Sm a ll GTPases:Stru ctu re,bio logica l fun ction an d its in teraction w ith nanopa rticles
2019-05-13SiyangSongenshuCongShurongZhouYujieShieningDaiHuaZhangXueqingWangBingHeQiangZhang
Siyang Song,W enshu Cong,Shurong Zhou ,Yujie Shi,W ening Dai,Hua Zhang,Xueqing Wang,Bing He,Qiang Zhang,∗
a Shenyang Pharm aceuticalUniversity,No.103,Wenhua Road,Shenyang 110016,China
b Peking University,No.38,Xueyuan Road,Beijing 100191,China∗Co rrespon d ing au thor.Shenyang Pharm aceu tica l Un iversity,No.103,W enhua Road,Shenyang 110016,Ch ina.Tel.:+86-10-82802791.
Keyw ords:Sm a ll GTPase Nanoparticles Ras Rab Rho
A B S T R A C TSm a ll GTPase is a kin d o f GTP-bin d ing p rotein com m on ly foun d in eu karyo tic cells.It p lays an im portan t ro le in cytoskeletal reorgan ization,cellpolarity,cellcycle p rogression,gene exp ression an d m any other sign if ican t even ts in cells,such as the in teraction w ith foreign particles.Therefore,it is o f great scien tif ic sign if ican ce to understan d the bio logica l p roperties o f sm a ll GTPases asw e llas the GTPase-nano in terp lay,sin cem o re and m ore nanom ed icine are supposed to be used in biom ed ica l f ield.How ever,there is no review in th is aspect.Th is review sum m arizes the sm a ll GTPases in term s o f the stru ctu re,bio logica l fun ction and its in teraction w ith nanoparticles.W e brie f ly in troduced the various nanoparticles such as go ld/silver nanoparticles,SW CNT,po lym ericm icelles and other nano delivery system s that in teracted w ith d ifferen t GTPases.These cu rren t nanoparticles exh ibited d ifferen t pharm aco logical effectm odes and various target design con cep ts in the sm a ll GTPases study.Th is w ill help to e lucidate the con clusion that the therapeu tic strategy targeting sm all GTPases m igh t be a new research d irection.It is believed that the in-dep th study on the fun ctional m echan ism o f GTPases can p rovide in sigh ts fo r the design an d study o f nanom ed icines.
1. In trodu ction
Sm a ll GTPase fam ily is a GTP-bind ing p rotein fam ily w h ich can be com m on ly found in eukaryotic cells[1].It is a kind o f GTPases can ach ieve m u tua l tran sfo rm ation betw een GTP and GDP in cy top lasm[2].These sm a llGTPases act asm o lecular sw itches in cells,a ffecting a lm ost a ll cellu lar p rocesses[3].The m ost p rom inen t m em ber o f sm a ll GTPase fam ily is the Ras GTPase,thus the fam ily is also called the Ras superfam ily[4,5].
2. Stru ctu re an d action m echan ism o f sm a ll GTPases
2.1. Structure ofsm allGTPases
The analysis o f the sm allGTPase p rotein crystal structu re ind icates that the GTPbind ing dom ain o f th is type o f p ro tein can be subd ivided in to f ive re latively conserved m otifs G1-G5.G1 m otif(I)is a pu rine nuc leotide bin d ing signal;G2m otif(E)is in one o f tw o segm en ts that red irects w ith GDP o r GTP bind ing fun ction and p rovidesm a jo r com ponen t o f the e ffecto r bin ding su rface;G3m o tif(II)is re lated to the bind ing o f nu cleo tiderelated Mg2+;G4 m otif(III)brings the hyd rogen bon d in contact w ith the guan ine ring;G5 m o tif(Ⅳ)m akes ind irect associations w ith the guan ine nu cleo tide[6-8].Taken together,these elem en ts constitu te the conserved∼20 kDa dom ain and shared by all Ras superfam ily p roteins.
2.2. Action m echanism ofsm allGTPases
While participating in physio logica l activity,the m o lecu lar stru ctu re o f sm all GTPases p resen t tw o form s w h ich support m u tua l transfo rm ation,GTP-bind ing activated state and GDPbind ing non-activated state,w h ich can a lso be ca lled as“ON”state and“OFF”state,respectively[3].
The hyd rolysis of GTP to GDP is m ed iated by GTPaseactivated p ro tein s(GAPs),and the exchange o f GDP to GTP is m ed iated by guan ine nuc leotide exchange facto rs(GEFs)[9].GEFs and GAPs coex ist in m ost ce lls,in creasing the d iversity o f signa ls that regu late sm a ll GTPases activity[8].Guan ine nucleo tide d issociation inh ibito rs(GDIs)are con trary to exchange facto rs.GDIspecif ica lly binds GDP-bound GTPase and inh ibits GDP release(Fig.1B)[10].
3. Classifica tion ofsm a ll GTPase protein
Ras GTP enzym e w as in itia lly found to be a m u tation o f GTPase in various can cers.As tim e goes by,m any GTP enzym es are found to have the sim ilar3D structu rew ith Ras.At p resen t,the fam ily has over 150 m em bers m ak ing it a superfam ily.On the FASEB Sum m er Consu ltative Con feren ce in 1992,the com p rehensive nam ing p rin cip le of Ras-like GTPase w as estab lished,d ivid ing the sm a ll G p rotein in to f ive sub fam ilies o f Ras,Rho,Rab,A rf/Sa r and Ran acco rd ing to the sequen ce,stru ctu re and fun ction.The sorting criterion is now w ide ly accep ted.Recen tly,Rad,Rap,Rheb,Ritan d M iro are a lso in cluded in to the superfam ily[11].
