夹竹桃科驼峰藤物候与繁育特征*
2019-05-27廖浩斌冯璐熊武建蒋谦才张信坚叶矾廖文波
廖浩斌,冯璐,熊武建,蒋谦才,张信坚,叶矾,廖文波
(1. 中山市国有森林资源保护中心,广东 中山528400; 2. 有害生物控制与资源利用国家重点实验室∥ 广东省热带亚热带植物资源重点实验室∥中山大学生命科学学院, 广东 广州 510275)
驼峰藤MerrillanthushainanensisChun et Tsiang 隶属于夹竹桃科Apocynaceae、驼峰藤属MerrillanthusChun et Tsiang[1],为单型属,仅1种,亦为国家Ⅱ级保护植物和第1批国家重点保护野生植物[2],中国特有种[3]。该种最早于1941年由我国著名分类学家陈焕镛教授与蒋英教授在海南白沙县发现并命名[4]。驼峰藤地域分布极其狭窄,仅在海南保亭、万宁、白沙及广东高要鼎湖山等地有过标本采集记录[5-7]。2007年中山市野生植物调查首次发现在中山田心伯公坳有驼峰藤野生分布[8]。2015年6月,在中山市田心森林公园再次看到驼峰藤的踪迹,林业部门立即对驼峰藤进行保护繁殖试验。
目前,关于驼峰藤的相关研究鲜有报道,仅有侯俊等[9]对驼峰藤的组织培养及植株再生进行了相关研究,而对其物候与繁育系统等方面尚未有研究报道。植物的繁育系统是指所有影响后代遗传组成的有性特征的总和,主要包括花的形态特征、花的开放式样、花各部位的寿命、传粉者种类和频率、自交亲和程度和交配系统[10]。近年来,繁育系统的研究越来越多地与传粉生物学研究相结合[11]。本研究以驼峰藤为研究对象,在野外开展生态观察研究,旨在了解该物种的繁育系统及物候特征,为驼峰藤的有效保护提供科学依据。
1 材料与方法
1.1 研究地点和材料
田心森林公园位于北回归线以南(113°28′21″E,22°24′39″N),属南亚热带海洋季风气候,年日照为1 694.4 h,四季分布比较均匀。年平均气温20.3 ℃。雨量充足,年平均降雨量1 858.2 mm,光照充足、水热丰富的自然条件使园区拥有相对原始的自然本底,形成了类型多样的生境。属于丘陵区,地势大致呈东北高西南低,平地较少、山地较多,最高峰海拔为383.7 m。本研究地点位于中山市田心森林公园内的3个驼峰藤自然居群,植被群落组成以驼峰藤为特征种,主要伴生种有艳山姜Alpiniazerumbet、鸭跖草Commelinacommunis、地桃花Urenalobata、水东哥Saurauiatristyla、土沉香Aquilariasinensis、粉单竹Bambusachungii、粗叶榕Ficushirta、红枝蒲桃Syzygiumrehderianum、海芋Alocasiamacrorrhiza、猴耳环Pithecellobiumclypearia、假苹婆Sterculialanceolata、鲫鱼胆Maesaperlarius和娃儿藤Tylophoraovata等。土壤以沙壤土和红壤土为主。
1.2 研究方法
1.2.1 物候周期的观察 驼峰藤为常绿木质藤本,2017年3月开始,次年3月为一周期进行持续观测,观察和统计物候周期的数据。
1.2.2 花部特征观察 在居群内,分别于 2017 年3月和4月挂牌标记。① 不同植株即将开花的花蕾30朵,连续多天定位观察并记录花朵个体的开放进程,直至花瓣脱落。随机选取当天开放的花30朵,用游标卡尺测量萼片、花瓣、副花冠、合蕊冠、子房的大小。② 在体式解剖镜下观察花药裂口、花粉块等花部微观结构。
1.2.3 柱头可授性的检测 采用联苯胺-过氧化氢法检验柱头的可授性[12]。取开花后不同天数的花各5朵,在解剖镜下小心将子房及柱头剥离出来,在凹面载玻片中加入体积比为w=1%联苯胺∶φ=3% 过氧化氢∶水 = 4∶11∶22 的反应液,将子房及柱头浸入其中。若柱头具可授性则周围反应液呈现蓝色并有大量气泡出现,根据气泡的多少和大小判断柱头可授性的相对强弱。
1.2.4 杂交指数的估算 按照 Dafni的标准,通过花序直径、性器官空间位置与成熟情况等评判繁育系统类型[12]。OCI = 0 为闭花受精;OCI = 1 为专性自交;OCI = 2 为兼性自交;OCI = 3 为自交亲和,有时需要传粉者;OCI = 4 为异交为主,部分自交亲和,且需要传粉者。
1.2.5 访花昆虫的观察 分别在 2017 年 4 月 23 日至 29 日,5 月 5 日至 11 日,每次连续 7 d,从 8:00~19:00 初步观察并记录所有驼峰藤的访花昆虫的访花行为、访问时间、单花停留时间等。观察过程中,不定时地网捕访花昆虫,检查它们是否携带花粉块,并制作标本,进行种类鉴定。
1.2.6 自交与对照实验 2017年4月23日开始进行实验,4月26日,4月29日,5月10日连续进行观察,得到结果。
对照组:每个植株随机选择 30个未开放的花序进行挂牌,不套袋,不去雄,检测自然条件下的结实情况。
自交组:每个植株选择 30个花序,在开花前(花蕾期)套袋,不去雄,花开后分别对其授自花花药的花粉,检测是否存在自交现象。
套袋组:每个植株选择 30个花序,在开花前完全套袋,不去雄,检测是否存在自交现象。
2 结果与分析
2.1 物候周期
2.1.1 中山市驼峰藤的周年物候周期 主要包括展梢期、开花期、盛花期、终花期、幼果期、果实成熟期、落果期。由表1可见,展梢期集中在一、二月份。驼峰藤的开花期包括花蕾期、展花期和盛花期,花蕾期出现在3月初期,在4月开始展花,每一朵花逐渐开放,四月中旬就到了盛花期,一直持续到5月上旬,在4月下旬,就开始结果。终花期从5月下旬开始,从4月下旬开始与结果期有一些重合。 幼果期从4月下旬开始,每个花序一般会结1~2个果,果逐渐增大变成熟,一直持续到6月上旬。6月底,果实基本成熟,由于果实很大,花序果期,果序仅存一枚果,其他败育。一直持续到9月,种子逐渐成熟,果皮开裂,向腹面折起,种子飞出。种子成熟后,基本进入落叶期,落叶主要是失去生理机能的老化叶片。10月份果实已经成熟,种子飞出,落叶明显。
2.1.2 花期物候 开花期从3月份开始,盛花期持续2个月,6月下旬,果实成熟,成熟期持续四个月。结果期会与当地的雨季有重合,频繁的下雨会严重影响果实的存活。在驼峰藤野生居群中,花期为3月至5月,初花期3月中旬,盛花期4月23日,末花期5月中旬。
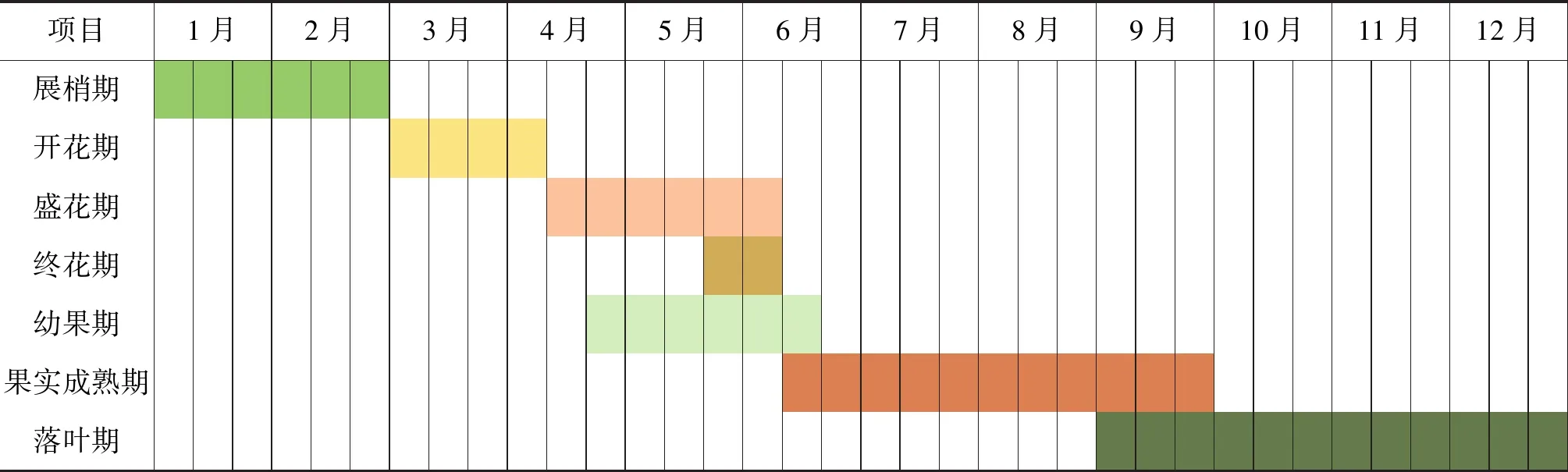
表1 驼峰藤物候周期Table 1 Phenological phase ofMerrillanthus hainanensis

