夏、秋季渤海小型底栖动物类群组成及分布特征
2019-05-31刘晓收张志南
何 蕾,华 尔,刘晓收,张志南
中国海洋大学海洋生命学院,青岛 266003
渤海地处中国大陆东部,是我国最北端的近封闭型的内海,三面环陆,是中国近海大陆架上的浅海盆地,由辽河、滦河、海河、黄河等带来大量泥沙堆积而成,按海底地貌可划分为辽东湾、渤海湾、莱州湾、中央盆地和渤海海峡5个类型,其中,位于渤海湾和莱州湾之间的黄河口外有发育良好的水下三角洲。渤海沿岸河口浅水区营养盐丰富,饵料生物繁多[1],是黄渤海渔业的摇篮,是多种鱼、虾、蟹和贝类繁殖、栖息、生长的良好场所。小型底栖动物是生态系统中的重要组成部分,是许多经济鱼类、虾类和贝类幼体阶段的优质饵料, 因而是底栖食物网中十分重要的环节[2]。对小型底栖动物的丰度和生物量进行研究可为海洋生物资源的可持续利用和海洋农牧化生产提供基本依据,同时为渤海的生态动力学研究提供参数[3]。我国海域小型底栖动物的研究最早始于渤海。目前,渤海海域小型底栖动物的生态学研究已有一定的积累,获得了很多重要的阶段性成果[1,3-6]。本文根据2011年6月和11月在渤海海域获得的样品,研究小型底栖动物的数量分布及群落结构,目的是:探讨渤海海域小型底栖动物的数量分布格局、受控因子,及其季节动态特征;分析渤海海域小型底栖动物群落十年际动态。
1 材料与方法
1.1 研究海域及站位
分别于2011年6月(夏季航次)和2011年11月(秋季航次)搭载“东方红2号”科学调查船,对渤海海域(37.0°—40.0°N,118.0°—122.0°E)16个站位开展了2个航次的调查研究(图1)。其中,6月涉及15个站位,11月涉及10个站位,两个航次重复调查站位有9个,分别是B41、B42、B45、B47、B50、B51、B65、B70、BY01(图1)。
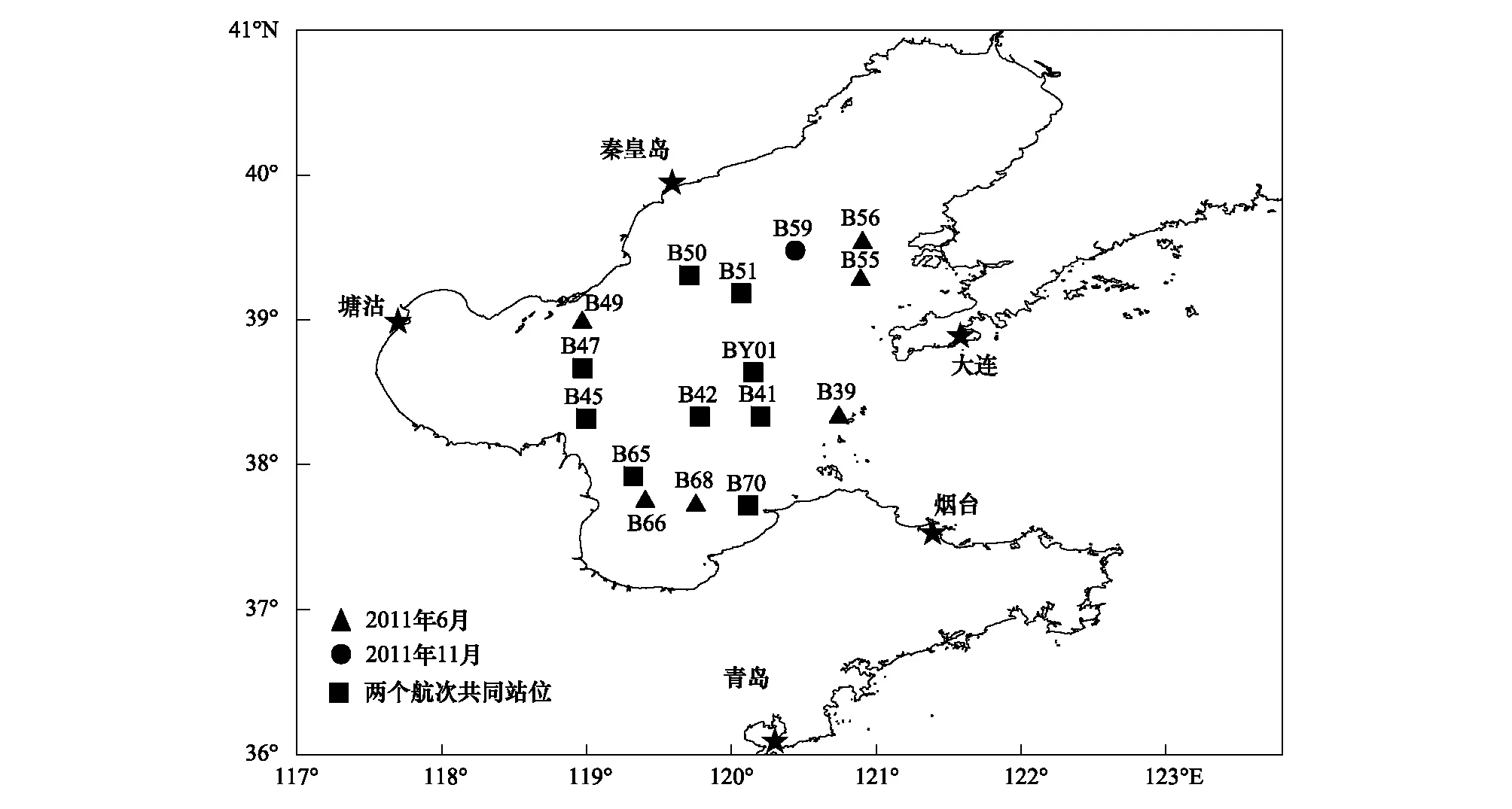
图1 渤海海域取样站位Fig.1 Sampling stations in the Bohai sea
1.2 取样方法
使用0.1 m2改进型Gray-O′Hara箱式采泥器取海底沉积物样品,并用内径为2.9 cm 的取样管(由塑料注射器改装)从中取从未受过扰动的3个重复样。芯样长度为8 cm,取出后立即按0—2、2—5和5—8 cm分层装入100 mL塑料样品瓶中,加入10%甲醛溶液进行固定,用于小型底栖动物研究分析。同时以相同方式另外取3个芯样按相应分层分别置于封口袋中,于-20℃冷冻避光保存,用于沉积物叶绿素a、有机质含量及粒度分析。其他环境资料(水深、底层水盐度、底层水温度等)由随船温盐深测定仪(CTD)现场测定。
1.3 室内分选计数