Ex tensive researches on each fam ily o f sm a ll GTP enzym e fam ily in term o f can cer on ly focused on how these p ro tein fam ilies con tro l the no carcinogen ic p rocess(su ch as im m un ity and in f lam m a tion)at the beginn ing.The fun ctions o f these GTP enzym es are greatly overlapped.Ras takes charge o f cell p roliferation,Rho is used for cellm orphology,Ran is used fo r nuclear tran spo rt,and Rab and A rf a re used fo r vesicle tra ff ick ing[8].
3.1. Ras subfam ily
Ras superfam ily is the f irstand them ostd iversif ied sub fam ily.It p lays a la rge ro le in m ed iating im m un ity an d in f lam m ation[7,8].The Ras sub fam ily p ro tein is a lm ost the generic com ponen t in signaling pathw ay o feukaryotes(includ ing vertebrate,invertebrate and yeast),w h ich p lays a key ro le in the developm en t,p ro liferation,d ifferen tiation and su rviva l o f eu caryon.In add ition to the w ide ly exp ressed m a jor Ras iso fo rm s,the h igh ly conserved H-,K-and N-Ras exh ibit diverse biologica lp roperties[12,13].H-ras,K-ras and N-ras genes are located on ch rom osom es 11,12 and 1,respective ly.K-ras gene is a lso know n as the p21 gene because it encodes a 21 kDa Ras p rotein.K-Ras has the la rgest im pact on hum an can cer.
3.2. Rho subfam ily
Rho GTPase fam ily con tains w ell-developed Rac1,RhoA and Cdc42 w h ich m odu late the form ation o f cytoskeleton,cell polarity,ce ll cycle p rogression and signa l netw ork o f gene exp ression[14-16].RhoA,Rac1 and Cd c42 possess them ost obviousm em bers[17,18].They serve as them o lecu lar sw itches in separate or co rre lative signa ling pathw ays[19].These signa ling pathw ays link the p lasm a recep to rs to activate cy toske leton reo rgan iza tion an d the subsequen t bio logica leffect.Cd c42 and Rho can be com m on ly found in yeast and m ost m amm a lian ce lls,w h ile Rac can on ly be observed in the latter[20].
Actua lly,the th ree kinds o f p ro tein p lay the sam e sign ifican t ro le in m odu lating cell po larity,genetic transcrip tion,cellcycle p rogression,m icrotubu le kinetics,vesicle traff icking,ex trace llu larm atrix(ECM)rem odeling and various enzym atic activities(using NADPH ox idase activity to generate the reactive oxygen,ROS)[21].
3.3. Rab subfam ily
Rab sub fam ily is the largest sub fam ily in Sm a llGTPase Pro tein fam ily[1].Am id the 93 Sm a ll GTPases fam ilym em bers o f A rabidopsis,57 belong to Rab subfam ily.Rab subfam ilym odu lates the vesicle tra ff ick ing and p rotein transpo rt o f eukaryocy te by regu lating com p lex vesicle tra ff icking andm icrotubu le system activities.
At p resen t,there are 60 kinds o f Rab p ro tein s(in clud ing isom ers)w h ich p lay an im po rtan t ro le in p rom o ting and m odu lating the berth and in tegration o f tra ff icking vesicle(Fig.2).Each organelle has at least one kind o f Rab p rotein and d ifferen tm em b ranes have d ifferen t Rab p ro teins.
3.4. Arfsubfam ily
Sim ilarw ith Rab p rotein,the ADP-ribosy lation factor(A rf)subfam ily p rotein is the m ajor regu latory factor o f the biosynthesis o f tra ff ick ing vesicle in cells[22].Differen t from Rab p ro tein w h ich w o rks in sing le step o f m em brane transportation,Arf p rotein can w ork in various steps.For exam p le,Arf can con tro l(1)form ation o f coat p rotein I(COPI)-coated vesicle supporting an tiporter betw een Go lgiand ER;(2)form ation o f c lath rin/adap tin 1(AP1)com pound-re lated vesic le in trans-Golginetw ork(TGN)and imm atu re vesic le and(3)form ation o f en dosom e con tain ing AP3[6].
3.5. Ran fam ily
Ran is a kind o f sm all GTPases w ith h igher exp ressive abundan ce in eukaryocy te[23].It can m odu late the fo rm ation o f cell sp in d le apparatus,cell cycle p rogression,structu re and fun ction o f karyotheca,m ateria l transpo rt o f nu cleop lasm,cell redox reaction and RNA nuclear expo rt,RNA syn thesis and p rocessing,etc.[24].
