机关非党化的总统令亦使党的新体制于苏共二十八大期间得以确立。多党制、政治多元化则在1993年获得通过的俄罗斯联邦宪法中被进而确认。自此在俄罗斯,多党制得以合法化。
1.6 细胞总RNA的提取及反转录PCR
用Trizol试剂提取处理第0、1、3、6、9天的成肌细胞总RNA,-80保存。用琼脂糖凝胶电泳和分光光度计测定提取的细胞RNA的质量。将质量符合要求的RNA用PrimeScriptTMRT Reagent Kit with DNA Eraser反转录试剂盒进行反转,反应得到的cDNA第一链保存于-20℃冰箱。
1.7 实时荧光定量PCR 检测
各取1µg RNA,用PrimeScriptTMRT Reagent Kit with gDNA Eraser反转录试剂盒进行反转录得到cDNA。用SYBR®Primix ExⅡ试剂盒进行实时定量PCR,反应体系为20 µL,其中SYBR®Primix ExⅡ 10 µL,上下游引物各0.8 µL,ROX Reference DyeⅡ 0.4 µL,cDNA模板2 µL,dH2O 6 µL。反应在7500 Real Time PCR仪(ABI公司)上进行,具体循环参数为:95℃预变性30 s;95℃变性5 s,60℃退火34 s,循环40次。熔解曲线程序为: 95℃ 15 s,60℃ 1 min,95℃ 15 s。每个样品进行3次试验重复。以GAPDH基因为内参,检测SMAD1基因及肌分化标志基因的表达。引物相关参数见表3。
1.8 统计分析
试验数据以“平均值±标准误(mean±SE)”表示,利用SPSS16.0软件进行单因素方差分析,用检验(-test)对不同试验处理之间的差异进行显著性分析,*<0.05为差异显著,**<0.01为差异极显著。
2 结果
2.1 物种间SMAD1蛋白质序列比对
对反刍动物、单胃动物等9种不同物种的蛋白质序列多重比较结果显示:牛()与山羊()、绵羊()、人()、狗()、猪()SMAD1蛋白质序列的相似性极高,均为99%;与小鼠()、大鼠()、鸡()SMAD1蛋白质序列的相似性稍低,分别为98%、97%和96%。因此,不难看出SMAD1蛋白序列在各物种间十分保守(图1)。
2.2 检测沉默效率
提取si转染牛原代成肌细胞第0、1、3、6、9天的细胞总RNA进行qRT-PCR。结果显示,与对照组相比,在转染si后,SMAD1基因分别被下调了75.4%、66.7%、60.0%、54.7%(<0.01,图2)。结果表明,通过RNA干扰技术可有效降低成肌细胞SMAD1基因的mRNA水平,且沉默效率至少可持续至第9天。

100%一致性氨基酸位点用白色背景标注,0%一致性的位点使用黑色背景标注,具体保守性在下方line图中

图2 siSMAD1干扰效率检测
2.3 构建pDC316-mCMV-EGFP-bSMAD1腺病毒载体
以秦川牛成肌细胞cDNA为模板,利用PCR技术扩增SMAD1基因CDS区序列,经1%琼脂糖凝胶电泳检测发现,条带清晰且符合预期(图3-A)。将PCR产物胶回收后克隆到pDC316-mCMV-EGEP载体上,构建pDC316-mCMV-EGFP-b腺病毒载体。质粒双酶切(I和I)后经电泳鉴定,得到5 803bp和1 452bp两条条带,所得线性化质粒和目的片段大小符合预期(图3-B),同时测序结果也显示pDC316-mCMV-EGFP-b腺病毒载体构建成功。