分选之前向每瓶样品中加入3—5 mL 1‰虎红荧光染液(Rose Bengal,由1 g虎红染料溶于1000 mL 5%的海水福尔马林溶液混合均匀得到),摇匀后静置染色24 h[7-8]。采用0.5和0.031 mm孔径的网筛套筛冲洗除去样品中的黏土、粉砂。将0.031 mm孔径的网筛上残留的沉积物样品用比重为1.15 g/cm3的Ludox-TM硅胶液转移至100 mL离心管中,以1800 r/min的转速离心10 min。将上清液倒出,重复离心3次。将3次离心所得上清液合并,通过0.031 mm 孔径的网筛,过滤掉Ludox溶液,用自来水将样品冲洗并转移至培养皿。在解剖镜下将小型底栖动物按不同类群分别计数,并收集至装有5%福尔马林溶液的塑料样品瓶中。
1.4 数据处理及分析
根据取样管的内径(2.9 cm), 计算每个站位的小型底栖动物丰度,即每10 cm2的个体数(个/10 cm2)。生物量的计算采用估算法,即以各类群丰度乘以个体平均干重。小型底栖动物不同类群个体的平均体重参照Jario[9],Widbom[10],Mclntyre[11]。生产力依照P/B=9估算。其中,P代表生产力,单位为μg干重 10 cm-2a-1,B代表生物量,单位为μg干重/10 cm2。
沉积物粒度分析采用Master Sizer3000型激光粒度仪进行测定。有机质测定参照《海洋调查规范》及刘昌岭等[12]改进的方法。Chl-a采用湿样法进行分析[13]。
使用Surfer8.0绘制站位分布图及丰度等值线图。使用SPSS19.0软件对小型底栖动物各类群丰度和生物量进行单因素方差分析(One-way ANOVA), 分析不同季节之间的差异;应用Spearman相关分析(Spearman correlation analysis)对各种环境因子与小型底栖动物的丰度和生物量进行相关性分析。使用PRIMER 6.0软件,进行等级聚类分析(CLUSTER),检验小型底栖动物类群组成在不同站位和季节之间的差异;进行BIOENV分析连接环境和生物矩阵,分析环境变量与小型底栖动物类群组成的相关性。
2 结果与分析
2.1 环境因子
海区整体水深较浅,2011年6月,15个站位的平均水深为23.8 m,11月10个站位的平均水深为25.1 m。6月各站位底层水温度(以下简称底温)平均值为15.7℃,水深最浅的B66站(14.0 m)和B65站(16.1 m)水温相对较高,分别为21.6℃和20.3℃(表1);呈现随水深和纬度升高底温降低的趋势。11月研究海域各站位间底温变化小,平均值为12.1℃。底层水盐度(以下简称底盐)的波动比较小,两个航次底盐分别为31.14±0.43和30.77±0.60。
研究海域沉积物类型主要包含4种:(1)黏土质粉砂(14个站位);(2)粉砂质砂(4个站位);(3)砂质粉砂(3个站位);(4)砂(2个站位)。B65站粉砂-黏土含量最高(6月100%,11月98.2%),B39站砂含量最高(6月85.5%)(表1)。沉积物的粉砂-黏土含量的高值区主要分布在黄河口外水下三角洲及以西的近岸水域,由近岸向外,沉积物中砂含量随水深增加而增加。
