籽粒苋优质贮藏蛋白AhAMA1基因功能分析
2019-06-01赵熙宁宋亚楠季春丽崔红利李润植
赵熙宁,张 飞,岳 敏,安 茜,宋亚楠,季春丽,崔红利,李润植
(山西农业大学分子农业与生物能源研究所,山西 太谷 030801)
籽粒苋(Amaranthus hypochondriacus L.)是苋科(Amaranthaceae)苋属(Amaranthus)1年生植物,可作为粮、饲、药、菜等多种用途的救荒作物[1-6]。籽粒苋富含油脂、蛋白质、碳水化合物和矿物质等,其中,种子油脂含量在10%以上,高于玉米、小麦和水稻等一般禾谷类作物,且油脂中角鲨烯等不饱和脂肪酸含量高达70%~80%[2],是一种健康型食用油脂资源;籽粒蛋白质含量大于30%,接近牛奶[3];种子贮藏蛋白品质好,没有引起人体过敏反应的蛋白,8 种人体必需氨基酸含量均高于其他谷类作物籽粒[4]。因此,籽粒苋营养丰富,是一种营养均衡、适宜于所有人群的健康食源。此外,籽粒苋亦是一种优良的高产饲料作物,其茎叶柔嫩,适口性好,纤维素含量低,粗蛋白质含量为17.7%~27.1%,用作青贮饲料,营养价值较高。其青贮饲料和籽实可代替部分精饲料或饵料,用于畜禽及水产业[5-6]。
源于籽粒苋成熟种子的AMA1 蛋白是一种优质贮藏蛋白,其中,人体必需氨基酸含量高且均衡。该蛋白不会导致人体过敏反应,所含8 种人体必需氨基酸含量都高于FAO/WHO 建议的优质蛋白质标准。已有研究将编码该蛋白的基因AhAMA1 应用于水稻籽粒遗传改良,以提高稻米营养品质[7]。
本研究从籽粒苋(Amaranthus hypochondriacus L.)(cv.SX-04)发育种子中分离到 AMA1 基因编码序列(AhAMA1),并检测其时空表达谱;应用生物信息学工具系统分析AhAMA1 蛋白理化特性;进一步构建AhAMA1 基因组成型植物表达载体,并通过农杆菌介导渗透侵染法在本氏烟草(Nicotiana benthamiana)叶组织瞬时表达AhAMA1,检测烟叶组织蛋白质、油脂、淀粉含量等生理生化性状,旨在评估AhAMA1 基因是否可用于改良生物量大的植物营养器官蛋白含量和营养品质。
1 材料和方法
1.1 试验材料