2.4 包装、扩繁pDC316-mCMV-EGFP-bSMAD1和对照腺病毒
共转染携带目的基因的重组质粒pDC316- mCMV-EGFP-b和骨架质粒到HEK293细胞中,在倒置荧光显微镜下观察到(图4 A-C):转染1d后有大量绿色荧光蛋白(GFP)表达;9d后出现彗星状的荧光聚集现象;在12—14d后大部分细胞病变并从培养瓶壁脱落,将其收集为病毒母液P1。为了获得强侵染能力的病毒,通过反复“侵染-冻融-收集”对病毒AD-b进行大量扩繁[14]。图4-D是第3代纯化后的腺病毒侵染细胞48 h时GFP表达情况。腺病毒滴度采用LaSRT法测定为1×1010pfu/mL。

A:秦川牛SMAD1基因CDS区序列PCR产物;B:pDC316-mCMV-EGFP-bSMAD1质粒酶切鉴定结果。M1:DL2000 DNA marker;M2:Trans2K plus marker;泳道1,2为SMAD1基因在两种成肌细胞cDNA中的PCR产物;泳道3为pDC316-mCMV-EGFP-bSMAD1的NheI和NotI双酶切鉴定结果
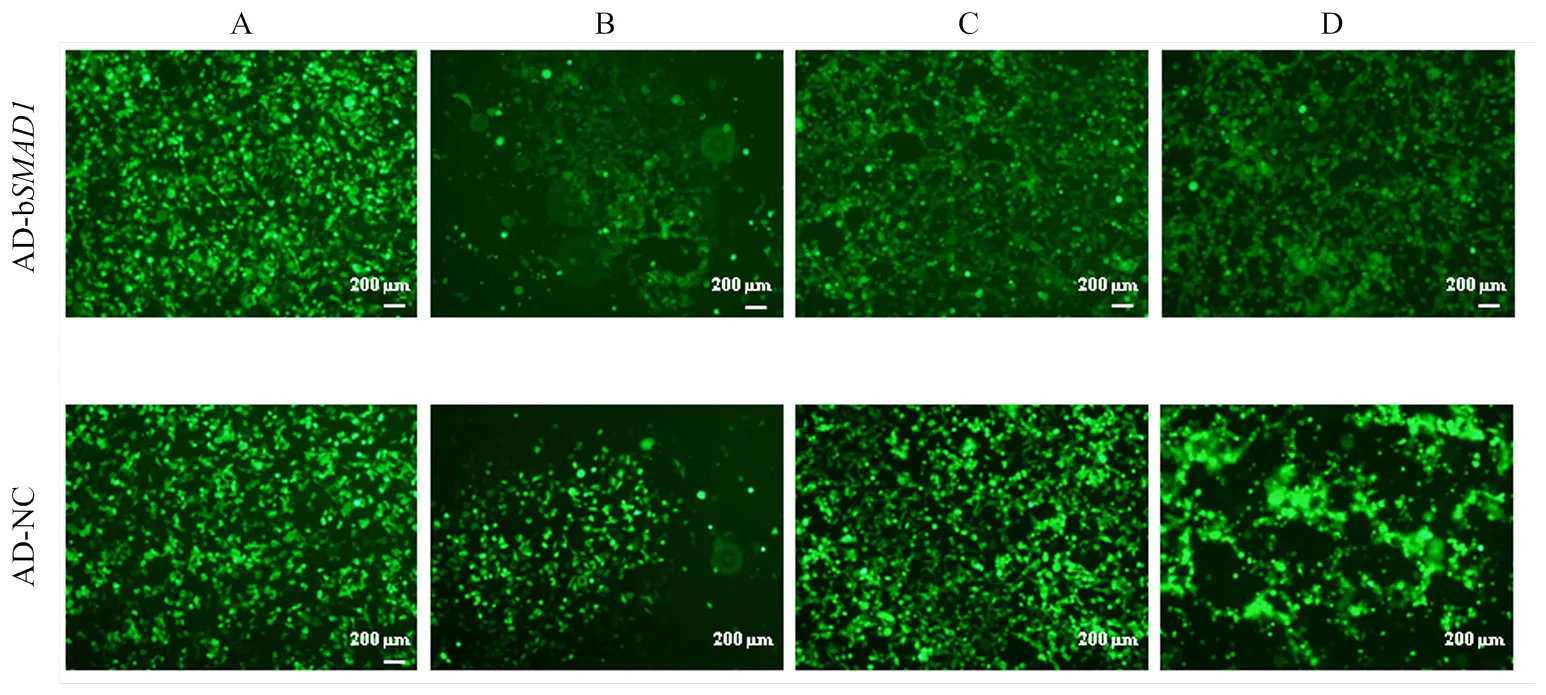
A:转染目的质粒到293细胞24h,GFP表达情况;B:转染9天后,病变细胞开始出毒,绿色荧光呈现葡萄串样的聚集现象;C:AD-bSMAD1转染14d以及AD-NC转染12d时,大部分细胞收缩变圆,开始从培养瓶壁脱落,将其收集为病毒母液P1;D:高滴度病毒(P3)侵染293细胞48h,GFP表达情况。原始放大倍数:40× (OLYMPUS IX71, 比例尺:200 µm)
2.5 测定腺病毒侵染细胞最佳MOI值
复苏的秦川牛原代成肌细胞经培养后得到的第3代细胞,在倒置显微镜下观察发现细胞形态多呈短梭形或多角形,细胞核呈圆形或椭圆形。将其铺至24孔培养板中,分别侵染不同剂量的腺病毒。48 h后在显微镜下观察到AD-b,AD-NC的MOI值(multiplicity of infection, 侵染复数)分别为100和25时细胞GFP表达量较多且尚未出现明显病变,因此确定此浓度为最佳侵染浓度。
2.6 检测过表达效率
腺病毒AD-b侵染秦川牛前体成肌细胞第1、3、6、9天后,在荧光显微镜下观察到95%以上的细胞可成功表达GFP。分别提取各时期细胞的总RNA,利用实时荧光定量PCR检测SMAD1基因的表达水平,结果显示SMAD1基因的表达量较对照组分别上调了约2.10倍(诱导1d)、3.19倍(诱导3d)、105.3倍(诱导6d,<0.01)和144倍(诱导9d,<0.01,图5)。上述结果说明所包装的过表达腺病毒可有效升高SMAD1基因的mRNA水平,且过表达效率可至少持续至第9天。
2.7 SMAD1基因促进秦川牛成肌细胞分化
显微镜观察转染si第6天的牛原代成肌细胞发现,处理组细胞的肌管形成过程较对照组明显受损,所形成的肌管更小、更短、更少(图6-A)。腺病毒AD-b侵染并诱导成肌细胞分化6天时,在显微镜下观察发现,与对照组相比,AD-b侵染组的细胞肌管的更大、更长、更多(图6-B)。由实时荧光定量PCR结果知(图6-C),与siRNA-NC相比,si处理组的肌分化标志基因,和的mRNA水平均显著下降(<0.01)。反之,与对照组AD-NC相比,AD-b组的成肌标志基因,和的mRNA水平均显著升高(图6-D,<0.01)。以上结果表明,SMAD1基因能够有效的正向调控秦川牛原代成肌细胞分化。

