基于DNA 池重测序技术的肉兔HSFs 基因多态性分析
2019-05-27李丛艳梅秀丽邝良德郭志强张翠霞谢晓红
李丛艳,梅秀丽,邝良德,李 娟,雷 岷,郭志强,张翠霞,谢晓红*
(1.四川省畜牧科学研究院,四川成都 610066;2.动物遗传育种四川省重点实验室,四川成都 610066;3.成都市农林科学院畜牧研究所,四川成都 611130)
热休克因子(Heat Shock Factor,HSFs)是调控热休克蛋白(Heat Shock Protein,HSFs)最重要的转录因子,它通过与HSPs 基因启动子区的热休克元件(HSE)结合,启动基因的转录,促进HSPs 的表达[1]。不同的热休克因子在不同组织中表达,其结构很相似,但功能上有着不同程度的差异。HSF1 是动物体内最具代表性也是最重要的HSFs,在进化中高度保守并存在于各个物种中,在热应激反应诱导HSFs 的表达中发挥着重要作用[2-3]。研究发现HSF1 上多个SNPs 位点与牛的耐热性有关[4-7]。HSF4通过剪切变异体促进或抑制HSPs 的表达,从而调控热休克反应的动态平衡[8],其他HSFs(HSF2、HSF3、HSF5)具有物种特异性,针对不同的组织或在不同条件下激活[9-12]。
肉兔全身被毛且汗腺较少,夏季高温时非常容易出现热应激,严重影响其生长速度、繁殖性能以及抗病力等[13],因此培育在高温环境条件下仍能维持高生产水平的抗热应激肉兔群体是育种者的长期目标。四川白兔和闽西南黑兔是在我国(中)亚热带气候下长期选择形成的地方品种,对高温高湿环境具有自然耐受性;齐兴肉兔含有四川白兔血缘,耐热性较好;齐卡巨型兔为引进的大型肉兔品种,耐热性差;加利福尼亚兔为中型肉兔品种,耐热性能优于齐卡巨型兔。