本研究所用的籽粒苋(Amaranthus hypochondriacus L.)种质(编号为 SX-04)与本氏烟草(Nicotiana benthamiana)种质材料均由山西农业大学分子农业与生物能源研究所提供。
1.2 试验方法
1.2.1 籽粒苋不同组织总RNA 提取及AhAMA1基因表达量分析 以播种后40 d 籽粒苋盆栽苗的根、茎、叶以及不同发育时期种子(开花后7,12,17,22 d)为材料。按照Trizol 法提取各组织总RNA。-80 ℃保存备用。在http://biodb.swu.edu.cn/qprimerdb查找,获得本氏烟草内参基因NtActin 引物NtActin-F,NtActin-R;获得籽粒苋内参基因AHYPO 引物AHYPO-F,AHYPO-R。依据 GenBank 中籽粒苋(Amaranthus hypochondriacus)AhAMA1 基因转录本序列(Z11577.1)ORF(915 nt)设计 PCR 扩增引物:全长克隆引物AhAMA1-F1(加粗序列为XbaⅠ)、AhAMA1-R1(加粗序列为SmaⅠ);全长PCR 引物AhAMA1-F2,AhAMA1-R2;定量 PCR 引物 AhAMA1-F3,AhAMA1-R3(表1)。
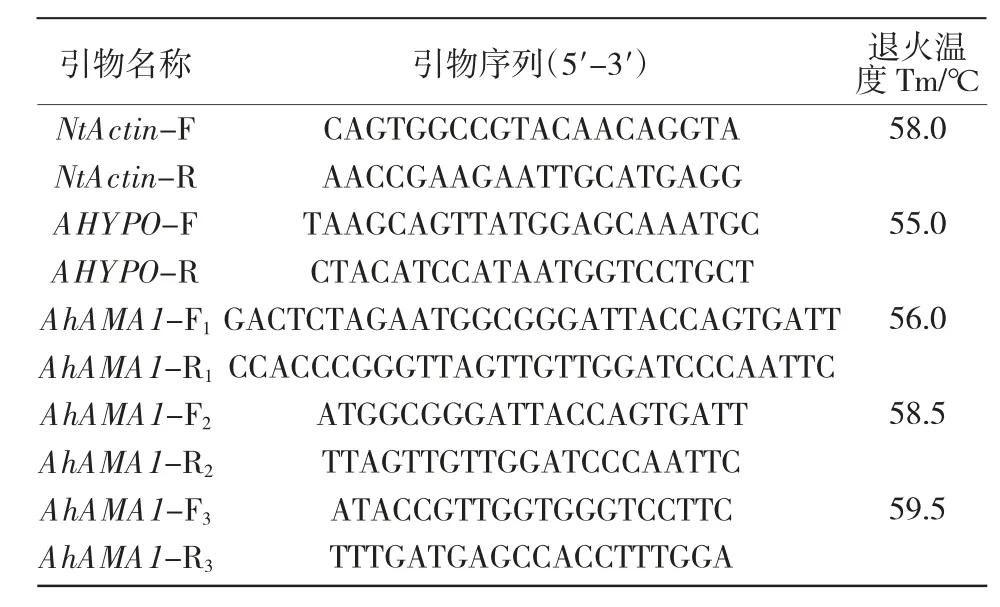
表1 PCR引物
1.2.2 AhAMA1 基因ORF 克隆 按照ABM 试剂盒使用方法,以经过浓度、纯度测试的发育种子总RNA 为模板,反转录得到cDNA,-20 ℃保存备用。高保真PCR 反应体系参照说明书:Sterilized ddH2O(20 μL),2×San Taq PCR Mix(25 μL),DNA(1 μL),上游引物(10 μmol/L,2 μL),下游引物(10 μmol/L,2 μL)。反应条件为:95 ℃预变性 5 min;S1:95 ℃变性 30 s,S2:退火 Tm(表1)30 s,S3:72 ℃延伸 40 s,循环数(内参、定量×25)(克隆、全长×35)次;72 ℃,10 min。按照琼脂糖凝胶DNA 回收试剂盒(Biomiga公司)操作说明书,将RT-PCR 扩增片段纯化回收,然后连接到PBI121 表达载体上。XbaⅠ,SmaⅠ双酶切验证连接成功后,使用热激法将重组质粒转入到大肠杆菌(Escherichia coli)感受态DH5a 细胞中,涂布于LB 琼脂培养基(含卡那霉素),37 ℃倒置培养。挑单克隆于LB 液体培养基扩大培养,PCR 检测后将有目的条带的菌落送至生工公司对重组质粒进行DNA 测序。