图5 AD-bSMAD1过表达效率检测
3 讨论
大理石花纹作为衡量牛肉品质的重要指标,主要受到肌肉和脂肪含量的比例影响。一直以来,人们对BMP信号通路的研究主要集中在骨的形成发育过程上,但自2013年起,其在调控肌肉发育上的重要功能开始为人们了解。研究证明,BMP信号通路在调控成年后的肌肉量变化的重要作用主要是由SMAD1/5/8完成的[11-12],且这条新的信号通路可以同广泛存在的TGF-β通路的activin/myostatin– SMAD2/3轴竞争结合SMAD4。这种竞争作用,一方面可以降低activin/ myostatin–SMAD2/3的作用浓度,减少其对肌肉发育的抑制作用;另一方面可以增加BMP-SMAD1/5/8的优势作用。获得优势的SMAD1/5/8磷酸化后可同SMAD4结合,进而将信号从细胞膜转录至细胞核内进而调控下游靶基因的转录,从而促进肌肉发育。
牛SMAD1基因定位在其17号染色体,编码区长1 398个碱基,共编码465个氨基酸。本试验主要通过在秦川牛成肌细胞中下调、上调SMAD1基因表达量,观察其对肌管融合过程及研究标志基因在肌生成过程中作用,从而研究该基因在成肌分化中的重要作用。与传统的基因工程技术相比,RNA干扰(RNAi)是一种高效、稳定、特异性强的干扰技术。因此本试验首先设计并筛选到一条有效的siRNA。试验发现,转染可有效下调目的基因的表达量,在转染后的第1、3、6、9天分别下调了75.4%、66.7%、60.0%、54.7%(<0.01)。说明所合成的si可持续稳定、高效得下调基因的mRNA水平,且该沉默水平可至少维持9d。另外,为了从另一角度研究基因功能,采取将外源基因导入真核细胞从而实现过表达目的基因以观察相关生理过程的变化。与脂质体介导的化学试剂转染法、电转移、核转移等物理方法相比,这种过表达方法能直接将目的基因插入到病毒基因组中,再利用病毒的侵染性实现目的基因在真核细胞内的过表达,最大程度的保护细胞保持良好状态而不受损伤。20世纪中期,重组腺病毒载体作为一个新兴的表达载体,开始应用于表达外源基因。与其他病毒载体系统相比,腺病毒载体系统可以有效地侵染原代细胞,并且可以侵染分裂期、非分裂期和增殖缓慢的细胞以及体内组织,更易获得高滴度病毒、可携带大片段外源基因而不与宿主基因组整合等[15-19]。腺病毒载体的这些优点使其成为细胞试验最常用的病毒载体之一,也是基因表达和传递的主要候选载体[14, 20]。本试验采用秦川牛原代成肌细胞所提的RNA进行反转录,以得到的cDNA为模板,在高保真酶的作用下成功克隆得到秦川牛SMAD1基因编码区序列。为了更好地研究SMAD1基因在秦川牛肌肉的生长发育中的调控作用,本试验选用Ad5腺病毒改造的AdMaxTM重组腺病毒包装系统,而不是前期使用的AdEasyTMAdenoviral Vector System。前期使用的这种腺病毒包装系统需要将目的基因线性化后转染到含有骨架载体pAd-Easy-1的.BJ5183大肠杆菌感受态中,再在细菌中实现腺病毒的重组[21-22]。