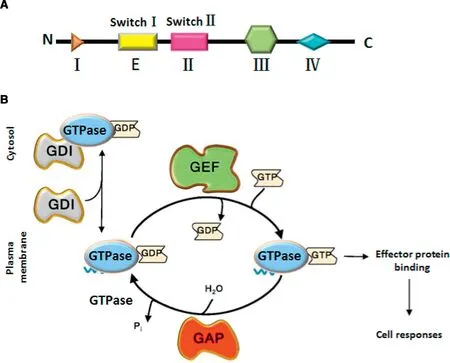
Fig.1-(A)Schem atic d iag ram o f the sm a ll GTPase p ro tein sequen ce.(B)Regu lation o f sm a ll GTPase activ ity.GTPases cy cle betw een their GTP-boun d active an d GDP-boun d inactive fo rm.They are activated by guan ine-nu cleotide exchange facto rs(GEFs),w h ich rem ove GDP,thus enab ling ex cess ce llu la r GTP to bind.The bin d ing o f active GTPase to dow n stream e ffecto r p ro tein s e licits ce ll respon ses.GTPase-activating p ro tein s(GAPs),w h ich in crease the GTPase activ ity,a re the o ff-sw itch.Inactive Rac is sequestered in the cy toso l by guan ine-nucleotide d issociation inh ibitors(GDIs).Rep roduced w ith perm ission from[78].Copy righ t 2018 John W iley&Son s,In c.
Ran participates in the m odu lation o f karyop lasm transpo rt o f m am m a l and yeast.Many nuc leopo rins o f m am m a l and yeast con tain Ran bind ing dom ain(RBD).RBD,Ran and nu clear im po rt factor fo rm the tripo lym er.A fter nu clear expo rt,w hen the recep to r com pound binds w ith Ran-GTP,the substrate w ill be re leased.The rem ain ing recep tors and Ran-GTP d im er w ill retu rn to cytop lasm and re-participate in the transport o f nu clear im po rt factors.Ran a lso regu lates the m o lecu le nu clear expo rt.The nuc lea r expo rt recep to r an d its substrate bind w ith Ran-GTP to form the com pound.A fter nuclear export,Ran-GTPw illbe hyd rolyzed in to Ran-GDP to separate recep to r and substra te[25].
4. Fun ction o f sm a ll GTPase an d its ro le in nano de livery system
Sm all GTPase can regu late various vitalm ovem en ts of cells,in clud ing cell grow th,d ifferen tiation,ce ll m ovem en t,lip id vesicle tra ff ick ing,etc.[26,27].In term o f fun ction,Ras subfam ily regu lates the gene exp ression;Rho sub fam ily regulates cy toskeleton reorgan ization,cellw a ll syn thesis,cell cycle p rogression,and MAPkinase signa l transduction;Rab subfam ily an d Sar/A rf sub fam ily regu late the vesicle tra ff ick ing and clath rin form ation;Ran sub fam ily regu lates karyop lasm transport,m icrotubu le fo rm ation,fo rm ation o f m ito tic sp ind le apparatus and the assem b ly o f karyo theca a fter cell d ivision.At p resen t,m o re andm o re researches suggest the im po rtan t ro le o f sm a llGTPase in the nanopartic le transpo rt[28-31].
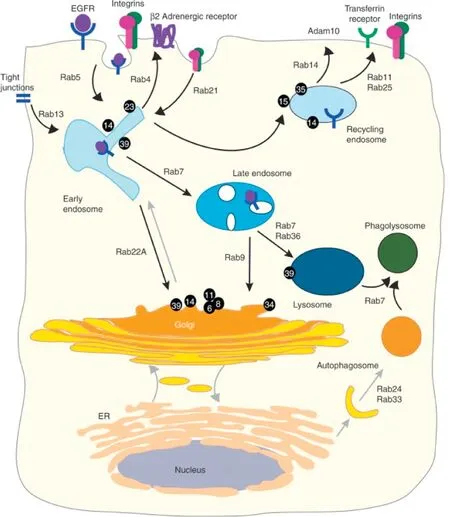
Fig.2-Overv iew o f Rab GTPases on the en docy tic pathw ay.Rab GTPases fun ction in in terna liza tion an d tran spo rt todeg radation,as w e ll as recycling to the p lasm a m em b rane an d the Go lgi.The num bers in the f igu re ind icate o ther k in ds o f Rab GTPases,fo r exam p le,14 rep resen ted o f Rab14.Rep rodu ced w ith perm ission from[40].Copy righ t 2014 Co ld Sp ringHarbor Laborato ry Press.
4.1. Ras
Ras is a GTP-bind ing p ro tein invo lved in m em b rane ru ff ling,p inocy tosis and the form ation o f stress f ibres.The ras on cogenes exh ibit transfo rm ing p roperties due to single poin tm utation in the sequen ce cod ing fo r the active site o f the p21 p ro tein[32-34].These m u tations lead to changes in cellu lar p ro liferation and indu ce tum o rigen ic p roperties.In an earlier study,an tisense o ligonu cleo tides d irected to poin t m utations(G→U)in codon 12 o f Ha-ras m RNA selectively inh ibited the p ro liferation o f cells exp ressing poin t m u tan t HRas genes[35].W hen it w as absorbed by PIHCA NPs,the sam e effect cou ld be ach ieved w hen the con cen tration w as 100 tim es low er than that o f the free o ligonuc leotide.NPs p ro tected o ligonuc leo tides from rap id in tracellu lar degradation,w h ich led to considerab ly h igher in tracellu lar con centrations o f in tact o ligonuc leotides.As a resu lt,they m arked ly inh ibited H-Ras depen den t tum o r grow th in nude m ice a fter subcu taneous in jection.These experim en ts show ed tha t using an tisense oligonu cleotides to inh ibit ras oncogenes can b lock tum or developm en t even though activation o f ras on cogen ic cou ld be seen in the early stage o f tum o r p rogression.Th is w ork w as very new in 1994 because RNAi em erged in 1990.In another sim ilar study,researchers also silenced the ras gene.They en trapped the an tisen se siRNA o f Ras inside a GSH-responsive nano tran spo rter to be delivered specif ica lly to a tum o r cell.Therefore,it knocked dow n severa l d ifferentia lly exp ressed genes that a re regu lated by the Ras activation pathw ay[36].Effo rts to perfo rm in vivo siRNA delivery w ith a p roperly designed GSH-responsive nanocarrier m igh t lead to transform ed targeted can cer treatm en ts.