本研究以上述5 个耐热性能差异较大肉兔品种(系)为对象,采用DNA混池重测序和直接测序技术,对HSF1 和HSF4 基因进行多态性检测和群体遗传学分析,以期为筛选耐热相关分子标记、开展耐热肉兔品种培育提供参考。
1 材料与方法
1.1 实验动物 实验选择齐卡巨型兔、齐兴肉兔、加利福尼亚兔、四川白兔以及闽西南黑兔各30 只(公、母各半),共计150 个个体,采集5 g 左右耳组织样品,置于装有干冰的采样箱带回实验室,-20℃冰箱冻存备用。其中齐卡巨型兔、齐兴肉兔以及加利福尼亚兔来源于四川省畜牧科学研究院种兔场,四川白兔来源于国家级四川白兔保种场成都鼎鑫兔业有限责任公司种兔场,闽西南黑兔来源于安岳晨阳兔业有限公司。
1.2 基因组DNA 提取 按照动物组织样DNA 提取试剂盒说明书提取样品中的DNA,1%琼脂糖凝胶电泳检测后利用紫外分光光度计测量其OD 值,根据测量的浓度将样本稀释到50~100 ng/μL,-20℃保存以进行下一步实验操作。
1.3 DNA 池重测序筛选SNPs 以齐卡巨型兔、齐兴肉兔以及加利福尼亚兔为实验对象,运用DNA 混池重测序技术筛选SNPs,每个品种选择10 个样本进行混池。测序深度为30,筛选的SNP 位点测序深度大于40。通过与NCBI 上家兔基因组对比,定位HSF1 和HSF4 基因序列,分析SNPs 位点,选择错义突变位点重新设计引物进行基因分型。