两个航次沉积物Chl-a含量均较低,平均值分别是0.79 μg/g和0.25 μg/g(B45、B59、B703个站位Chl-a的数据缺失)。其垂直分布表现为随着深度的增加而减少。
两个航次沉积物有机质含量与含水率差异小。其中,2011年6月航次沉积物有机质含量平均值为0.84%,11月为1.28%。沉积物含水率平均值分别为33.67%(6月)和37.09%(11月)。

表1 渤海海域各调查站位环境因子
表格中的B代表站位,与站位图中的对应
2.2 小型底栖动物的类群组成和时空分布
本次调查共鉴定出18个小型底栖动物类群,分别是线虫(Nematoda)、桡足类(Copepoda)、多毛类(Polychaeta)、动吻类(Kinorhyncha)、双壳类(Bivalvia)、介形类 (Ostracoda)、端足类(Amphipoda)、异足类(Tanaidacea)、涡虫类(Turbellaria)、涟虫类(Cumacea)、缓步类(Tardigrada)和甲壳类幼体(Nauplii)等(表2)。2011年6月航次共鉴定出14个小型底栖动物类群,没有发现等足类(Isopoda)、涡虫类(Turbellaria)、海螨类(Halacaroidea)、寡毛类(Oligochaeta),11月航次共鉴定出13个小型底栖动物类群,没有发现异足类(Tanaidacea)、涟虫类(Cumacea)、腹足类(Gastropoda)、缓步类(Tardigrada)和蛇尾类(Ophiuroidea)。海洋线虫为最优势类群,在两个航次中分别占小型底栖动物总丰度的94.95%和90.34%,总生物量的67.38%和45.32%。其次是桡足类和多毛类,其丰度和生物量所占比例11月有所增加(表2)。One-way ANOVA检验结果显示,研究海域11月多毛类丰度极显著地高于6月丰度(F=8.044,P<0.01)。
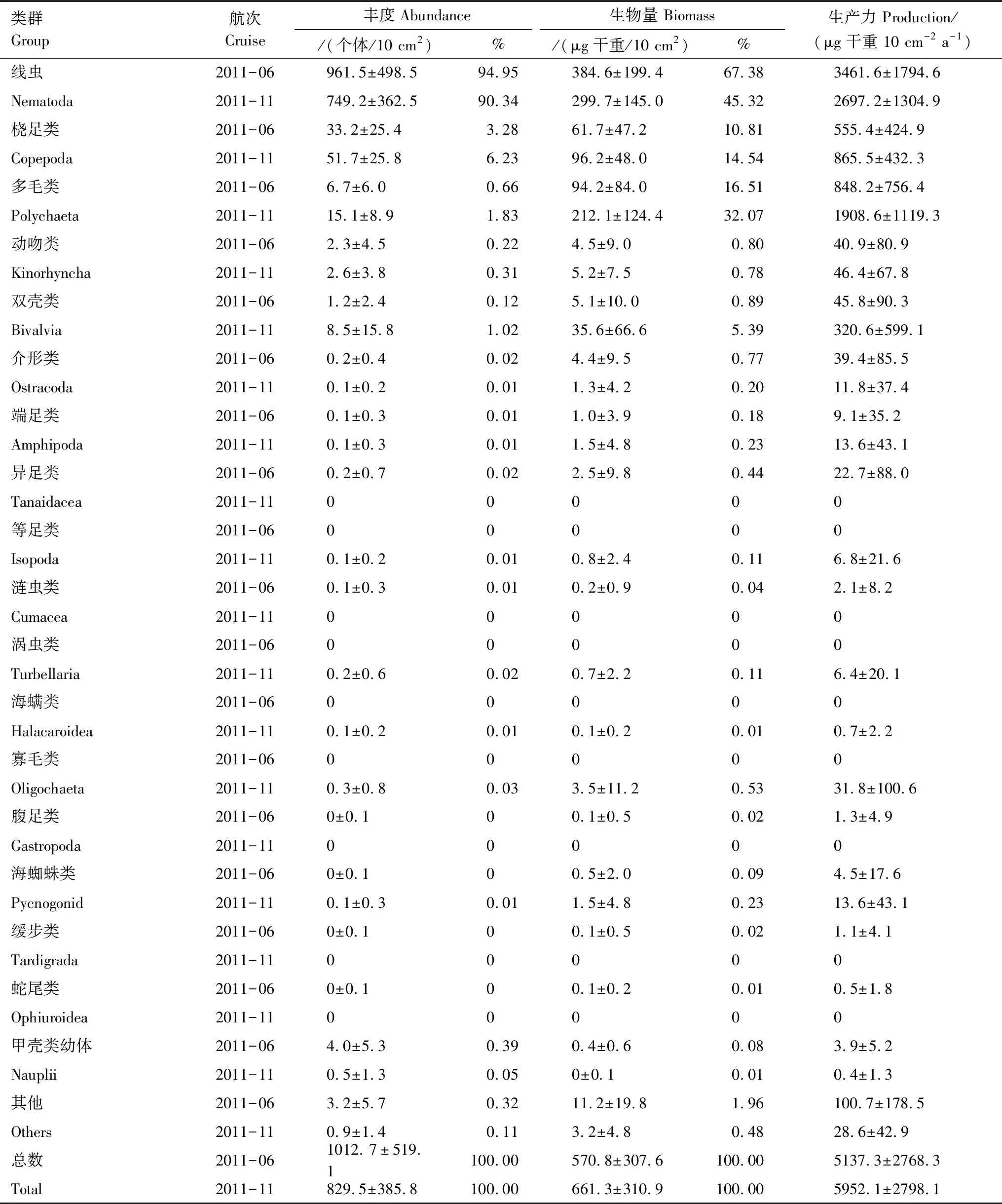
表2 2011年6月和11月小型底栖动物各类群的丰度、生物量和生产力
研究海域2011年6月的小型底栖动物平均丰度为(1012.7±519.1)个/10 cm2,最高值达到(2522.0±737.3)个/10 cm2,出现在研究海域最东部的B56站位;最低值出现在莱州湾口的B68站,为(425.6±200.0)个/10 cm2(图2)。小型底栖动物分布不均匀,One-Way ANOVA检验结果显示,各站位间差异极显著(F=3.567,P<0.01),表现为黄河口附近海域的小型底栖动物丰度较低,渤海中部小型底栖动物丰度较高(>1000个/10 cm2)。