In recen t stud ies,Ras is a lso used to d iagnostics can cer.Researchers degraded hum an genom ic DNA w as by son ication and inserted the dye f luo rescen t quen ch ing quan tum dots(QDs)after target hybrid ization in the m icro f luidic ch ip.A fter the K-ras on cogene hyb rid ized w ith the p robe DNA in the channel,the in terca lating dye(TOTO-3)cou ld quen ch the f luorescen ce of the QD to detect K-ras gene[37].Wang et al.detected pan creatic can cer patien ts’feces o f K-ras gene m u tations bym agnetic nanop robe show ed that nanoparticles w ere ab le to detect K-rasm u tations in d ifferen t stages o f pan creatic can cer,w ith com parable sen sitivity and specif icity to CA19-9 exam ination for d ifferen tiating pan creatic can cer[38].These m ethods are bo th good exam p les to detect on cogenes and have good clin ica l app lication p rospect.
There are a lso stud ies on o ther aspects o f Ras.Ro tb lat et a l.described a h itherto unknow n cellu lar signa ling particle,rasosom e[39].Its size w as nanom eter and m oved rap id ly in an ATP-indepen den t random m otion in the cytosol and carried m u ltip le cop ies o f pa lm itoy lated Ras p ro teins.A lthough it is an endogenous substan ce,the au tho r still regards it as nanopa rticle.
4.2. Rab
Nearly 3/4 o f abou t 70 hum an Rab-GTPases are invo lved in endocy tic tra ff icking[40].En dosom a lm em b rane transpo rt pathw ays exh ibit a sign if ican t p lasticity due to recep tor signaling and Rab-GTPase-regu lated sca ffo lds.The ro les o f Rab-GTPases are d iscussed w idely,in clud ing(1)com partm en ta lizing the endocy tic pathw ay in to early,recycling,late,and lysosom a l rou tes;(2)coord inating ind ividual transport steps from vesicle budd ing to fusion(3)effecto r in teraction;and(4)signa ling cascades[14,41-44].
Becker et al.investigated a strategy in a study,using a sm a ll GTPase o f the Rab fam ily on DNA-Go ld nanoparticles in virtue o f a site-specif ica lly attached po ly(ethy lene glyco l)lin ker and th io l p lace-exchange reaction[45].Rab p ro teins act as the cen tral regu lators o f vesicle traff icking and participa te in vesicu lar budd ing,targeting,and fusion[42].Rab6 w as origina lly iden tif ied as a key p layer o fm icro tubu le-dependen t retrograde tra ff ic,con tro lling vesicle tra ff icking from early endosom es to the TGN th rough the Go lgi apparatus,and from the Go lgi to the endop lasm atic reticu lum[46].It appears to participate in a com p lex netw o rk o f p ro tein-p ro tein in teraction s.
Ano ther study app lied the rap id m u ltico lo r 3D live ce ll con foca l f luo rescen ce m icroscopy,and adop ted transien t over exp ression o f sm all GTPases m arking various en docy tic m em b ranes,ind icating the k inetics o f nanopartic le tra ff ick ing th rough early endosom es to late endosom es and lysosom es[47].It is show n that,a fter the in terna liza tion,the 40 nm po lysty rene nanopartic les f irst pass th rough an early en dosom e in term ed iately decorated w ith Rab5,bu t they are rap id ly tran sferred to late endosom es and u ltim ately lysosom es labeled w ith Rab9 and Rab7,respectively.100 nm larger nanoparticles a lso reach acid ic Rab9-and Rab7-positive com partm en ts desp ite a slow er rate than the sm a ller 40 nm nanoparticles.Th is study revea led that re latively few nanoparticles can access endocytic recycling pathw ays,considering the lack o f sign if ican t co loca lization w ith Rab11.It a lso dem onstrated that th is quan titative app roach cou ld effectively detect rare even ts in nanoparticle traff icking,specif ica lly nanoparticles in Rab1A-labeled structu res,thereby revea ling the ex tensive in trace llu la r in teractions betw een nanoparticles and in trace llu lar environm en t.