1.4 引物设计 根据筛选获得的HSF1 和HSF4 基因突变位置,以及GenBank 中提供的家兔HSF1(基因登录号:NW_003160976.1)和HSF4(基因登录号:NC_013673.1)基因序列,利用Primer Premier 5 设计特异性引物,送至上海生工生物工程股份有限公司合成。由于HSF1 上筛选出的3 个SNPs 位点间距离较短,因此设计1 对引物进行扩增,引物信息见表1。
1.5 PCR 扩增 PCR 反应体系(20 μL):DNA 模板(50 ng/μL)1 μL,上、下游引物(10 μmol/μL)各1 μL,2×Taq Master Mix(1.5 mmol/L)10 μL,ddH2O 7 μL。PCR 反应扩增程序:95℃预变性5 min;95℃变性30 s,退火30 s,72℃延伸1 min,循环35 次;72℃延伸7 min,4℃保存。110 V条件下,用1.5%琼脂糖凝胶电泳检测PCR 扩增产物,电泳30 min。
1.6 测序 利用ABI 3730XL 测序仪采用双脱氧链末端终止法对5 个群体150 个个体的PCR 扩增结果直接进行测序,根据测序结果进行个体基因分型。
1.7 统计分析 基因型频率和基因频率采用基因直接计数法计算;有效等位基因数(Ne)、基因杂合度(He)、基因纯合度(Ho)和多态信息含量(PIC)以及Hardy-Weinberg 平衡状态的检验采用PopGene 软件进行分析;应用Haploview 软件进行连锁不平衡(LD)和单倍型分析;不同群体的基因(型)和单倍型(组合)分布采用SPSS 17.0 进行χ2检验。
2 结 果
2.1 SNPs 的确定 3 个DNA 池重测序结果(表2)发现,HSF1 基因及其上、下游序列有32 个单碱基突变位点和8 个插入/缺失突变位点,其中有2 个单碱基突变及1 个插入/缺失位点位于基因外显子区,导致了错义突变;HSF4 基因及其上、下游序列有11 个单碱基突变位点和3 个插入/缺失突变位点,其中有1 个单碱基突变位于外显子上,引起错义突变。