1.2.3 AhAMA1 蛋白理化特性的生物信息学分析通过NCBI 数据库Blast 发现,籽粒苋AhAMA1(AAM09540.1)与籽粒苋凝集素(AAD33922.1)和尾穗苋(Amaranthus caudatus L.)凝集素(AAQ03084.1)氨基酸序列相似性较高,进一步通过DNAMAN对这3 个蛋白进行同源性分析。使用ProtParam(https://web.expasy.org/protparam/)软件预测AhAMA1的氨基酸理论等电点、不稳定系数和总平均疏水性等理化性质;通过NCBI 预测分析AhAMA1 保守结构域;通过SignalP 分析AhAMA1 信号肽;通过NetPhos(http://www.cbs.dtu.dk/services/NetPhos/)分析AhAMA1 潜在磷酸化位点;通过SOPMA 软件折叠预测分析AhAMA1 蛋白二级结构;应用SWISSMODEL 对AhAMA1 蛋白进行三维结构预测。
1.2.4 农杆菌介导烟草瞬时表达验证AhAMA1 基因功能 将重组质粒AhAMA1-GFP-PBI121 以及PBI121 空载体通过热激法转入农杆菌(Agrobacterium)GV3101 中,在含有利福平和卡那霉素的LB琼脂培养基上28 ℃恒温倒置培养。挑取单克隆扩大培养后进行菌检,将有目的条带的农杆菌和含空载的农杆菌重悬后,采用渗透法侵染本氏烟草叶片。所侵染烟草叶片 3 d 后提取RNA,检测A-hAMA1 基因是否表达;侵染5 d 后的烟叶样品和对照烟叶样品,经冷冻干燥后,用于测定蛋白质、油脂、淀粉、必需氨基酸含量等生理生化性状。
2 结果与分析
2.1 籽粒苋不同组织器官AhAMA1基因表达分析
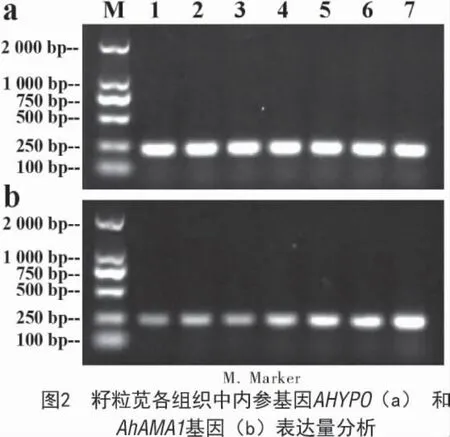