表2 驼峰藤种群开花进程Table 2 Flowering process ofMerrillanthus hainanensispopulation
2.1.3 单花开花进程 单花花期为6~8 d、花序花期为16~20 d。伞形花序,自下而上依次开放。在单花开放过程中,一片花瓣先打开,随后其余4 片花瓣依次顺序开放,一般在上午10:30 左右开放,花冠呈绿色,访花昆虫开始到访;到第3天下午时,花瓣开始向外卷曲,颜色由绿色逐渐变成淡黄色,访花昆虫种类与数量均有所增加;到第6天花瓣开始卷曲萎蔫变成棕黄色,访花者数量明显减少;一般到第7天整朵花脱落。受精后的花不会立即脱落,其花期比未受精的短,干枯后紧包着幼果(图1)。
2.2 花器官的发育特征
驼峰藤的花序为聚伞形花序,具有较长的总花梗,花梗长(15.90±2.03) mm。花被由5个花萼和5个花瓣组成。花萼裂片卵圆形。花冠绿色,辐状,直径(19.19±1.55) mm,裂片卵圆形,先端急尖。副花冠膜质,从合蕊冠的基部长出并包围着合蕊冠。合蕊冠呈五角形,高约(3.33±0.51) mm。雌蕊由2枚离生心皮组成,子房上位,长(2.68±0.13) mm,宽(1.24±0.09) mm。雄蕊5个,花药2室,每花药有花粉块2个,每药室藏1个,花粉块卵圆状,下垂,相邻2药室中的2个花粉块分别通过2个花粉块柄固定在1个棕色的着粉腺上。着粉腺位于花药裂口上端,花药裂口通向柱头腔。子房内具多个胚珠(图2)。