2011年11月的小型底栖动物平均丰度为(829.5±385.8)个/10 cm2,最高值达到(1542.0±701.4)个/10 cm2,出现在渤海中部的B41站位;最低值出现在秦皇岛以南的B51站,为(369.1±124.7)个/10 cm2(图2)。11月的小型底栖动物的水平分布格局与6月相似。值得一提的是,研究海域北部近岸站位(B50和B51)11月小型底栖动物丰度显著低于6月丰度值(F=22.688,P<0.05),小型底栖动物低值区域由黄河口附近海域扩展至北部近岸海域。

图2 渤海小型底栖动物丰度的水平分布Fig.2 Horizontal distribution of meiobenthos abundance in the Bohai Sea
小型底栖动物生物量分布与丰度相似,2011年6月小型底栖动物生物量平均值为(570.8±307.6)μg干重/10 cm2,最高值出现在B56站位,为(1425.4±535.6)μg干重/10 cm2。2011年11月平均生物量为(661.3±310.9)μg干重/10 cm2,最高值出现在B70站位,为 (1255.3±723.3)μg干重/10 cm2。小型底栖动物2011年6月和11月的平均生产力分别为(5137.3±2768.3) μg干重 10 cm-2a-1和(5952.1±2798.1) μg干重 10 cm-2a-1。
基于小型底栖动物丰度数据进行的等级聚类CLUSTER分析显示:2011年6月,小型底栖动物群落可以划分为2组(图3):组1,主要由黄河口附近海域站位组成(6-B66,6-B68,6-B47,6-B50,6-B45,6-BY01),组2,由其他站位组成(6-B41,6-B39,6-B70,6-B65,6-B51,6-B42,6-B49,6-B55)。B56站位小型底栖动物类群组成与其他站位的相似性为66%,出现了涟虫类、异足类、海蜘蛛类、蛇尾类和缓步类等类群,类群多样性较高于其他站位。考虑其特殊性,分组时未予考虑。2011年11月,小型底栖动物群落也可以划分为2组(图3):组1(11-B42,11-B45,11-B47,11-B65,11-B50,11-B51,11-BY01)以及组2(11-B41,11-B59,11-B70)。
为准确评价两个航次间小型底栖动物群落特征的差异,本研究针对两个航次9个相同站位,进行小型底栖动物丰度、生物量及群落结构的比较。结果显示:2011年6月小型底栖动物平均丰度高于2011年11月,分别为(926.6±286.3)个/10 cm2和(665.4±402.7)个/10 cm2;而2011年6月平均生物量低于11月,分别为(505.6±186.3)μg干重/10 cm2和(519.1±301.8)μg干重/10 cm2。One-Way ANOVA检验结果显示,两个航次间丰度和生物量的差异均不显著(P>0.05),表明小型底栖动物丰度及生物量没有明显的季节变化。等级聚类CLUSTER分析显示,小型底栖动物群落在80%相似度水平可以划分为3组(图3):组1(11-B45,11-B51)、组2(11-B70,11-B41,6-B41)和组3(6-B42,6-B70,6-B65,6-B51,11-B50,11-BY01,6-B45,6-B47,6-B50,11-B47,11-B42,11-B65,6-BY01)。分组主要体现了研究站位的空间异质性,并未呈现季节更替引起的小型底栖动物类群组成的差异(ANOSIM,P>0.05)。