使用Trizol 法分别提取籽粒苋根、茎、叶以及不同发育时期种子总RNA。1%琼脂糖凝胶电泳检测结果显示(图1),各 RNA 样品的 28S,18S,5S 条带完整,质量符合试验要求;半定量PCR 检测各组织器官AhAMA1 基因表达表明(图2),各组织中烟草内参基因AHYPO(a)表达量一致(条带亮度相同),AhAMA1 基因(b)表达量有所差异(条带亮度不同),揭示AhAMA1 基因在所测各组织中均有表达,其中在发育种子中高量表达。
2.2 AhAMA1基因ORF克隆
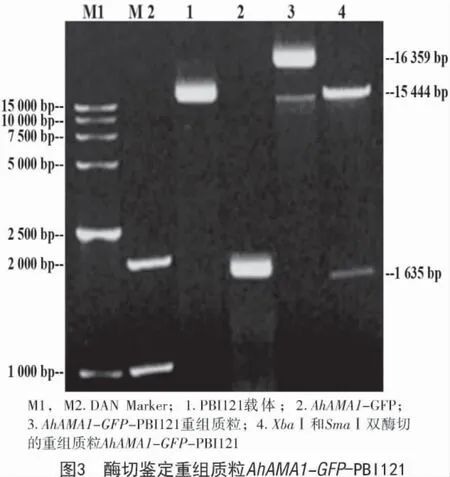
以籽粒苋(cv.SX-04)发育种子RNA 反转录获得的cDNA 为模板,用分别带有XbaⅠ和SmaⅠ酶切位点的目的基因引物进行高保真PCR,扩增AhAMA1 基因ORF,扩增的靶基因片段AhAMA1(915 bp)与 GFP(720 bp)基因连接后经 XbaⅠ和SmaⅠ双酶切后电泳检测结果如图3所示,PCR 扩增的条带与预期目的基因条带(1 635 bp)相符。经测序验证,该PCR 扩增产物序列与AhAMA1 基因ORF 序列完全一致。
2.3 AhAMA1基因植物组成型表达载体的构建
经XbaⅠ和SmaⅠ双酶切后的AhAMA1-GFP与经XbaⅠ和SmaⅠ双酶切的PBI121 表达载体进行连接获得重组质粒,再用重组质粒转化大肠杆菌感受态DH5a,获得阳性菌落。提取质粒,用XbaⅠ和SmaⅠ进行双酶切鉴定以及DNA 测序,结果如图3所示,AhAMA1 基因 ORF-GFP 亚克隆到 PBI121 载体的多克隆位点,获得了AhAMA1 基因组成型植物表达载体AhAMA1-GFP-PBI121(图4)。
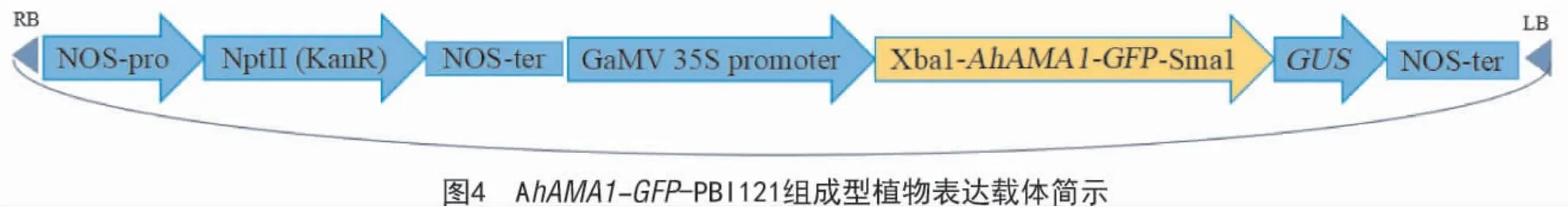
2.4 AhAMA1蛋白的生物信息学分析

使用NCBI 对AhAMA1 基因序列进行分析可知,AhAMA1 cDNA 全长为1 183 bp,起始密码子位于cDNA 的47 bp 处,终止密码子位于961 bp 处,开放阅读框(ORF)为915 bp,编码304 个氨基酸,其所编码的蛋白命名为AhAMA1 蛋白(AAM09540.1)。将其与籽粒苋凝集素(AAD33922.1)和尾穗苋凝集素(AAQ03084.1)进行氨基酸序列多重比对显示,其蛋白序列一致性高达99.12%,亲缘关系很近(图5)。