2.2 PCR 产物测序 个体的PCR 产物测序结果见图1。150 个个体中检测到3 个SNPs 位点,分别为SNP1、SNP2 以及SNP4。SNP3 为假阳性SNP 位点,在群体中未发现多态,均为正常CCT 序列(正常序列为连续4 个CCT,缺失序列为连续3 个CCT),未出现缺失(表2,图1)。
2.3 HSFs 基因的遗传多态性分析 HSFs 基因的遗传多态性分型结果见表3~4。3 个SNPs 位点的基因型和基因分布在5 个群体中均存在显著(P<0.05)或极显著差异(P<0.01),SNP1 在群体的基因型频率与耐热性高低呈现互相对应关系,而SNP2 和SNP4 无明显规律可循。地方品种四川白兔和闽西南黑兔在SNP1 中无多态,均为GG 纯合体,在SNP2 和SNP4 中均呈现中度多态(0.25<PIC<0.5),且为哈- 温平衡群体(P>0.05)。齐兴肉兔和加利福尼亚兔含有全部3 个SNPs,均呈中度多态(0.25<PIC<0.5),除齐兴肉兔在SNP1、SNP2中表现为哈- 温不平衡群体外(P<0.05),其余均表现为哈- 温平衡群体(P>0.05),2 个群体在SNP1 优势基因型为GG,齐兴肉兔的GG 型频率高于加利福尼亚兔。齐卡巨型兔在3 个SNPs 上均表现中度多态性(0.25<PIC<0.5),且为哈- 温平衡群体(P>0.05),其在SNP1 上的优势基因型为GA。

表1 PCR 扩增引物序列
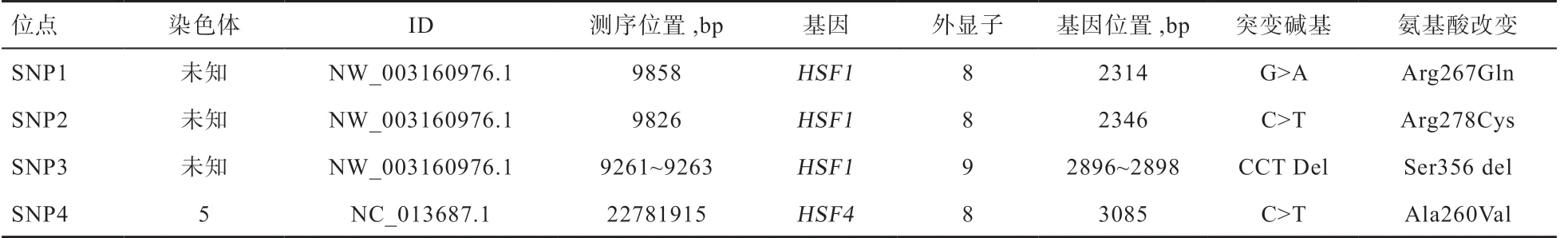