而本试验采用的AdMaxTM系统是由Ad5腺病毒改建,不需要多次连接,可直接将共转染到HEK293细胞中的腺病毒载体穿梭质粒和骨架质粒在重组酶的作用下产生重组腺病毒,病毒包装不需要不断传代就可获得高滴度和高侵染效率的腺病毒[14, 23]。包装成功的病毒扩繁到第三代时滴度可达到1×1010pfu/mL。按MOI值(AD-b,AD-NC分别为100和25)侵染秦川牛原代成肌细胞第6和9天时,与对照组相比,SMAD1基因的表达水平显著上调了105.3倍(<0.01)和144倍(<0.01)。由此获得了可有效调控基因表达的工具,si和AD-b,为下一步研究SMAD1基因在成肌分化过程中的作用奠定了强有力的基础。

A:转染siSMAD1并诱导分化6天时牛原代成肌细胞分化融合情况;B:侵染腺病毒AD-bSMAD1并成肌诱导分化6天时秦川牛原代成肌细胞分化融合情况;C:沉默SMAD1后肌分化标志基因的mRNA水平检测;D:过表达SMAD1后肌分化标志基因的mRNA水平检测。原始放大倍数:40× (OLYMPUS IX71, 比例尺:200 µm)
骨骼肌分化是一类由多种信号通路和转录因子调控的复杂的级联反应[24-26]。成肌因子能够诱导肌肉干细胞的激活和终末分化,在成肌增殖和分化中发挥重要作用。在成肌发生的早期阶段,甚至是在增殖的成肌细胞分化前,MyoD和Myf5作为成肌分化的早期决定因子首先被激活。MyoD在骨骼肌卫星细胞的自我更新和增殖方面起到至关重要的作用[27]。已有研究表明,在肌卫星细胞中缺失 MyoD,会使卫星细胞更倾向于自我的更新交替,而不会进行细胞的融合形成肌管[28]。MyoD被激活后可进而调控下游成肌基因MyoG的表达,是在成肌终末分化期起决定性作用的转录因子,进而促进肌管的形成[29-32]。最新研究表明,SMAD1对成年后肌肉量的调控十分关键,但其具体的调控机制及在肌肉形成过程中的功能还不甚清楚。为了更好地研究SMAD1基因的作用机制,本试验利用前期获得的有效RNA干扰序列和高滴度腺病毒侵染秦川牛原代成肌细胞,通过对SMAD1基因mRNA水平表达量及细胞分化融合情况观察,发现SMAD1基因的表达量在转染si后的诱导分化1—9d内均极显著低于对照组,但是诱导1—6d的干扰效率在60%以上,而第9天的干扰效率有所下降(为54.7%),此外,在诱导第6天时成肌细胞的分化过程已呈现出明显的细胞表型(具体表现为肌管融合过程明显受损,所形成的肌管更小更少更短);而过表达SMAD1基因后其mRNA水平从细胞诱导分化的第六天开始显著上调且超过100倍,对照组相比,所形成的肌管与更多、更长。因此,综合干扰、过表达SMAD1基因后其mRNA水平和细胞表型的变化来看,诱导分化第6天都是一个很重要且有效的时间点,可以作为后续检测标志基因的时间点。另外,实时荧光定量PCR结果证明,SMAD1基因可正向调控成肌基因、的表达。以上结果提示,很有可能参与了成肌分化的早期/终末期调控,是成肌分化的一个正向调控基因。而对、的准确调控机制仍需进一步探究。
4 结论
本研究成功筛选到靶向沉默秦川牛SMAD1基因的RNA干扰序列si,同时成功构建了秦川牛SMAD1基因过表达重组腺病毒载体。通过在秦川牛原代成肌细胞中沉默、过表达SMAD1基因,检测相关标志基因的转录表达,证实SMAD1基因可正向调控原代成肌细胞分化。
[1] 昝林森. 牛生产学. 北京: 中国农业出版社, 2007.
