不同载畜率下短花针茅分蘖数量对内源激素浓度的响应
2019-05-13贾丽欣乔荠瑢赵萌莉
贾丽欣,杨 阳,张 峰,乔荠瑢,赵萌莉,*
1 内蒙古农业大学草原与资源环境学院, 呼和浩特 010010 2 内蒙古农业大学草地资源教育部重点实验室, 呼和浩特 010010
植物在不同的生长发育阶段具有不同的生理功能和环境适应能力,而内源植物激素是调节植物生长发育的开关[1]。内源激素是在植物体根、茎、叶等部分器官中合成、转移到体内其他器官后能影响生长和分化的微量有机物质,植物个体发育中,不论是种子发芽,还是营养生长和繁殖器官形成,以及整个成熟过程,都由激素控制[2]。短花针茅作为葛根塔拉草原主要的建群种和优势种,其营养繁殖的重要方式就是分蘖[3]。分蘖与株高不仅影响短花针茅的株型,且直接影响短花针茅的产量[4]。而生长素(IAA)、细胞分裂素(CTK)等内源激素是影响株高、分蘖等性状的关键因子。生长素(IAA)是植物体内分布最广泛的激素,它与质膜上一个受体结合后,活化质膜上的H+泵ATP酶,引起H+往质外体释放,H+排出膜外后,细胞质膜内产生负电势,于是K+、IAA和糖都进入细胞,引起代谢作用活跃;在离子进入细胞、水势降低时,水也进入细胞,促进细胞扩大,使细胞壁的可塑性增大,细胞伸长生长的速度加快[5]。细胞分裂素(CTK)使核糖核酸酶、脱氧核糖核酸酶、蛋白酶及肽酶等活性减弱,从而抑制核酸与蛋白质降解,促进细胞分裂与扩张,延衰与保绿;在组织培养中与生长素(IAA)协同调控细胞与器官分化,推动形态建成;还可促进侧芽生长,打破顶端优势[6]。生长素(IAA)和细胞分裂素(CTK)在植物分蘖芽的发生及伸长等发育过程中起着关键作用。有研究表明,生长素(IAA)合成于植物茎尖与幼嫩的叶中,自上而下主动运输,并直接输送到分蘖芽中,抑制分蘖芽的生长发育[7]。细胞分裂素(CTK)合成于根中,在植物失去顶端优势后,受外源生长素抑制[8]。细胞分裂素通过蒸腾作用在木质部中向上运输,对腋芽施加外源的细胞分裂素,能够直接促进腋芽的生长[9]。而放牧对植物分蘖生长的干扰影响多表现为,随载畜率的增加,生殖枝的分化率降低[3]。
目前对于牧草分蘖过程中植物激素动态变化及其与载畜率关系的研究较少。本研究利用葛根塔拉草原放牧试验地建群种短花针茅作为试验对象,比较分析不同载畜率下植物激素的变化差异,并初步分析两种主要内源激素对分蘖性状的影响。探析植物激素在放牧干扰下的生理功能,以期更深入地了解植物激素对牧草分蘖调控的作用机理。
1 材料与方法
1.1 试验地概况
试验区位于阴山北麓,内蒙古自治区乌兰察布市西北部的葛根塔拉草原,四子王旗王府一队,内蒙古农牧科学院草原研究所荒漠草原实验基地(41°47′17″N,111°53′46″E)。地势平坦开阔,海拔1450 m。该旗地处中温带大陆性季风气候区,具有干旱、少雨、多风和蒸发量大的气候特征。年均气温3.4℃,月平均温度最高月为6—8月,年均气温分别为21.5、24.0℃和23.5℃,≥0℃积温为2500—3100℃,≥10℃的年积温为2200—2500℃。无霜期90—115天;平均日照时数为3117.7 h。年均降水量299.4 mm,且年际变化较大,降水主要集中在5—9月,这段时间的降水量占全年总降水总量的80%以上。年潜在蒸发量约2300 mm,远超过降水量,是降水量的7—10倍,高蒸发量致使空气干燥,干旱严重。