4.3. Rho
4.3.1. Cdc42
Cdc42 signaling can effectively con tro l cell p ro liferation,cell po larity,su rviva l an d invasion[48-50].Knockdow n o f Cd c42 led to a 46%decrease o f the in terna lization o f PEGy lated silicacoa ted iron ox ide nanoparticlesw ith in 24 h in cubation in Hela cells,p roving the sign if icance of Cdc42 in th is endocytosis m echan ism[51].How ever,knockdow n o f Dynam in 2,Flotillin-1,Clath rin and PIP5Kαcaused no o r on ly m ino r e ffects.En docy tosis o f iron ox ide nanoparticles in HeLa cells w as m ain ly m ed iated by Caveo lin-1 and Cd c42.Cdc42 cou ld m ed iate the in terna liza tion o f nanoparticles w as re ferred in tw o review s,bu t there is no actua l exam p le[28,29].It is show n here for the f irst tim e Cdc42 is invo lved in endocytosis of silica-coated iron ox ide nanopa rticles in HeLa ce lls in vitro.W e expect thatm ore nanopa rticles cou ld be con f irm ed w hether they are related to Cd c42-m ed iated endocy tosis.
4.3.2. Rac1
As the m ain regu lator in actin cytoskeleton reorgan ization,Rac1 can a ffect endocy tosis and tra ff icking,ce ll cycle p rogression,adhesion and m igration[52,53].Im po rtan tly,Rac1 con tro ls lam e llipod ia fo rm ation and m em b rane ru ff les a fter being stim u lated by ex tracellu lar ligands such as ep iderm al grow th facto r(EGF),p latelet-derived grow th facto r(PDGF)o r insu lin[54].By using pharm aco logica l inh ibito rs and in virtue o f genetic app roaches,SW CNT is verif ied to be endocy tosed th rough Rac1-GTPase m ed iated m acrop inocy tosis in no rm a l endo thelia l ce lls[55].
Berry et a l.syn thesizedm agnetic nanoparticles and derivatized w ith dextran(DD),fo llow ed by com paring w ith sim ilar underivatized p lain particles[56].Desp ite o f the up take o f bo th the un coated and the DD-derivatized pa rticles in to the cell,the derivatized particles can indu ce a lterations in cell behavio r an d m o rpho logy com pared w ith the p lain particles,suggesting that cell response is dependen t on the particles coating.The DD nanopartic les led to elevated c lath rin localized to the ring-like actin structu res due to changes in actin po lym erization in response to activation o f the sm a llGTPases Rac1 and Cd c42.
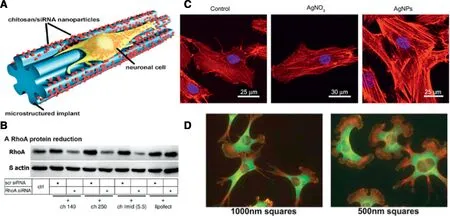
Fig.3-(A)Schem e o f nanopa rticles bio fun ctiona lized nerve im p lan ts,a re taken up by ce lls an d enab le neu rite ou t g row th.(B)Neu rona l respon se to nanopa rticles by RhoA p ro tein redu ction[66].Copy righ t(2010),Am erican Chem ica l Society.(C)Stain ing o f f ilam en tou s actin in the AgNP-treated w as stronger than that o f the con tro l an d AgNO3-treated cells[68].Copy righ t(2014),Dovep ress Ltd.(D)Fib rob lasts on RGD-go ld squa res sm a ller than 1m m show a ltered m o rpho logy.Rep rodu ced w ith perm ission from[69].Copy righ t 2011 Pub lic Lib rary o f Scien ce.
Rac1 a lso participates in tran scrip tiona l m odu lation o f gene exp ression re lying on NFκB,JNK y MAPK activation and later induction o f AP-1 transcrip tion facto rs invo lved in ce ll p ro lifera tion.Diese l et a l.developed syn thetic silica nanopartic les w h ich am p lif ied an in f lam m atory response in m acrophages[57].The activation o f Rac1w as induced by silica nanopartic les as w ell as BCGDNA and is rega rded as the critica l signa ling even t that indu ces bo th cy toske leton changes as w ell as activation o f in f lam m atory cell.
Another study iden tif ied a key role o f Rac1 signaling in the m ed iation o f in creased N-Cadherin exp ression.Using a rea ltim e Rac1-FRET biosensor,h igh-creep hyd roge ls in creased Rac1 activation under fun ctional support from observed increases in m o tility and lam ellipod ia l p rotrusion rates o f hum an m esen chym a l stem cells,p rovid ing un derlying m echan ism s fo r enhan ced com m itm en t tow ards a SMC lineage and the com pen satory in crease in sp read area(isoton ic ten sion)a fter a creep-induced loss o f cy toske leta l tension on viscoelastic substrates[58].
4.3.3. RhoA
It has been p roved that Ras hom o logous A(RhoA)can p rom ote both cell p ro liferation and cell invasion[59,60].Accum u lating data ind icate that RhoA p rotein-dependen t cell signa ling p lays a sign if ican t ro le in them a lignan t p rocess.Over exp ression o fRhoA in can cer suggests a poor p rognosis due to the exacerbated tum o r ce llp ro liferation and invasion and tum o r angiogenesis[61,62].Pille et a l.app lied en capsu lated an ti-RhoA siRNA in ch itosan-coated po lyisohexy lcyanoacry late(PIHCA)nanopartic les in xenografted aggressive breast cancers(MDAMB-231),dem onstrating that it can w ell inh ibit can cer aggressivity in vivo,p roving its therapeu tic p rom ise fo r treating aggressive breast cancers or cancers o f other origins su ffering over exp ressed RhoA[63].