图3 小型底栖动物聚类分析图Fig.3 CLUSTER analysis based on meiofaunal abundance
2.3 小型底栖动物的垂直分布
两个航次中分别有93.6%(6月)和91.1%(11月)的小型底栖动物分布在0—5 cm的表层沉积物中。2011年6月和11月航次各站位的小型底栖动物垂直分布图显示,2个季节小型底栖动物垂直分布没有显著的差异(图4)。6月航次,小型底栖动物分布于0—2 cm、2—5 cm和5—8 cm的数量比例分别为68.2%、25.4%和11.0%。11月航次,小型底栖动物分布于0—2 cm、2—5 cm和5—8 cm的数量比例分别为62.2%、28.9%和8.9%。尽管有部分站位(例如B42、B45、B50、B70站位)出现0—2 cm层小型底栖动物丰度比例在2个采样季节间的差异,但并没有表现出显著的垂直迁移。

图4 小型底栖动物丰度的垂直分布Fig.4 Vertical distribution of meiobenthos abundance in the Bohai Sea
2.4 小型底栖动物与环境因子的关系
将研究站位小型底栖动物的丰度、生物量等生物数据与水深、沉积物粒度主要参数(中值粒径、粉砂-黏土含量等)、沉积物含水率、叶绿素a含量、有机质含量(%)等环境因子进行Spearman相关分析(表3),结果表明:小型底栖动物的丰度与中值粒径(R=-0.419,P<0.05)、沉积物含水率(R=-0.462,P<0.05)呈显著负相关,与有机质含量(R=-0.565,P<0.01)呈极显著负相关;小型底栖动物的生物量与中值粒径(R=-0.459,P<0.05)、粉砂-黏土含量(R=-0.443,P<0.05)、沉积物含水率(R=-0.460,P<0.05)、有机质含量(R=-0.426,P<0.05)呈显著负相关,与水深呈极显著正相关(R=0.533,P<0.01)。海洋线虫的丰度与小型底栖动物的丰度一致,与中值粒径(R=-0.425,P<0.05)、沉积物含水率(R=-0.475,P<0.05)呈显著负相关,与有机质含量呈极显著负相关(R=-0.572,P<0.01)。桡足类与所测环境因子未见显著的相关关系,表明研究海域受测环境因子可能不是影响其数量分布的主要影响因子。多毛类丰度与水深呈极显著正相关(R=0.570,P<0.01),与底层水温度呈显著的负相关(R=-0.441,P<0.05)。

表3 小型底栖动物的丰度、生物量与环境因子的相关分析结果