ZAN L S.. Beijing: China Agricultural Press, 2007. (in Chinese)
[2] 柳泽新, 张宏兴, 闫文杰, 昝林森. 陕西畜牧业发展现状、存在问题及对策建议.畜牧兽医杂志, 2017, 36(3): 35-36.
LIU Z X, ZHANG H X, YAN W J, ZAN L S. Current situation, existing problems and suggestions of animal husbandry development in Shaanxi province.. 2017, 36(3):35-36. (in Chinese)
[3] 昝林森, 梅楚刚, 王洪程. 我国肉牛产业经济发展形势及对策建议. 西北农林科技大学学报(社会科学版), 2015, 15(6): 48-52.
ZAN L S, MEI C G, WANG H C. Current situation, existing problems and countermeasures on development of beef cattle industry in China.. 2015, 15(6): 48-52. (in Chinese)
[4] 昝林森, 梅楚刚, 王洪程. 中国黄牛选育改良及肉牛种业发展建议. 中国牛业科学. 2016, 42(6): 1-4.
ZAN L S, MEI C G, WANG H C. Breeding an improvement of Chinese yellow cattle and suggestions on beef cattle industry.. 2016, 42(6): 1-4. (in Chinese)
[5] DROGUETT R, VERRUGIO C C, SANTANDER C, BRANDAN E. TGF-β receptors, in a Smad-independent manner, are required for terminal skeletal muscle differentiation., 2010, 316, 2487-2503.
[6] 王今越. 转化生长因子β信号与肌肉调控. 中国组织工程院, 2016, 20(33):4979-4984.
WANG J Y. Transforming growth factor-beta and muscle regulation., 2016, 20(33): 4979-4984. (in Chinese)
[7] 陈兵, 易斌, 鲁开智. Smad蛋白家族调控细胞分化的研究进展. 医学研究生学报, 2013 26(5): 544-547.
CHEN B, YI B, LU K Z. Advances in researches on Smad proteins in cell differentiation., 2013, 26(5): 544-547. (in Chinese)
[8] SAVAGE C, DAS P, FINELLI A L, TOWNSEND S R, SUN C Y, BAIRD S E, PADGETT R W.genes sma-2, sma-3, and sma-4 define a conserved family of transforming growth factor beta pathway components., 1996, 93(2): 790-794.
[9] SHI Y, MASSAGUE J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus., 2003, 113(6):685-700.
[10] DIJKE P T, HILL C S. New insights into TGF-beta-Smad signalling.2004, 29(5):265-273.
[11] SARTORI R, SCHIRWIS E, BLAAUW B, BORTOLANZA S, ZHAO J H, ENZO E, STANTZOU A, MOUISELE E, TONIOLO L, FERRY A, STRICKER S, GOLDBERG A L, DUPONT S, PICCOLO S, AMTHOR H, SANDRI M. BMP signaling controls muscle mass., 2013, 45(11): 1309-1318.
[12] SARTORI R, GREGOREVIC P, SANDRI M. TGFβ and BMP signaling in skeletal muscle: potential significance for muscle-related disease., 2014, 25(9): 464-471.
[13] WANG Y N, YANG W C, LI P W, WANG H B, ZHANG Y Y, ZAN L S. Myocyte enhancer factor 2A promotes proliferation and its inhibition attenuates myogenic differentiation via myozenin 2 in bovine skeletal muscle myoblast., 2018, 13(4): 1-16.