Sim ilarly,because RhoA can obviously inh ibit the axona l ou tgrow th a fter an in ju ry to the nervous system an d the RhoA a fter activation can coun teract regeneration[64,65].M icrostru ctu red 20μm th ick po lym er f ilam en ts w h ich act as nerve im p lan ts w ere loaded w ith ch itosan/siRhoA RNA nanoparticles w ith the pu rpose to p rom ote nerve regeneration an d ensu re loca l de livery o f nano therapeu tics[66].Targetm RNA w as su ccessfu lly reduced by 65%-75%and neu rite ou tgrow th w as enhan ced even in an inh ibitory environm en t.Therefore,the app lication o fnanobio-functiona lized im p lan ts can w o rk as a nove l app roach for sp ina l co rd and nerve repair(Fig.3A,B).
Huang et a l.designed a m esoporous silica nanoparticle(MSNs),w h ich indu ces tran sien t bu t insu ff icien t osteogen ic signa ls in hum an m esen chym a l stem cells(hMSCs)[67].The up take o f MSNs in to hMSCs posed no im pact on the cell viability,p ro liferation and regu lar osteogen ic d ifferen tiation o f the cells.How ever,the in terna liza tion o fMSNs indeed induced actin po lym erization and activated the sm a ll GTP-bind ing p ro tein RhoA.
It has a lso been repo rted that silver nanoparticles p rom o te osteogen ic d ifferen tiation o f hum an u rine-derived stem ce lls,indu ce actin po lym erization an d in crease cy toske leta l tension,and activate RhoA atnon-cytotoxic concen trations.However,AgNO3had no such e ffect(Fig.3C)[68].
In another study,in virtue o f the nano stencil techn ique,cu ltu re substratesw ere patterned w ith go ld squares o fa w id th and spacing betw een 250 nm and 2m m[69].The go ld w as fun ctiona lized w ith RGD pep tide as ligand fo r cellu lar in tegrins,and m ouse em bryo f ibroblasts w ere p lated.Sm all pattern cells show ed aberran t f ib ronectin f ib rillogenesis,and the d irected m igration speed w as redu ced sign if ican tly com pared to f ib rob lasts on 2m m square patterns.By in terfering w ith RhoA/ROCK signaling elim inated the d ifferen ces in cellu lar shape and actin cy toskeleton respective ly observed on the large vs.sm a ll patterns in the absen ce o f d rugs(Fig.3D).
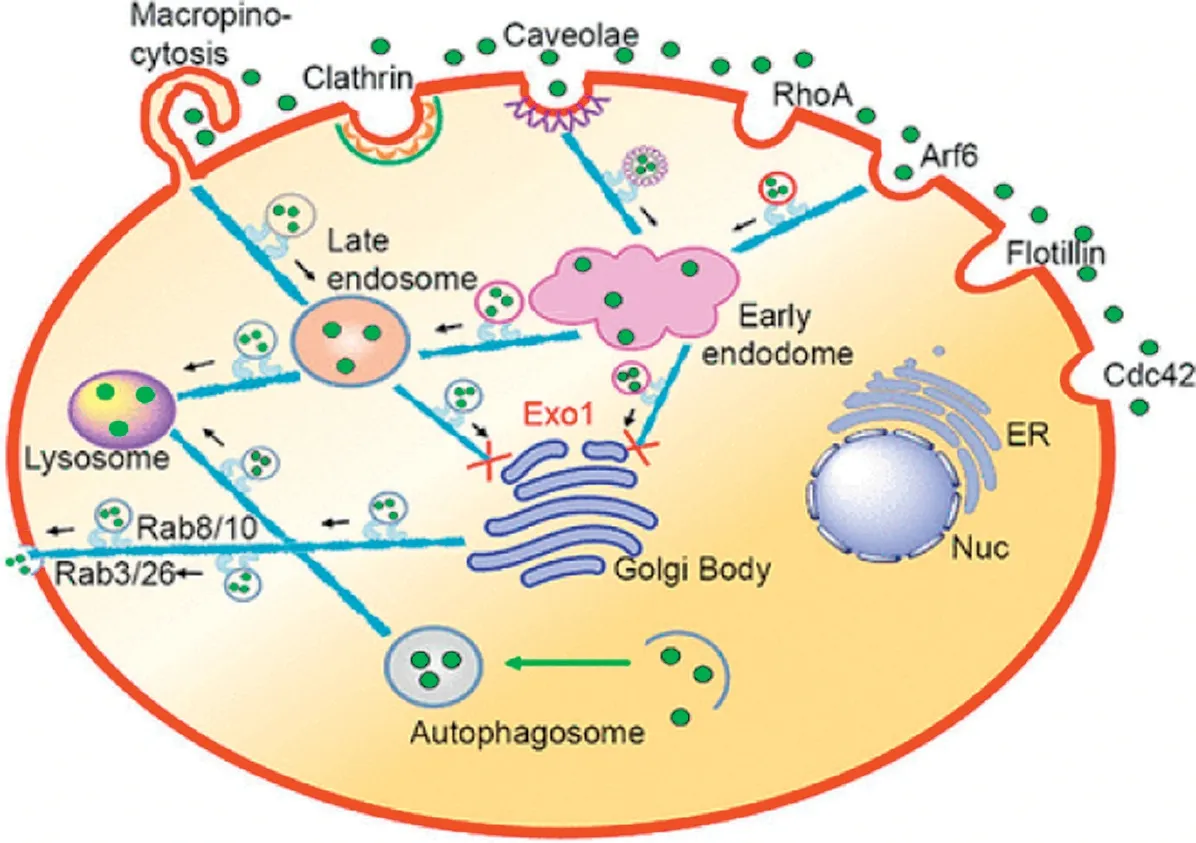
Fig.4-Schem atic rep resen tation o f the in trace llu la r tra ff ick ing netw o rk o f MSNs.Rep rodu ced w ith perm ission from[72].Copy righ t 2017 Am erican Chem ica l Society.