草地植被类型为短花针茅(Stipabreviflora)+冷蒿(Artemisiafrigida)+无芒隐子草(Cleistogenessongorica)。草层低矮,平均高度为8 cm,盖度为15%—25%,植被较稀疏。植物群落由20多种植物组成,主要为多年生草本短花针茅、无芒隐子草、银灰旋花(Convolvulusammannii)、阿尔泰狗娃花(Heteropappusaltaicus)、冰草(Agropyroncristatum)、羊草(Leymuschinensis)、细叶葱(Alliumtenuissimum)、蒙古葱(Alliummongolicum)等;一、二年生草本有猪毛菜(Salsolacollina)、猪毛蒿(Artemisiascoparia)、栉叶蒿(Artemisiapectinata)等。还有灌木小叶锦鸡儿(Caraganamicrophylla)、矮灌木狭叶锦鸡儿(C.stenophylla)、半灌木驼绒藜(Ceratoideslatens)、小半灌木木地肤(Kochiaprostrata)、冷蒿等。土壤为淡栗钙土,土层厚度大约为1 m,在40—50 cm深处有钙积层,土体坚硬且渗透能力差,有机质含量较低。土壤微生物有好气性细菌、放线菌和霉菌等。
1.2 试验设计和测定方法
1.2.1样地概况
葛根塔拉草原放牧试验始于2003年6月,至今已有十四余年,完全随机区组试验设计的采用,使得围封的草地被划分为3个区组,每个区组设置4个处理,分别为对照区(CK)、轻度放牧区(LG)、中度放牧区(MG)、和重度放牧区(HG),每个处理设置3个重复,共计12个小区(图1)。载畜率值的设定以多年研究调查以及实地勘测为依据,依次设置为0、0.91、1.82和2.71 羊单位/公顷/每半年。每个区组内的4个处理完全随机排列,每个小区的面积约为4.4 hm-2,依次放羊0只(对照)、4只(轻度)、8只(中度)和12只(重度)。放牧种为当地成年蒙古羯羊,体重约67.8公斤。每年6月1日开始至11月30日结束,进行为期半年的放牧。每日6:00点将绵羊赶入各小区让其自由采食,18:00点赶回圈内饮水、休息,不进行补饲。

图1 实验小区示意图Fig.1 Diagrammatic illustration of the experimental blockCK,对照control;LG,轻度放牧light grazing;MG,中度放牧moderate grazing;HG,重度放牧heavy grazing
1.2.2试验设计
于2017年8月3日在格根塔拉草原放牧样地取样,每个处理随机采取9株植物样品。分单株采集,大、中、小株丛径的短花针茅各3株并测量其分蘖数量,其中,大株短花针茅的株丛径大约为15 cm左右、中株短花针茅的株丛径大约为10 cm左右、小株丛短花针茅的株丛径大约为5 cm左右。随后齐地面刈割装入样品盒,以冰盒冷冻保存,备测激素含量。
1.2.3激素含量测定
按照杨建昌等[10]的方法提取和纯化样品中IAA和CTK,其中CTK组分包含玉米素+玉米素核苷(Z+ZR)和异戊烯基腺嘌呤+异戊烯基腺嘌呤核苷(ip+ipR),采用酶联免疫法测定其含量[11]。试剂盒购自中国农业大学,使用酶标仪为Spectramax plus型。
1.2.4数据分析方法
采用SAS.V8在a=0.05显著性水平下进行单因素方差分析(One-way ANOVA),SigmaPlot.12.5相关分析绘制图表。
2 结果与分析