[14] LODDE B M, DELPORTE C, GOLDSMITH C M, TAK P P, BAUM B J. A recombinant adenoviral vector encoding functional vasoactive intestinal peptide., 2004, 319(1): 189-192.
[15] SCHMID M, ERNST P, HONEGGER A, SUOMALAINEN M, ZIMMERMANN M, BRAUN L, STAUFFER S, THOM C, DREIER B, EIBAUER M, KIPAR A, VOGEL V, GREBER U F, MEDALIA O, PLUCKTHUN A. Adenoviral vector with shield and adapter increases tumor specificity and escapes liver and immune control., 2018, 9(450): 1-16.
[16] HE T C, ZHOU S, DACOSTA L T, YU J, KINZLER K W, VOGELSTEIN B. A simplified system for generating recombinant adenoviruses., 1998, 95(5): 2509-2514.
[17] LEE J, LAKS H, DRINKWATER D C, BLITZ A, LAM L, SHIRAISHI Y, CHANG P, DRAKE T A, ARDEHALI A. Cardiac gene transfer by intracoronary infusion of adenovirus vector-mediated reporter gene in the transplanted mouse heart., 1996, 111(1): 246-252.
[18] 魏胜娟, 昝林森, 王洪宝, 成功, 季舒涵, 王虹, 付常振, 姜碧杰.秦川牛FABP4基因重组腺病毒载体的构建与鉴定. 中国农业科学, 2013, 46(7): 1434-1440.
WEI S J, ZAN L S, WANG H B, CHENG G, JI S H, WANG H, FU C Z, JIANG B J. Construction of the recombinant adenovirus of FABP4 gene of Qinchuan cattle., 2013, 46(7): 1434-1440. (in Chinese)
[19] 王红娜, 孙洪新, 张英杰, 刘月琴, 谷振惠, 王苏瑶. 腺病毒介导shRNA干扰绵羊MSTN基因效果及对生肌调节因子和干扰素反应基因表达的影响. 畜牧兽医学报, 2017, 48(10): 1833-1842.
WANG H N, SUN H X, ZHANG Y J, LIU Y Q, GU Z H, WANG S Y. Adenovirus-mediated shRNA interference ofgene and its effect on expression of myogenic regulatory factor and interferon response gene in sheep., 2017, 48(10): 1833-1842. (in Chinese)
[20] YIN G, CHEN J, WEI S, WANG H, CHEN Q, LIN Y, HU J, LUO E. Adenoviral vector-mediated overexpression of osteoprotegerin accelerates osteointegration of titanium implants in ovariectomized rats., 2015, 22(8): 636-644.
[21] LUO J Y, DENG Z L, LUO X J, TANG N, SONG W X, CHEN J, SHARFF K A, LUU H H, HAYDON R C, KINZLER K W, VOGELSTEIN B, HE T C. A protocol for rapid generation of recombinant adenoviruses using the AdEasy system., 2007, 2(5): 1236-1247.
[22] 付常振, 昝林森, 王虹, 姜碧杰, 成功, 王洪宝, 朱光星, 李耀坤, 王洪程. 秦川牛SREBP1基因重组腺病毒载体的构建与病毒包装.畜牧兽医学报, 2013, 44(8): 1323-1329.
FU C Z, ZAN L S, WANG H, JIANG B J, CHENG G, WANG H B, ZHU G X, LI Y K, WANG H C. Construction of recombinant asenovirus vector specific togene of Qinchuan cattle and the packaging and amplifying of the corresponding adenovirus., 2013, 44(8): 1323-1329. (in Chinese)
[23] VORBURGER S A, HUNT K K. Adenoviral gene therapy., 2002, 7:46-59.
[24] WEINTRAUB H. The MyoD family and myogenesis: redundancy, networks, and thresholds., 1993, 75(7): 1241-1244.
[25] BERTI F, NOGUEIRA J M, WOHRLE S, SOBREIRA D R, HAWROT K, DIETRICH S. Time course and side-by-side analysis of mesodermal, pre-myogenic, myogenic and differentiated cell markers in the chicken model for skeletal muscle formation., 2015, 227(3): 361-382.
[26] BERKES C A, TAPSCOTT S J. MyoD and the transcriptional control of myogenesis., 2005, 16(4-5): 585-595.
[27] MONTARRAS D, MORGAN J, COLLINS C, RELAIX F, ZAFFRAN S, CUMANO A, PARTRIDGE T, BUCKINGHAM M. Direct isolation of satellite cells for skeletal muscle regeneration., 2005, 309(5743):2064-2067.