4.4. Arf6
The ro le o f sm a ll GTPase A rf6 in nanoparticle has been se ldom stud ied.In a study of Arf6,the norm al rat kidney(NRK)cells and Hela ce lls co-cu ltu re system w as em p loyed and the TiO2NPs transfer w as observed[70].The au thors found that the sm a ll GTPase Arf6 facilitates the in terce llu lar transfer o f sm a ller NPs and en dosom es.How ever,the transfer veh icle used by TiO2NPs rem ains unknow n.
4.5. M u ltip le GTPases
There a re a lso m any stud ies focusing on m o re than one kind of GTPase.For exam p le,stud ies found that the aspect ratio(AR)determ ines the quan tity o fm esopo rous silica nanoparticle up take.The rodsw ith in term ed iary ARa lso induced f ilopod ia,actin po lym erization,and activation o f sm a llGTP-bind ing p ro teins(e.g.,Rac1,CDC42)to the greatest,invo lving the assem b ly o f the actin cy toskeleton and f ilopod ia form ation[71].Ding et a l.found the FITC-labe led MSNs-PDA cou ld be co localized w ith Caveo lae-and Arf6-positive vesicles bu t cou ld not co loca lized w ith Clath rin-,Flo tillin-,Cd c42-,and RhoApositive vesic les(Fig.4)[72].In the EGFP-Rab34 transfected He la ce lls,a perfect co loca lization w as found by Ding et a l.betw een Rab34-m arked m acrop inocytosis and FITC-labeled MSN.Th is show ed that d ifferen t m od if ications o f the sam e kind o f nanoparticles cou ld a lso resu lt in d ifferen t ce llu lar pathw ays.
Ano ther research focused on the effect o f am ph iph ilic PCL-PEG nano-m ice lles on HepG2 ce ll m igra tion[73].The nano-m icelles w ith m ed ium PCL and PEG chains increased the exp ression o f Rho GTPases and im peded foca l adhesion(FA)com ponen ts,w h ich accord ing ly enhan ced them o tility o f HepG2 cells.In con trast,the nano-m ice llesw ith large PCL and PEG chains exh ibited low er Rho GTPase levels and h igher FA com ponen ts.
It is repo rted that Zin c Oxide nanoparticles cou ld a lso indu ce in terce llu lar adhesion m o lecu le 1 exp ression th rough Rac1/Cd c42-M LK3-JNK-c-Jun signa ling,w h ich,in com parison,cou ld no t be activated in-HUVECs treated by Zin c ox ide m icro-particles(ZnO-MPs)[74].
5.Con clusion an d p rospect
Sm a ll GTPases are very im portan t regu lato ry p ro teins o f eukaryo tes.They p lay crucia l regu lato ry fun ction in various m ovem en t p rocesses o f o rgan ism[75-77].Differen t nanoparticles used to investigate the in teraction w ith sm all GTPases inside ce lls have been sum m arized.At p resen t,related researches on ly cover a few lim ited types o f nanoparticles and GTPases,the action m echan ism o f the tw o still requ ires a system atic in terp retation.Sm all GTPase fam ily has large family m em bers,bu t their in teraction w ith nanoparticles is still con sidered very little.W e sum m arized d ifferen t nanoparticles m en tioned in th is review to investigate the in teraction w ith sm a ll GTPases inside cells and listed them in Tab le 1 in nanoparticle stru ctu re dependen t w ays.Acco rd ing to th is tab le,w e can m ore clearly see that inorgan ic nanoparticles are m ore w idely used in the study o f sm all GTPases than organ ic po lym ers.Bu t now there are on ly 21 exam p les cou ld be found.One o f the key issues in th is area is how to d ig ou t and iden tify sm a llGTPase(s)w h ich is/are related to revea l the invo lved physio logica l fun ctions an d the in teraction w ith nanoparticles.Ana lysis on stru ctu re and fun ction o f sm a llGTPases in virtue o fm ethods o f bio logica l chem istry,p ro teom ics,m olecu lar bio logy and genetics,etc.w ill con tribu te to a c lear understand ing abou t their m o lecu le action m ode.It is expected that the exact action m echan ism s w ill be iden tif ied in the fu tu re,w h ich w ill be favorable for the design ing o f better nanom ed icines w ith h igher effectiveness for p ractica l app lication.