应用ProtParam 对AhAMA1 蛋白理化性质进行预测和分析,结果表明,其分子式为C1582H2406N412O461S12,原子总数为 4 873,相对分子质量为 34.96 ku。AhAMA1 蛋白由304 个氨基酸组成,其中,Arg+Lys(带正电)29 个、Asp+Glu(带负电)30 个,pI 值6.65;总平均亲水性-0.343,是亲水蛋白;不稳定指数值32.78,属于稳定蛋白。
应用NCBI 相关工具对AhAMA1 蛋白保守结构域预测分析表明,该蛋白属于Agglutinin 超家族(图6)。SignalP 分析显示,AhAMA1 蛋白不具有信号肽。NetPhos 潜在磷酸化位点分析显示,AhAMA1的磷酸化位点有丝氨酸(11 个)、苏氨酸(7 个)和酪氨酸(8 个)3 种。

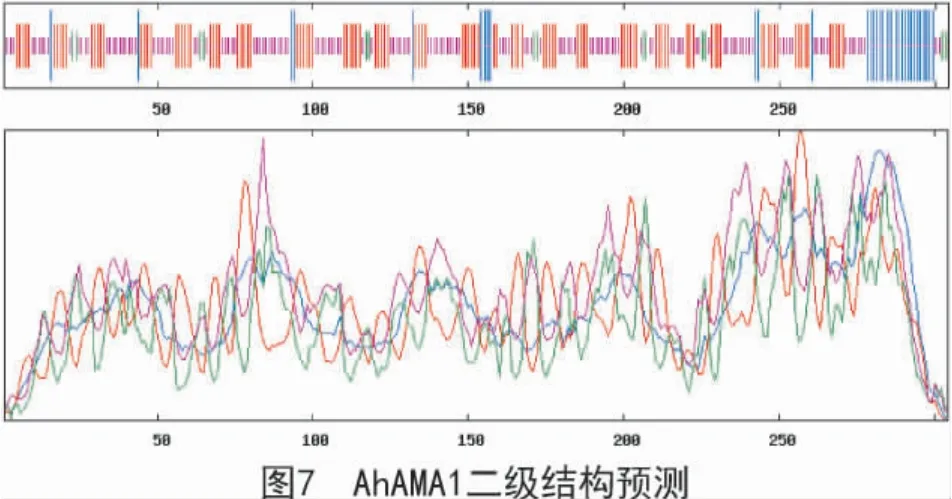
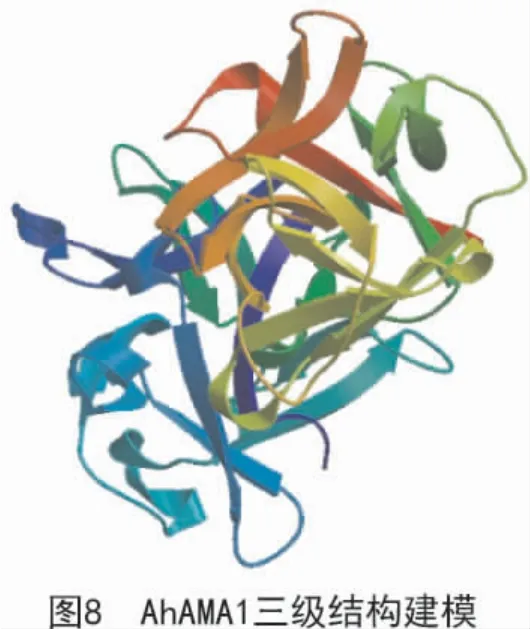
由SOPMA 软件折叠预测分析得出(图7),该蛋白二级结构由α 螺旋(蓝色区域)、伸展链(红色区域)、β 转角(绿色区域)和无规则卷曲(紫色区域)组成,且占比分别为11.51%,36.18%,4.93%和47.37%。应用SWISS-MODEL 软件对AhAMA1 蛋白进行三维结构图建模(图8),结果表明,该蛋白为单体,无配体;该蛋白的氨基酸序列与模板序列的一致性达到61%;X-射线晶体衍射发现,该蛋白是同型二聚体。
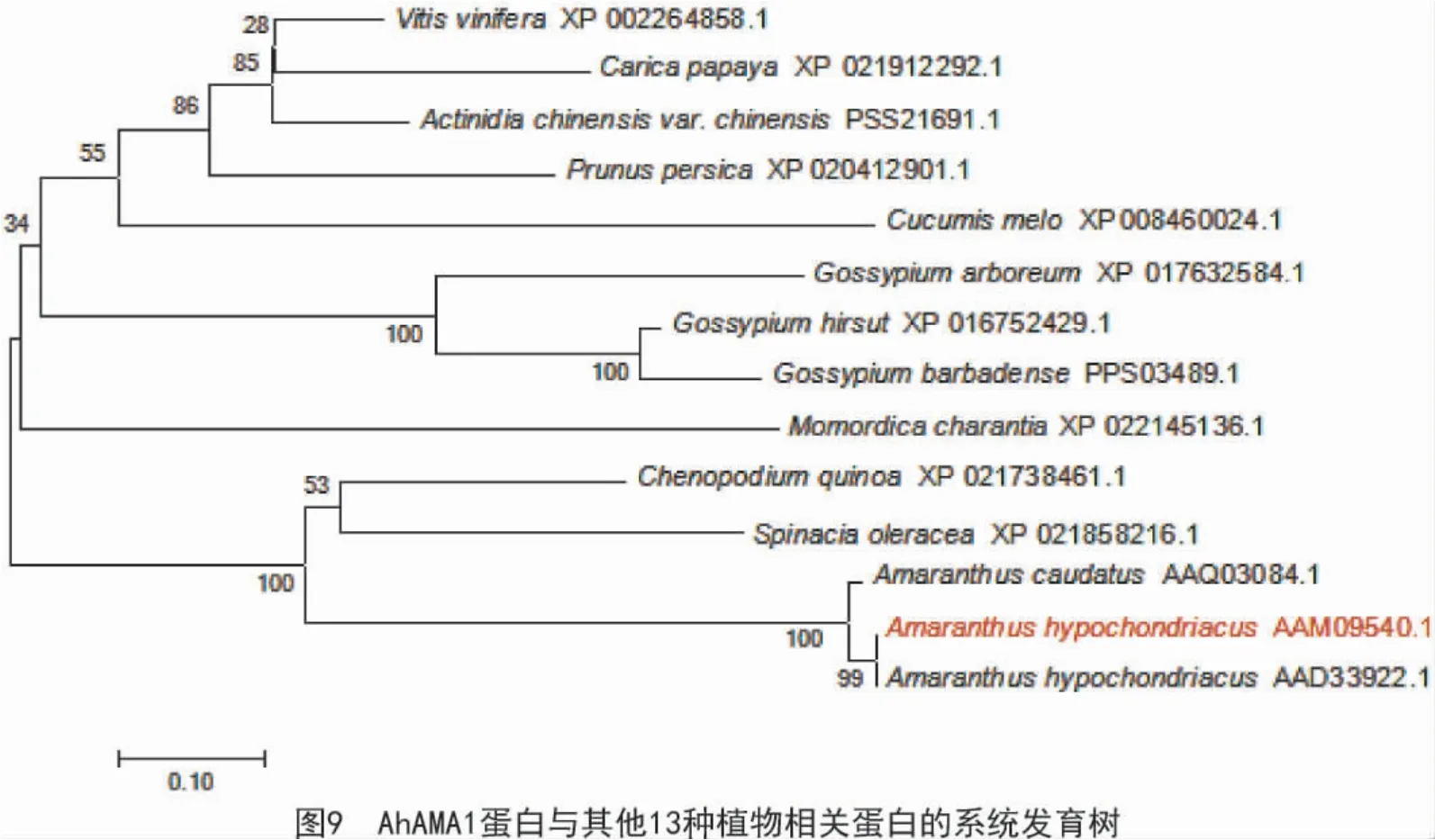
通过MEGA7 软件构建AhAMA1 蛋白(AAM09 540.1)与其他植物相关蛋白的系统进化(图9)结果显示,AhAMA1 蛋白与籽粒苋凝集素亲缘关系较近,这些蛋白包括籽粒苋凝集素(AAD33922.1)、尾穗苋凝集素(AAQ03084.1)、藜麦(Chenopodium quinoa willd)蛋白(XP_021738461.1)、葡萄(Vitis vinifera L.)蛋白(XP_002264858.1)、猕猴桃(Actinidia)蛋白(PSS21691.1)、番木瓜(Carica Papaya L.)