表2 DNA 池重测序结果重要SNPs 信息
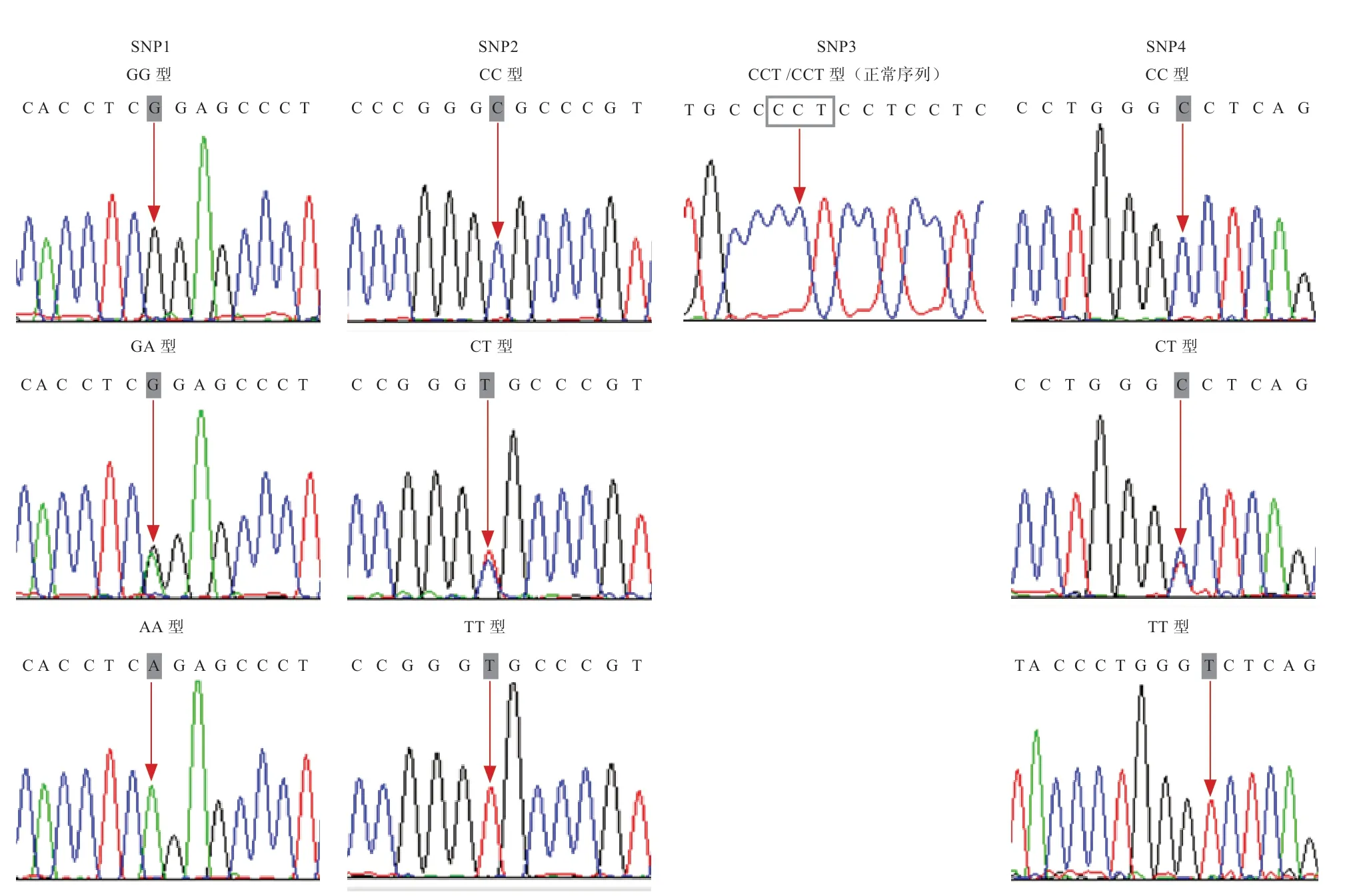
图1 SNPs 测序结果
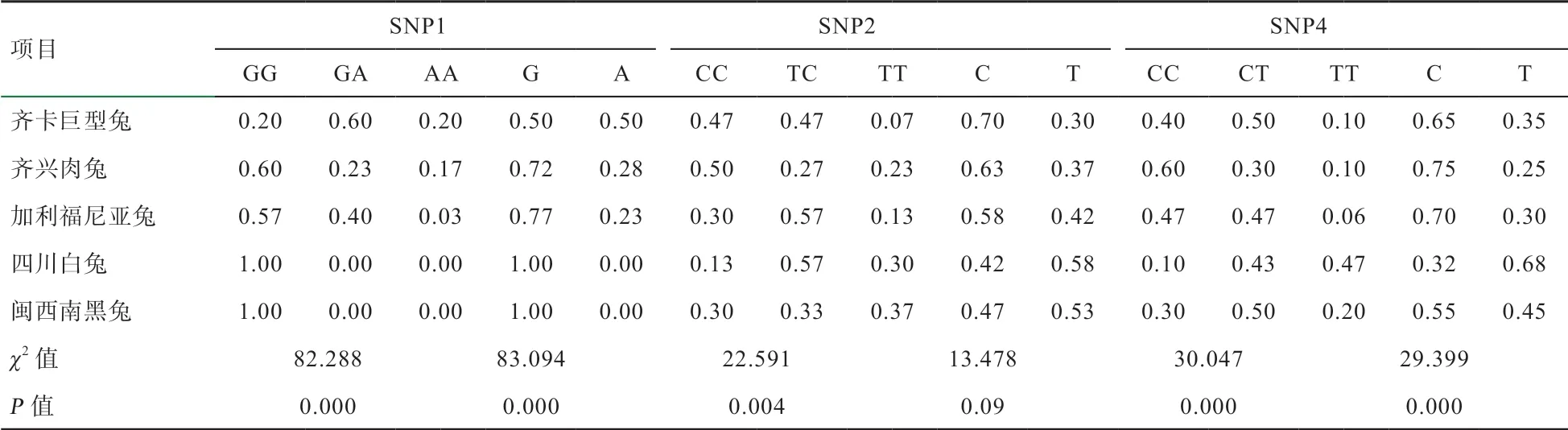
表3 5 个肉兔品种(系)HSFs 基因的基因型频率和基因频率
2.4 连锁不平衡和单倍型分析 利用Haploview 软件进行连锁不平衡分析(表5,图2),结果表明,SNP1 与SNP2 处于完全连锁不平衡(complete LD)(LOD=12.87,D'=1,r2=0.201),2 个多态位点共组成了CG、CA、TG 3 种单倍型;SNP1 与SNP4(LOD=1.23,D'=0.403,r2=0.028)以及SNP2 与SNP4 间(LOD=0.03,D'=0.032,r2=0.001)连锁的可能性极小。
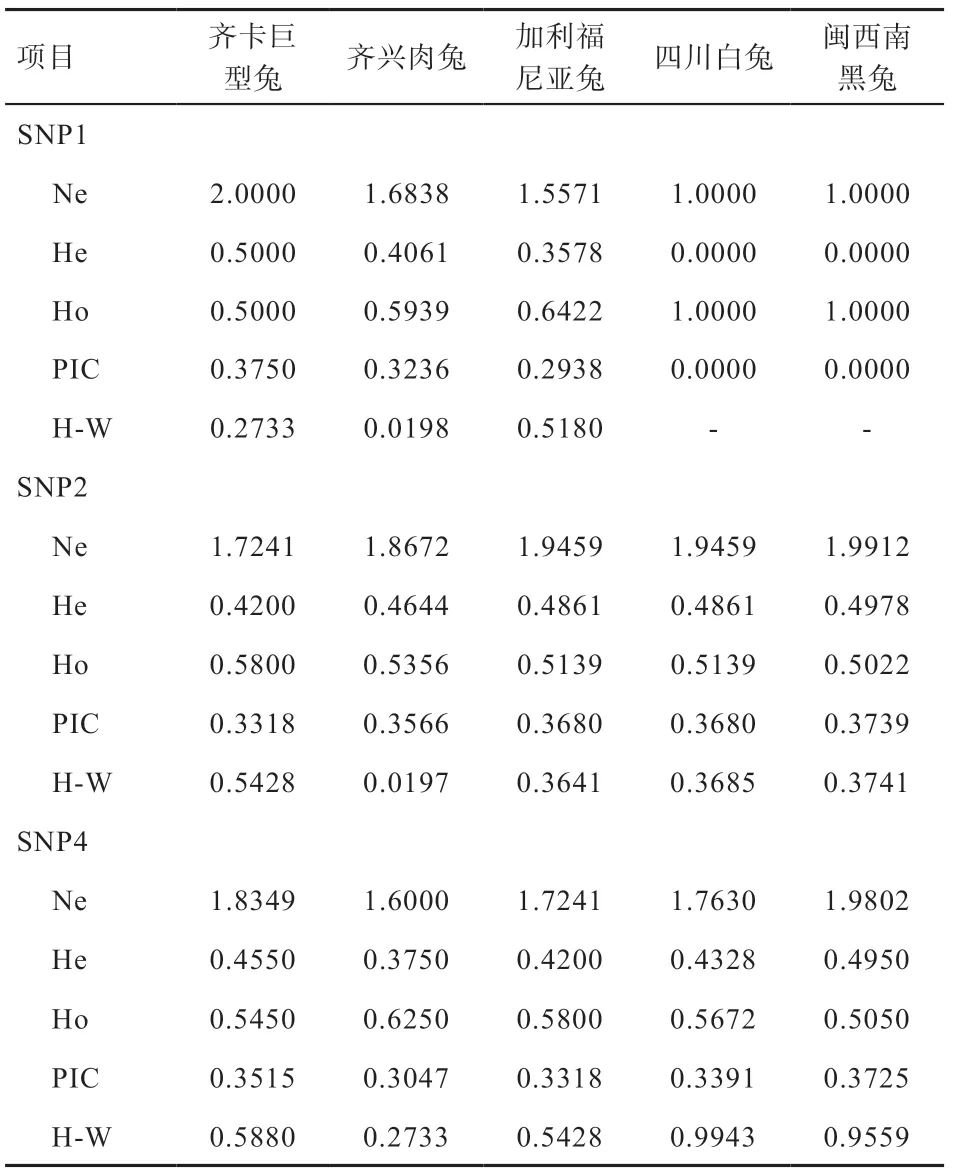
表4 5 个肉兔品种(系)HSFs 基因的多态性分析
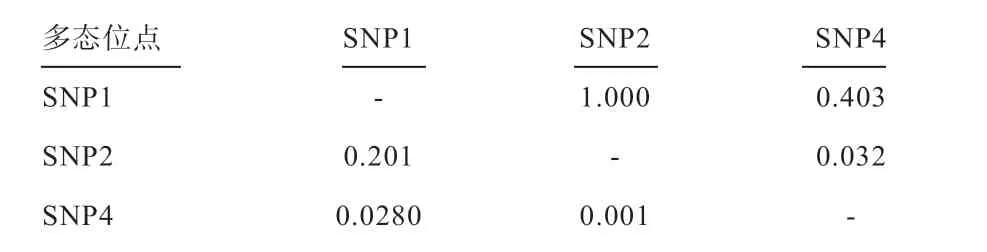
表5 肉兔HSFs 基因的连锁不平衡
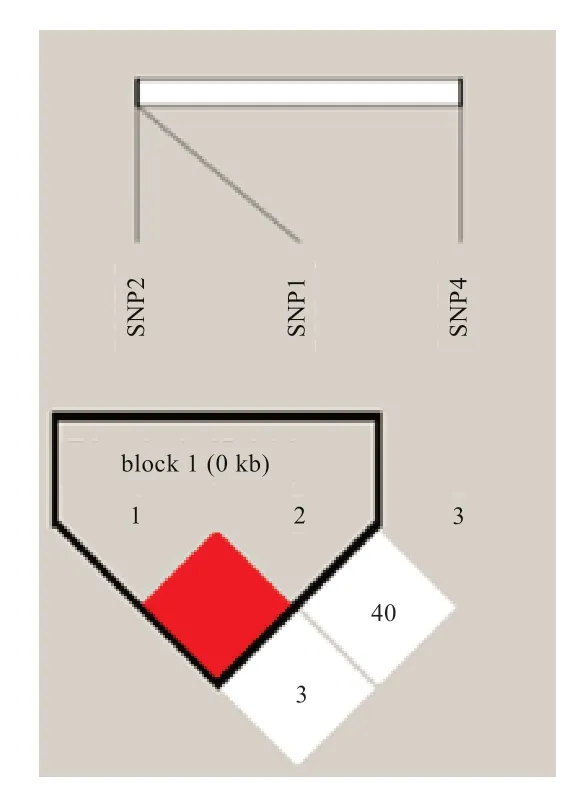
图2 单倍型块图
对5 个肉兔群体构建单倍型,共得到3 种单倍型(表6)。在地方品种四川白兔和闽西南黑兔中只存在H1 和H3 2 种单倍型,优势单倍型均为H3;齐卡巨型兔中包含所有单倍型,其优势单倍型为H2;齐兴肉兔和加利福尼亚兔中包含所有3 种单倍型,其优势单倍型均为H3。
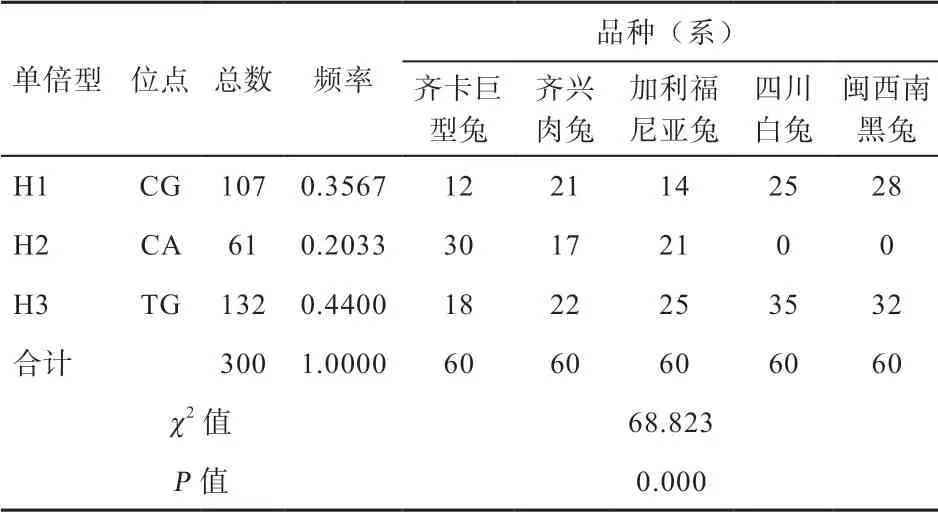