*在 0.05 水平(双侧)上显著相关;**在 0.01 水平(双侧)上显著相关
对小型底栖动物类群组成与环境因子的BIOENV分析显示,小型底栖动物群落不是受单一环境因子的影响,而是受多种环境因子的综合影响。水深、有机质含量能够最好地匹配小型底栖动物群落结构,相关系数为0.216。
3 讨论
3.1 渤海小型底栖动物分布特点
研究海域小型底栖动物分布不均匀,体现了小型底栖动物斑块分布的特点,表现为黄河口附近及近岸海域小型底栖动物丰度显著低于其他海域(P<0.05)。最优势类群海洋线虫丰度也表现此分布特征。研究海域,黄河径流带来大量泥沙的同时带来大量的陆源营养物质。但是,在黄河口附近以及近岸水域因为强烈的湍流和水体高浑浊度限制了浮游植物和底栖藻类的光合作用。因此沉向底部的藻类残体量少,以此为主要食物源的小型底栖动物丰度较低。随着黄河口及海湾沿岸向外水深逐渐增加,受河流影响减少,水体浑浊度减少,利于浮游植物和底栖藻类的光合作用,形成小型底栖动物的高值区。
小型底栖动物的分布及类群组成受许多沉积环境因子的综合影响。沉积物中的有机质含量、叶绿素a是底栖生物赖以生存的有机环境,是底栖生物的食物来源[14]。不同类型的底质以及水深、底层水盐度、沉积物含水率和有机质含量等是控制底栖生物群落分布与结构的重要因素[15-18]。本研究结果显示,小型底栖动物丰度与沉积物中值粒径、含水率和有机质含量呈显著的负相关,说明研究海域各站位沉积物粒级分配不同、有机质和含水率差异是引起小型底栖动物分布差异的主要因子。沉积物粒度特征是影响小型底栖动物群落最直接的环境因子。当沉积物粉砂质黏土含量降低,粒径增加时,其异质性增加,小型底栖动物多样性提高[19-20]。在砂质底沉积物中,小型底栖动物,特别是海洋线虫的多样性高于泥质底沉积物,同时也存在着更多的特异种[21]。本研究结果显示,B56站沉积物砂含量最高,小型底栖动物丰度最高、类群最丰富,显示了相似的结果。同时,沉积物粒度特征也可以通过影响其他环境因子,间接影响小型底栖动物群落。沉积物颗粒越细,含水率越高,有利于有机质保存,从而有机质含量越高。本文结果显示沉积物含水率、有机质含量与中值粒径具有极显著正相关关系(R=0.878和R=0.614,P<0.01),表明研究海域沉积物中值粒径对含水率和有机质含量的重要影响。因此,可以认为沉积物粒度是影响研究海域小型底栖动物数量和分布的最主要因子。
水深决定了沉降到海底的浮游植物碎屑的质量和数量,因此对小型底栖动物的分布及类群组成的影响非常重要[22]。在陆架海域,随水深的增加小型底栖动物可利用的浮游植物碎屑数量减少,会使小型底栖动物数量减少。Soltwedel等[23]的研究显示,小型底栖动物丰度及生物多样性随水深的增加而降低。本研究多毛类丰度与水深的显著相关关系表明,水深影响其丰度。此外,小型底栖动物类群组成与水深、有机质含量的相关性也说明,水深是影响研究海域小型底栖动物类群组成的主要因子之一。但是,渤海海域平均水深为24 m,海区整体水深较浅,变化梯度小;因此,对该海域小型底栖动物分布及类群组成的影响应为间接影响。