蛋白(XP_021912292.1)、碧桃(Amygdalus persica L.var.persica f.duplex Rehd.)蛋白(XP_020412901.1)、菠菜(Spinacia oleracea L.)蛋白(XP_021858216.1)、甜瓜(Cucumis melo)蛋白(XP_008460024.1)、陆地棉(Gossypium hirsutum Linn.) 蛋白(XP_016752429.1)、海岛棉(Gossypium barbadense Linn.)蛋白(PPS03489.1)、木本棉(Gossypium arboreum(A2))蛋白(XP_017632584.1)和苦瓜(Momordica charantia L.)蛋白(XP_022145136.1)。
2.5 农杆菌介导AhAMA1基因在烟叶组织瞬时表达分析
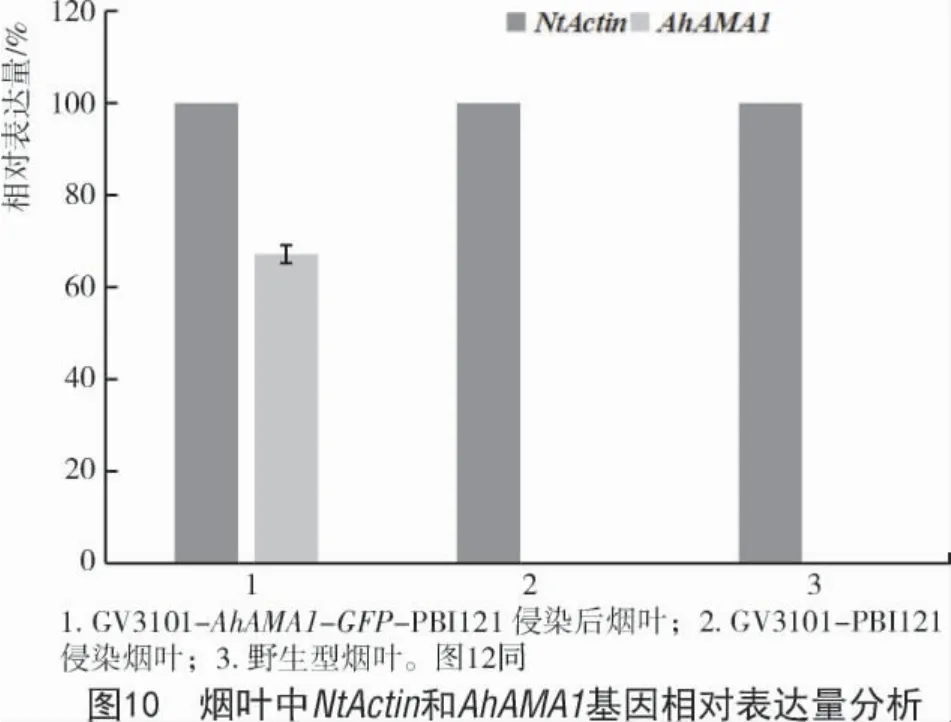
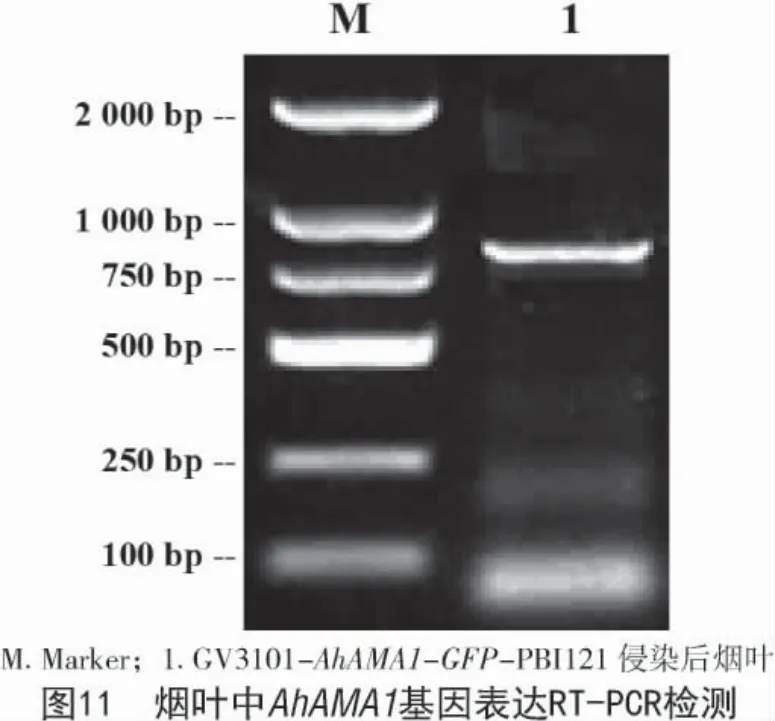
将重组表达质粒 AhAMA1-GFP-PBI121 和PBI121 空载分别通过热激法转化导入农杆菌GV3101 中,在含有利福平和卡那霉素的LB 琼脂培养基上28 ℃恒温倒置培养,挑取单克隆扩大培养后进行菌检。将有目的条带和含空载的农杆菌重悬后侵染烟草叶片。选用侵染2 d 后的烟草叶片提取RNA 检测,检测AhAMA1 基因是否成功表达,结果如图10,11 所示,AhAMA1 基因在烟叶组织中成功表达。
2.6 瞬时表达AhAMA1基因的烟叶油脂、蛋白质和淀粉积累分析
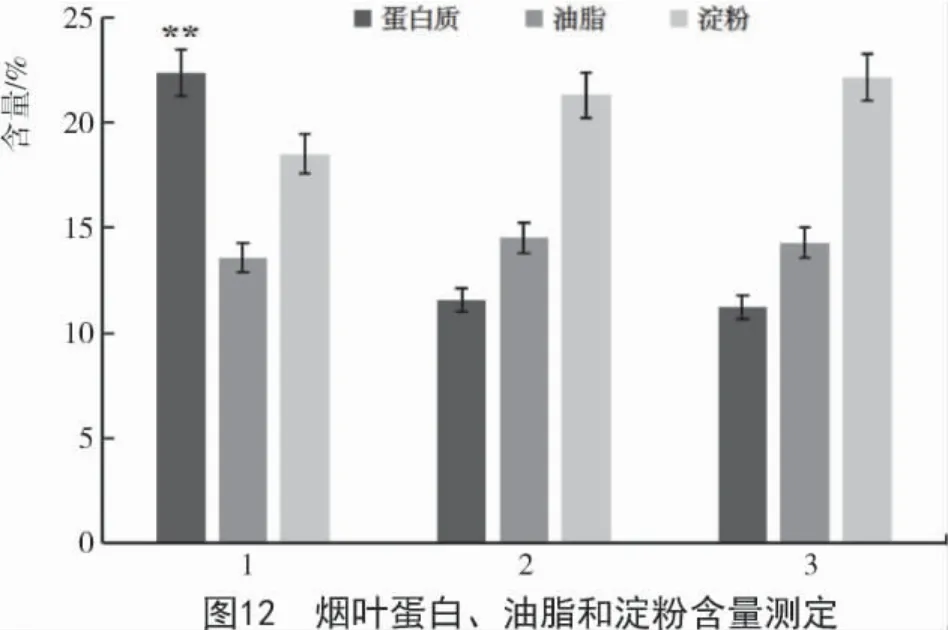
选取侵染5 d 后的烟叶样品,冷冻干燥后,萃取并测定蛋白质、油脂以及淀粉含量。如图12 所示,野生型烟草及对照叶片蛋白含量为11.19%,瞬时表达AhAMA1 基因的烟草叶片蛋白含量为22.39%,是野生型烟草的2 倍;油脂及淀粉含量检测相比野生型有所降低;必需氨基酸含量相比野生型均有增加(表2),异亮氨酸、亮氨酸、赖氨酸、甲硫氨酸、苯丙氨酸、苏氨酸、色氨酸、缬氨酸相比野生型分别增长 42%,53%,33%,48%,47%,11%,14%,21%。