表6 单倍型统计分析
3 讨 论
本研究利用DNA 池重测序技术检测HSF1 和HSF4 的重要SNPs 位点,结果在2 个基因及其上、下游共检出54 个SNPs,选择其中4 个错义突变位点进行一代测序分型,发现其中3 个SNPs 表现出预期多态,而SNP3(缺失突变)并未表现出预期多态,推测该位点可能是一个假阳性SNP。赵冰茹等[14]采用DNA 混池重测序时也发现细毛羊LAMB1 基因10 个错义突变中有3 个假阳性SNPs,推测可能是由于重测序技术的碱基偏好性使得个别SNP 测序时的覆盖率不高,或由于二代测序读长较短,进而造成后续序列拼接、组装等生物信息学分析困难。因此,尽管DNA 池重测序筛查的SNP 位点的准确性较高,但仍需要采用其他方法进行验证。
HSF1 是HSFs 家族中最重要的一员。Hong 等[3]研究发现,在温度不断升高的条件下,哺乳动物细胞通过激活HSF1 使HSFs 表达量增加,充分表明了HSF1在动物的热应激反应中具有重要作用。有鉴于此,国内外多个研究者对HSF1 基因与耐热的关系进行了研究,发现其与牛的耐热性有关。Li 等[6-7]发现,HSF1基因的T909C 和G4693T 突变及其构建的单倍型与中国荷斯坦奶牛的耐热性能有关,4 年后Li 等[15]在3´-UTR 区域新发现1 个SNP 位点,并进一步证实了之前变异位点是重要的耐热性分子标记;王延久等[5]发现,HSF1 基因第3 内含子中存在1 个多态位点,在耐热性能不佳的荷斯坦奶牛、耐热性能好的鲁西黄牛和渤海黑牛的群体中分布差异较大,推测其可能与耐热性有关;Baena 等[4]也发现,HSF1 上的多态位点与安格斯牛的耐热性有关。