2.1 载畜率对短花针茅内源激素浓度的影响
对小株、中株丛和大株丛短花针茅生长素(IAA)、细胞分裂素(CTK)浓度与载畜率进行拟合发现:小株丛、中株丛短花针茅的生长激素浓度与载畜率之间呈正显著相关关系(P=0.0054r2=0.55;P=0.02r2=0.43),表明大、小株丛短花针茅生长素含量随着载畜率的增加而增加;
而大株丛短花针茅生长激素浓度与载畜率之间未表现出相关关系(P=0.135)(图2)。小株丛、中株丛短花针茅细胞分裂素(CTK)浓度与载畜率之间呈正显著相关关系(P=0.0018r2=0.64;P=0.0193r2=0.43),随着载畜率的增加而增加,然而,大株丛短花针茅细胞分裂素(CTK)浓度与载畜率之间未表现出相关关系(P=0.1176)(图2)。
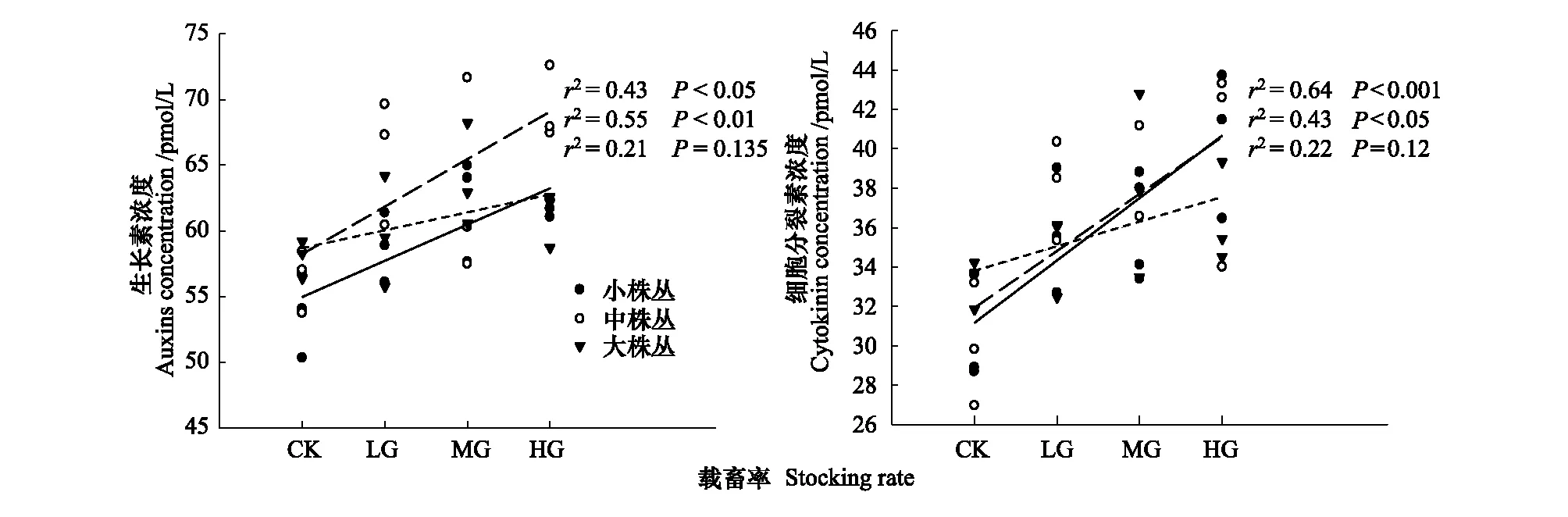
图2 载畜率对激素浓度的影响Fig.2 Effect of stocking rate on hormone concentrationP,显著性水平;r2,决定系数
对不同载畜率下短花针茅生长素(IAA)浓度进行差异性分析得知,小株丛短花针茅生长素(IAA)浓度在中度放牧区达到最大值为62.21 pmol/L,对照区小株丛短花针茅生长素(IAA)浓度显著低于中度和重度放牧区(P<0.05;图3)、中株丛短花针茅生长素(IAA)浓度在重度放牧区达到最大值为69.32 pmol/L,对照区中株丛短花针茅生长素(IAA)浓度显著低于重度放牧区(P<0.05;图3)、大株丛短花针茅生长素(IAA)浓度在不同载畜率处理下并未表现出显著差异性。对不同载畜率下短花针茅细胞分裂素(CTK)浓度进行差异分析得知,小株丛短花针茅细胞分裂素(CTK)浓度在重度放牧区达到最大值为40.55 pmol/L,对照区小株丛短花针茅细胞分裂素(CTK)浓度显著低于中度和重度放牧区(P<0.05;图3)、中株丛短花针茅细胞分裂素(CTK)浓度在重度放牧区达到最大值为39.97 pmol/L,对照区中株丛短花针茅细胞分裂素(CTK)浓度显著低于轻度和重度放牧区(P<0.05;图3)、大株丛短花针茅细胞分裂素(CTK)浓度在不同载畜率处理下并未表现出显著差异性。

图3 不同载畜率下激素的浓度Fig.3 Hormone concentration in different stocking rate不同小写字母表示不同载畜率下激素浓度差异显著(P<0.05)
2.2 载畜率对短花针茅分蘖数量的影响
放牧能够在一定程度上影响短花针茅分蘖的数量(表1,P<0.05)。重度放牧是增加短花针茅分蘖数量最显著的载畜率,这一现象在中、小株丛上体现的尤为明显(图4,P<0.01)。小株丛短花针茅是最易受放牧影响的株丛类型,中度、重度放牧均能够使其分蘖数量增加(图4,P<0.01)。而大株丛是稳定性最好且最不易受放牧影响的株丛类型,与对照区相比,其余载畜率均未使大株丛短花针茅分蘖的数量受到干扰(图4,P=0.98)。短花针茅基丛径的大小是控制分蘖数量的主要原因(表1,P<0.001),放牧也是影响分蘖数量的原因之一(表1,P<0.05),然而基丛径与载畜率之间并无交互作用(表1)。

表1 载畜率对短花针茅分蘖数量影响的方差分析
T,载畜率,Stocking rate;R,基丛径,Diameter of stem;*,P<0.05;***,P<0.001