表2 烟叶中必需氨基酸质量分数 mg/g
3 结论与讨论
农作物及饲草植物蛋白质含量及氨基酸组成决定其蛋白营养品质和用途。分离一些编码优质蛋白质的基因,应用于农作物等植物的遗传改良以提高其营养价值,近年来取得了一些可喜进展。ZHENG 等[8]利用PEG 法分别将来源于豌豆(Pisum sativum Linn)和菜豆(Phaseolus vulgaris Linn)的球蛋白基因整合到水稻基因组中,结果表明,菜豆球蛋白占总内胚乳蛋白质的4%。张秀君等[9-11]将马铃薯(Solanum tuberosum L.)花粉特异水溶性蛋白的cDNA 用基因枪法转入玉米(Zea mays L.)中,结果显示,获得的转基因玉米成熟种子蛋白含量提高,赖氨酸提高10%~54%。王为民等[12]将马铃薯高赖氨酸基因用基因枪法转入水稻(Oryza sativa)龙特甫B和中优早5 号,结果显示,龙特甫B 种子中蛋白质含量提高27.69%、赖氨酸含量提高16.15%;中优早5 号种子中蛋白质含量提高35.18%、赖氨酸含量提高了45.09%。高越峰等[13]将四棱豆(Psophocarpus tetragonolobus(L.)DC.)高赖氨酸蛋白基因 Lys 通过基因枪法转入水稻,结果显示,转基因植株叶片中赖氨酸含量均提高,最高增幅16.04%。王逸群等[14]将从辣椒(Capsicum annuum L.)成熟花粉中克隆的高赖氨酸蛋白基因CFLR 通过农杆菌介导法转入谷秆两用水稻,结果发现,9 个转基因植株叶片的赖氨酸含量均有提高,最高增幅22.71%。蒋家焕等[15]将四棱豆Lys 基因用农杆菌介导法转入水稻台粳9 号,并获得了转基因抗性植株,野生水稻种子赖氨酸含量仅为0.27%,而转基因水稻种子为0.349%,相比野生型提高了29.3%;转基因水稻种子蛋白含量提高了27.4%,转基因水稻秸秆蛋白含量提高了30.9%。孔维文等[16]将抗虫基因半夏(Pinellia ternata)凝集素基因PTA 和马铃薯的高赖氨酸基因SB401通过农杆菌介导法同时转入杂交水稻1826 中,获得了含PTA,SB401 基因的转基因水稻植株。吴超等[17]将四棱豆高赖氨酸基因通过农杆菌介导法转入脆茎粳稻中,结果发现,转基因水稻秸秆中赖氨酸含量最高为0.350%,粗蛋白含量最高为7.27%。