图1 驼峰藤的开花进程Fig.1 Picture of the flowering process ofMerrillanthus hainanensisA.花蕾期;B. 盛开期; C. 开花后期;D. 谢花期

图2 驼峰藤的花期果期物候Fig.2 Flowering phenology ofMerrillanthus hainanensisA.花蕾期;B .完全打开的花:正面;C.完全打开的花:背面;D.花末期;E.双生果幼期; F.双生果成熟期; G.单生果;H. 果剖开:幼期;I.果剖开:成熟期
2.3 柱头可授性
用联苯胺—过氧化氢法检验柱头的可授性,可以看出驼峰藤在整个花期都具有柱头可授性,从开花当天到第3天均具有强可授性,随后逐渐降低(表3)。

表3 不同花期驼峰藤柱头可授性测定1)Table 3 Stigma receptivity ofMerrillanthus hainanensisin different flowering dates
1)“-”柱头不具可授性;“+”柱头具可授性;“++”柱头可授性较强;“+++”柱头可授性最强
2.4 杂交指数(OCI)的估算
根据 Dafni的标准[12],驼峰藤花序和花朵直径均大于6 mm,记为3;驼峰藤为两性花,花粉聚集在一起形成花粉块,不能散落到柱头,开花时花药低于柱头且存在隔离,两者无法接触,记为1;花粉和柱头同步成熟,记为0。据此,驼峰藤的杂交指数为4,其繁育系统为异交为主,部分自交亲和,需要传粉者(表4)。
2.5 访花昆虫及其访花行为
访花昆虫种类:主要有切叶蜂Megachilesp.、中华蜜蜂Apiscerana和台湾马蜂Polistesformosanus,还有离斑棉红蝽Dysdercuscingulatus、大绢斑蝶Paranticasita、宽边黄粉蝶Euremahecabe等。
访花具体行为:昆虫在花周围盘旋,借助足部特殊结构将花粉块移出并将花粉块通过花药裂口插入柱头腔中以完成授粉。访花时间:上午10点左右。阳光充足。
2.6 自交与对照实验结果
实验结果见表5,实验结果见表5,结果表明:套袋处理的均未结果实,说明驼峰藤不存在自花授粉机制,也无法进行无融合生殖,在自然情况下必须依靠传粉媒介才能完成受精过程。驼峰藤自然坐果率较低,为 13.5%。花序坐果率(指标记的花序是否结果)较高,达到90% 以上,每个花序最终结果一个,极少有双生果(其中一个子房不育),每个果实含有种子约 100 颗。

表4 驼峰藤杂交指数测定Table 4 Out-crossing index ofMerrillanthus hainanensis

表5 驼峰藤套袋实验处理结果Table 5 Test results of emasculation ofMerrillanthus hainanensis
3 讨 论
3.1 驼峰藤的物候分析
该试验一定程度上填补了驼峰藤物候研究的空白,同时应该指出的是物候研究需要一定的时间积累,时间越久得到的数据越是弥足珍贵。另外,植物生活在复杂多变的环境中,植物的生长发育过程在很大程度上也受其小环境的影响,如海拔 、坡向 、地形等因素。该试验只是对中山市田心森林公园这一个群落点的驼峰藤进行观察记录,还有一定的局限性。
3.2 繁育系统分析
濒危植物的生殖力、存活力、适应力低下等内在因素是其走向濒危的根本原因[13]。近年来,珍稀濒危植物开花生物学及繁育系统的研究受到重视[14]。驼峰藤的花粉粒聚合成花粉块,不易被风吹走,使其在自然条件下,需要有传粉者参与,繁殖以异交为主。驼峰藤的花粉活力、花粉数量、柱头可授性等生殖构件及开花过程并不是导致其濒危的原因,但与其开花期结果期与中山市雨季重合,驼峰藤本身果实比较重,果柄却比较细,有很大的影响。驼峰藤属与萝藦科娃儿藤属很相似,因本属花冠裂片的端部互相粘合,花粉块柄先横平,次上升,后斜下,蓇葖仅一个成熟等与娃儿藤属很不同。