综上所述,本研究小型底栖动物的数量分布及类群组成受多种环境因子的综合作用。其中,沉积物中值粒径、含水率、有机质含量和水深是决定渤海海域小型底栖动物分布及类群组成的最重要因素。
3.2 小型底栖动物群落的季节动态
本文对两个航次9个相同站位的小型底栖动物丰度及群落结构的分析结果显示:小型底栖动物的平均丰度和平均生物量出现季节差异,但并不显著(P>0.05)。渤海是一个典型的北方温带半封闭海区,6月代表夏季,浮游植物和底栖藻类的生产力高,沉入底部的藻类残体量也较多,底栖动物食物来源丰富,小型底栖动物丰度高。而11月随温度降低和光照强度的减弱,浮游植物和底栖藻类的光合作用减少,因此沉入底部的藻类残体量变少,以此为主要食物源的小型底栖动物丰度降低。沉积物叶绿素a代表底栖藻类和沉降到底层的浮游藻类,是小型底栖动物的主要食物来源之一。本研究中,6月叶绿素a的含量极显著高于11月(F=16.129,P<0.01),表明小型底栖动物的食物来源有明显的季节变化。但是,9个相同站位小型底栖动物丰度与叶绿素a含量之间的正相关性不显著(R=0.356,P>0.05)。表明,研究海域叶绿素a含量的降低会引起小型底栖动物丰度降低。但是,由于生态因子间的补偿性作用特点,未能引起后者发生显著的变化。
本研究多毛类丰度在2个季节间具有显著的差异,11月高于6月(F=9.435,P<0.01)。有研究显示,多毛类丰度与有机质含量有关:当底质富含有机物时,多毛类数量较多,其生物量和栖息密度也较高,大多数小型多毛类的比例会增高,而肉食性和杂食性的种类会减少,个体平均大小减小[24]。本研究中,有机质含量未呈现显著的季节差异,多毛类丰度也未与有机质含量呈显著的相关性。但是,多毛类丰度与底层水温呈显著负相关,表明多毛类丰度受温度影响,出现季节性变化。
沉积环境的异质性、食物来源和小型底栖动物自身生殖补充等多种因素影响着生物的分布[9,25],要真正了解研究海域小型底栖动物的季节动态,必须进行连续几年的采样调查。
3.3 十年际小型底栖动物群落数量变化趋势
渤海海域小型底栖动物生态学研究始于20世纪80年代。受海上取样、室内分选方法及分选网筛孔径的差异,对不同时期小型底栖动物平均丰度的比较虽然存在一定的困难,也存在一定的误差,但是研究渤海海域小型底栖动物群落数量分布格局及十年际动态仍然具有一定的意义。