本研究在家兔HSF1 基因外显子8 上发现的SNP1 和SNP2 均造成了氨基酸突变,且位于该基因4 个可变剪切体的共有序列内,单倍型分析发现它们呈完全连锁状态(D'=1)。5 个群体在SNP1 上的GG 基因型频率大小及耐热性能均表现为四川白兔= 闽西南黑兔>齐兴肉兔>加利福尼亚兔>齐卡巨型兔,说明SNP1 的GG 基因型可能对兔耐热性的形成有积极作用,是重要的耐热性候选分子标记,与上述研究结论一致。SNP2 在5 个群体的优势基因型分布并无规律,因此推测该位点的变异可能与肉兔的耐热性无关。单倍型分析发现,四川白兔、闽西南黑兔以及耐热性次之的齐兴肉兔和加利福尼亚兔的优势单倍型均为H3,而耐热性差的齐卡巨型兔的优势单倍型为H2,与前面单个SNPs多态分析的结果相符,推测单倍型H3 可能是肉兔耐热性的优势单倍型。
HSF4 在HSFs 中发现较晚,Tanabe 等[8]研究结果表明,HSF4 通过可变剪接形成HSF4a 和HSF4b 剪切体,通过抑制和激活热休克蛋白的表达,参与热应激的调节。此外,HSF4 参与晶状体的正常发育,该基因突变是白内障产生的重要原因[16-18]。本研究选择HSF4 作为肉兔耐热的候选基因,发现5 个肉兔群体在HSF4 上基因型分布和基因分布与品种间的耐热性能高低并无规律的对应关系,推测该位点可能与肉兔的耐热性无关。
4 结 论
本研究表明,肉兔HSF1 基因中SNP1 的基因型GG 及单倍型H3 可能与肉兔的耐热性有关。下一步应检测不同基因(单倍)型个体间HSF1 基因表达量是否存在显著差异,以对本实验结果的可靠性进行验证,为在生产上应用该分子标记开展耐热肉兔新品种培育提供依据。