[28] GUSTAFSSON M K, PAN H, PINNEY D F, LIU Y L, LEWANDOWSKI A, EPSTEIN D J, EMERSON C P.is a direct target of long-range Shh signaling and Gli regulation for muscle specification., 2002, 16(1):114-126.
[29] OLIANI C, LIDANI K C F, GABRIEL J E. Differentiated evolutionary relationships among chordates from comparative alignments of multiple sequences of MyoD and MyoG myogenic regulatory factors., 2015, 14(4): 12561-12566.
[30] RICHARDSON R B, CURRIE P D. The genetics of vertebrate myogenesis., 2008, 9(8): 632-646.
[31] BUCKINGHAM M, VINCENT S D. Distinct and dynamic myogenic populations in the vertebrate embryo., 2009, 19(5): 444-453.
[32] YOKOYAMA S, ASAHARA H. The myogenic transcriptional network., 2011, 68: 1843-1849.
(责任编辑 林鉴非)
Silencing and Overexpressing SMAD Family Member 1 () Gene and Its Effect on Myogenesis in Primary Myoblast of Qinchuan Cattle ()
NING Yue1, MI Xue1, CHEN XingYi1, SHAO JianHang1, ZAN LinSen1,2
(1College of Animal Science and Technology, Northwest A&F University, Yangling 712100, Shaanxi;2National Beef Cattle Improvement Center, Yangling 712100, Shaanxi)
【Objective】The aims of the present study were to investigate molecular function ofgene in the bovine myoblast cell differentiation and to explore its role in the growth and development of Qinchuan cattle for beef production.【Method】SMAD1 gene was silenced and overexpressed through RNA interference and adenovirus recombinant over expression vector containingThe negative control siRNA-NC and RNA interference specific targeted SMAD1 gene (si) were designed and synthesized against the bovinemRNA sequence (CDS), which was obtained by RT-PCR using cDNA of Qinchuan cattle myoblast as template, and then, which was inserted into a shuttle vector pDC316-mCMV-EGFP to construct the over expression of adenovirus shuttle vector pDC316-mCMV-EGFP-b. Adenovirus genome plasmid and adenovirus shuttle vectors were cotransfected into HEK293 cell line to pack and obtain recombinant adenovirus. Adenovirus containing target gene was named as AD-b,whilepDC316-EGFP was used as a reference vector and was considered as control group "AD-NC". Their titers were tested by using LaSRT method. Bovine myoblast were transfected with siand AD-b, the mRNA level of SMAD1 gene and myogenesis-related genes, such as,and, were detected by real-time quantitative PCR, and the impact on myotube formation was observed. 【Result】The mRNA level of SMAD1gene transfected with siRNA was reduced 75.4% (induced differentiation for 1 day,<0.01), 66.7% (induced differentiation for 3 days,<0.01), 60.0% (induced differentiation for 6 days,<0.01), 54.7% (induced differentiation for 9 days,<0.01), respectively, compared with the siRNA-NC. The optimum titer of AD-binfectious was found at 1×1010pfu/mL. The expression levels ofwere rapidly increased 2.10 (induced differentiation for 1 day), 3.19 (induced differentiation for 3 days), 105.3 (induced differentiation for 6 days,<0.01) and 144 (induced differentiation for 9 days,<0.01) times of the control group after infected by AD-b, respectively. In addition, both myotube formation and qRT-PCR results proved that SMAD1 gene promoted myoblasts differentiation and myotube formation, as well as the mRNA expression level of,and. 【Conclusion】We could conclude from the present study thatSMAD1 gene performed its function in the myoblast differentiation and myotube formation. Based upon these findings, SMAD1 gene could be used as potential marker for the breed improvement of Qinchuan cattle through marker assisted selection for beef production.
mothers against decapentaplegic homolog 1 gene (); siRNA; adenovirus vector; primary myoblast; myotube formation; Qinchuan cattle
10.3864/j.issn.0578-1752.2019.10.014
2018-12-06;
2019-02-26
国家重点研发计划(2018YFD0501700)、国家肉牛牦牛产业技术体系(CARS-37)、陕西省科技统筹创新工程计划(2016KTCL02-15)
宁越,E-mail:ningyueny@163.com。通信作者昝林森,E-mail:zanlinsen@163.com