李科等[18]将马铃薯中高赖氨酸蛋白基因SB401 通过农杆菌介导法导入优质水稻品种日本晴中,结果发现,9 个纯合的T4水稻转化株系种子的蛋白质、赖氨酸和其他氨基酸含量均提高。MOMMA 等[19]将大豆球蛋白基因通过电击法转入水稻,结果发现,转基因水稻种子中大豆球蛋白含量为40~50 mg/g,占种子总蛋白含量的4%~5%。这些研究表明,转基因提高植物蛋白含量和改良蛋白营养品质是可行的,但不同蛋白基因异源表达产生的效果不同。
许明等[20]采用 RT-PCR 方法,从籽粒苋(cv.千穗谷1 号)幼嫩种子克隆出AMA1 基因的完整开放阅读框序列,结果表明,该序列与GenBank 登录的籽粒苋AMA1 基因的核苷酸序列只有一个碱基的差异,但是不影响所编码氨基酸,原核表达获得了相应的目标蛋白;进一步将AMA1 基因在水稻胚乳中特异性表达,可一定程度提高水稻种子3 种限制性必需氨基酸含量。彭雪娟[7]用胚乳特异性启动子驱动AMA1 基因在水稻种子中表达,改进了稻米的蛋白营养品质。TAMÁS 等将[21]AMA1 基因通过基因枪转化法成功导入小麦中,结果使小麦氨基酸含量提高,而且面粉中有关面粉质量的功能参数得以改善。
本研究从籽粒苋种植材料(cv.SX-04)发育种子分离到 AhAMA1 的 ORF 序列(916 bp),与 Gen Bank 登录的籽粒苋AMA1 基因序列完全一致;AhAMA1 编码304 个氨基酸,分子量为34.96 ku。表达分析揭示该基因在发育种子后期高表达,与籽粒苋种子蛋白质快速富集期相吻合。成功构建了AhAMA1的组成型植物表达载体AhAMA1-GFP-PBI121,并通过农杆菌介导侵染使其在本氏烟叶组织中瞬时表达。烟叶生化测试显示,AhAMA1 瞬时表达导致烟叶组织中蛋白质含量显著提高,含量达22.39%,是野生型烟草叶片蛋白含量的2 倍;8 种必需氨基酸均有显著增加,且对油脂及淀粉含量影响未达显著水平。
总之,本研究表明,源于籽粒苋编码优质贮藏蛋白(8 种必需氨基酸含量高且均衡)的AhAMA1基因可以应用于高生物量植物营养器官的蛋白营养品质遗传改良,从而提高这些蛋白及8 种必需氨基酸用于功能饲料/饵料或加工其他产品的市场价值。