本研究两个航次渤海小型底栖动物丰度与张志南等[1]2008年渤海海域平均丰度无显著差异(表4);沉积物中值粒径、有机质含量和叶绿素a浓度平均值与2008年结果相近:沉积物中值粒径为5.15[1](2008)、5.02(2011-06)和5.59(2011-11);有机质含量为1.17%±0.68%[1](2008)、0.83%±0.48%(2011-06)和1.28%±0.30%(2011-11);叶绿素a浓度为0.812±0.643 μg/g[1](2008)、0.791±0.368 μg/g(2011-06)和0.245±0.047 μg/g(2011-11)。因此,本研究结果与2008年渤海研究结果可代表同一年代际不同季节的数据。本研究两个航次小型底栖动物平均丰度为(921.1±452.5)个/10 cm2,分别高于1998年和1999年渤海海域的平均丰度,低于1997年的平均丰度(详见表4)。线虫丰度的比较结果与此相同。需要指出的是1997年仅有5个采样站位,丰度偏高,不具代表性,不宜做平均水平的比较。如不考虑该年度,本研究与20世纪90年代末相比,渤海小型底栖动物(包括海洋线虫)丰度增加,而其中的底栖桡足类丰度却已减少(表4)。该结果与张志南等[1]研究中对渤海海域小型底栖动物30年来变化趋势的总结一致。
Zhou等的研究显示,20世纪80年代至90年代渤海沉积物粒度及有机质含量发生了显著变化,沉积物粒度变细,有机质含量增加,从而导致大型底栖动物群落结构发生变化,多毛类、双壳类及甲壳类等小型个体数量增加,棘皮动物等较大个体类群数量减少[26]。本研究结果与Zhou等[26]的研究结果相比,进入21世纪,渤海中部沉积物粉砂-黏土含量进一步增加,自20世纪80年代中值粒径由6.10降低至5.25(本研究平均值),有机质含量由0.53%增加至1.04%。沉积环境的变化必然影响栖息于此的底栖动物数量及分布。此外,渤海中部深水区既是黄渤海经济鱼、虾、蟹类洄游的集散地,又是渤海地方性鱼、虾、蟹类的越冬场,海洋资源的开发是环渤海地区经济发展重要的领域之一;受到人类活动的严重影响。因此,过去30年来,沉积环境的变化以及人类干扰是引起渤海海域小型底栖动物丰度变化的重要因素,与张志南等[1]研究结果一致。
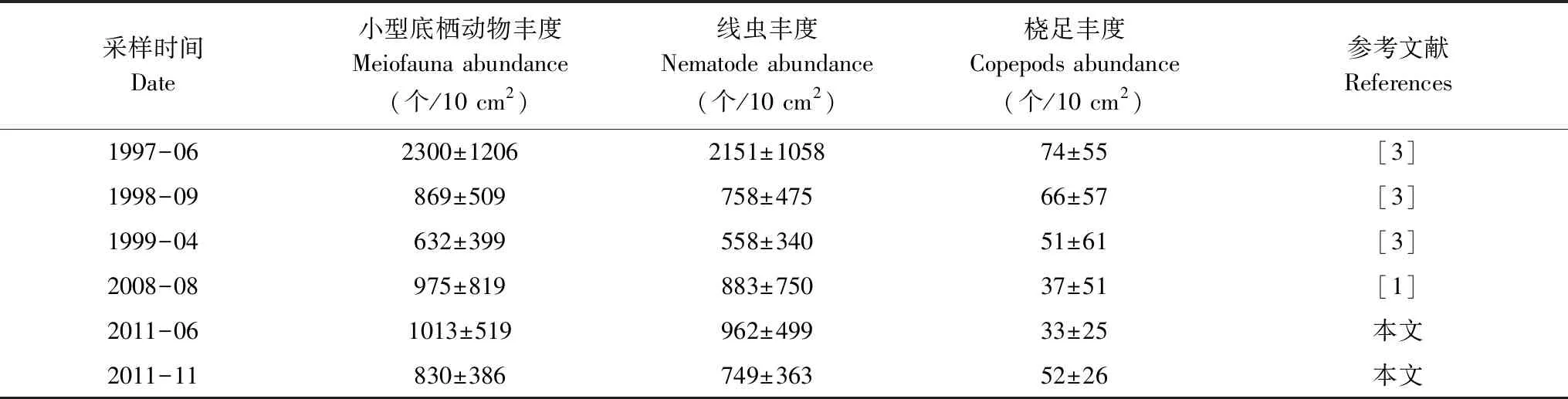
表4 本研究与历史资料小型底栖动物丰度的比较
4 结论
(1)研究海域小型底栖动物分布不均匀,表现为黄河口附近及近岸海域小型底栖动物丰度显著低于其他海域。最优势类群海洋线虫丰度也表现相似分布特征。沉积物中值粒径、含水率、有机质含量是决定研究海域小型底栖动物丰度的重要因素。
(2)研究海域小型底栖动物总丰度6月>11月,小型底栖动物总生物量6月<11月,但不构成显著的季节差异。最优势类群海洋线虫丰度和生物量也没有显著的季节差异。但是,多毛类11月丰度高于6月,表现出显著的季节差异。小型底栖动物各类群没有显著的季节性垂直迁移现象。
(3)小型底栖动物群落结构没有明显的季节变化,类群组成较稳定。与季节差异相比,小型底栖动物群落结构的空间异质性更显著。水深、有机质含量的组合能够最好地解释小型底栖动物群落结构的差异。
致谢:感谢王长江、付家霖、王礼鹏、战培旭和周腾腾协助完成小型底栖动物分选工作。感谢所有2011年6月和11月航次的调查人员。