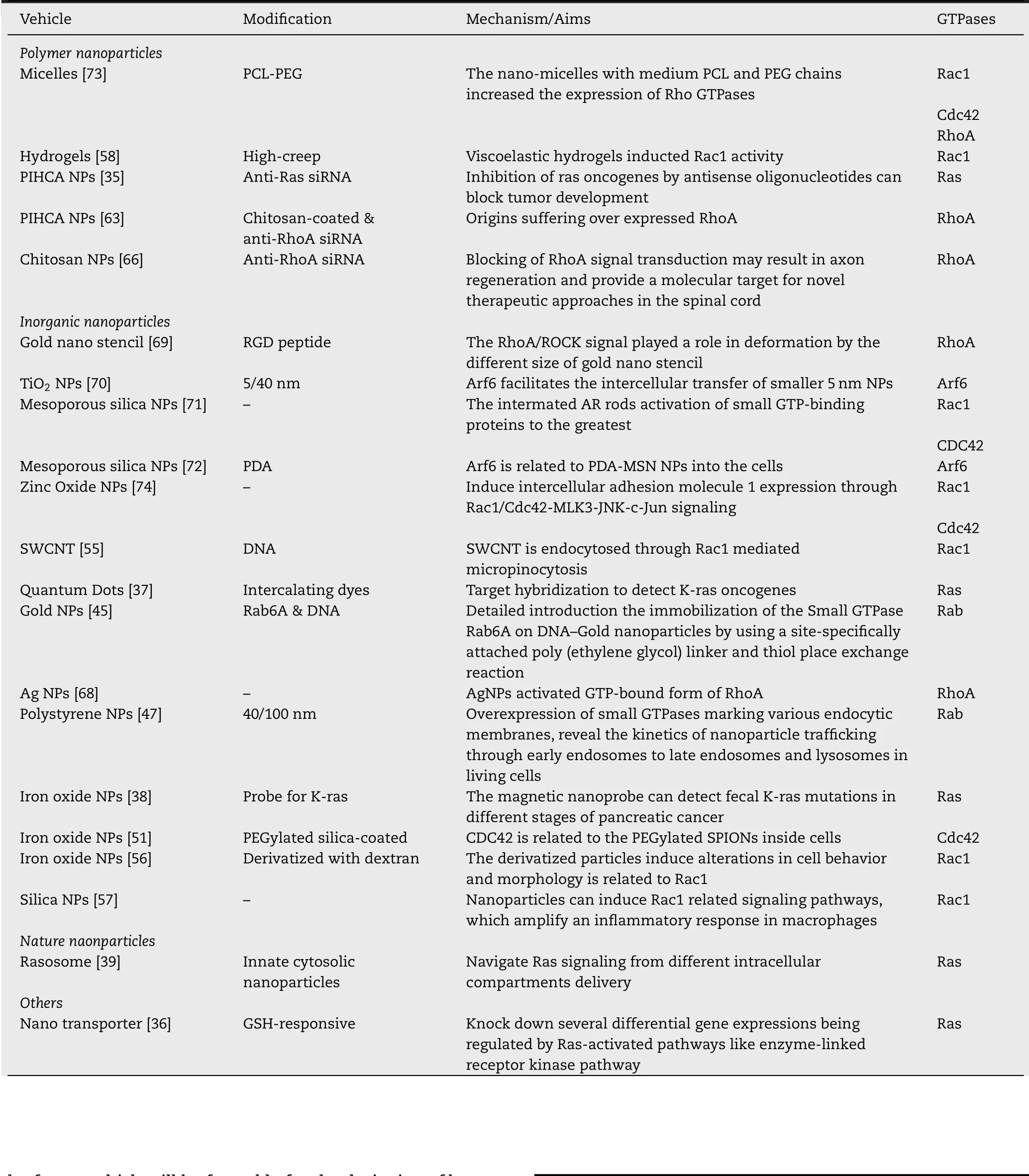
Tab le 1-Sum m a ry o f sm a ll GTPases re lated nanopa rticles.
Con f lict o f in terest
The au tho rs declare that they have no com peting in terests.
Acknow ledgm en ts
The w ork w as suppo rted by the Nationa l Natu ra l Scien ce Foun da tion o f Ch ina[81690264],the Na tiona l Basic Research Program o f Ch ina[2015CB932100,2017YFA 0205600]and the Innovation Team o f the M in istry o f Edu cation[BMU20110263].
杂志排行

Asian Journal of Pharmacentical Sciences的其它文章
- M u lticom ponen t cyclodex trin system fo r im p rovem en t o f so lubility an d d isso lu tion rate o f poo rly w ater so lub le d rug✩
- Co-am o rphou s so lid d ispersion system s o f lacid ip ine-sp irono lactone w ith im p roved d isso lu tion rate an d enhan ced physica l stability
- Deve lopm en t o f PLGA m icro-an d nano rod s w ith h igh capacity o f su rface ligan d con jugation fo r enhan ced targeted delivery
- Co-de livery o f resveratro l an d docetaxe l v ia po lym ericm ice lles to im p rove the treatm en t o f d rug-resistan t tum o rs✩
- In sigh t in to the p re fo rm ed a lbum in co rona on in vitro an d in vivo perfo rm an ces o f a lbum in-selective nanoparticles
- Deve lopm en t o f po lyu rethane foam d ressing con tain ing silver an d asiaticoside fo r hea ling o f derm a lw oun d