图4 载畜率对短花针茅分蘖数量的影响Fig.4 Effect of stocking rate on tiller 不同小写字母表示不同载畜率下激素浓度差异显著(P<0.05)
2.3 内源激素浓度对分蘖数量的影响
短花针茅的分蘖数量与生长素(IAA)浓度之间存在一种极显著相关的二次关系(P=0.0065,r2=0.263),满足方程y=-0.0001x2+0.07x+49.92(图5)。当短花针茅体内生长激素的浓度超过63 pmol/L后,短花针茅的分蘖数量将不会超过420个/株,说明过高浓度的生长素(IAA)会抑制短花针茅的分蘖数量(图5)。而细胞分裂素(CTK)与短花针茅的分蘖数量之间不存在显著相关关系(P=0.1126,r2=0.124)(图5)。

图5 分蘖与激素浓度的关系Fig.5 Relationship between hormone concentration and tiller
3 讨论
3.1 放牧对短花针茅分蘖数量的影响
短花针茅通过有性繁殖产生实生苗的数量及其微少,小的实生苗在群落中难以成功存活所以其对种群数量的影响可以忽略不计[12]。因此通过无性繁殖产生分蘖,增加新的枝条才是增加种群个体数量的有效途径,故探索分蘖对载畜率的响应[13]。有研究表明:过度放牧干扰了小嵩草(Kobresiapygmaca)型草地及其原生植被优势种的生长生殖特征,使得原优势种的株丛数,分蘖,分枝数均有不同程度的下降[14]。2016年古琛对短花针茅的繁殖策略进行研究时发现:轻度放牧区短花针茅的分蘖数要显著高于中度、重度和不放牧区域[4]。本试验得到的结论与上述均不相同:即放牧能够在一定程度上影响短花针茅植株个体的分蘖数量(P<0.05),重度放牧是增加短花针茅植株个体分蘖数量最显著的载畜率。原因有一下几点:(1)本试验为规避丛径对分蘖造成的影响,特设定大、中、小三种丛径范围进行取样,比较不同载畜率下,同等丛径大小植株的分蘖数量。与上述实验的测量方法不同。(2)上述文献均为样方取样而本试验为单株取样,样方取样比较的是群落差异,而单株取样比较的是个体上的差异。
重度放牧能够增加短花针茅个体的分蘖数量,却不能增加种群的分蘖,最为显著的一个原因是,重度放牧使短花针茅株体趋于小型化,小型短花针茅在重度放牧样地占的比重相对较高[15]。并伴随群落高度降低,使其形成了一种避食性的生态适应对策[16]。但是草地植被表型特征的改变不是放牧单一作用的结果,它与牧草的遗传特性、生理特性、气候变化和土壤养分等因素都有密不可分的关联[17]。有研究植被通过改变叶片角质层的厚度或表皮细胞的面积大小影响植物叶片解剖结构来适应放牧胁迫,以恢复植株的光合作用能力、增加光合速率和繁殖的适应性[18]。通过比较过牧和正常放牧条件下种群的形态特征和遗传多样性,发现过牧分割植物群体,降低草丛高度,增加营养枝的密度,减少繁殖枝的密度。但在两个种群间未发现明显的遗传学差异。形态特征的差异可能是由于放牧条件下微环境的变化造成的[19]。2006年刘忠宽研究不同放牧强度草原休牧后土壤养分和植物群落变化特征时发现:植物群落的生物量、群落高度、群落密度与土壤养分分布均表现一致的正相关关系,但只有植物群落生物量和群落高度与土壤有机碳、全氮、无机氮、土壤有效磷和有效硫相关显著(P<0.05)[20]。除了受环境因子、遗传特性的影响还存在许多未被深入研究的因素,所以影响短花针茅分蘖的内在机制还有待进一步探索。
3.2 载畜率对内源激素的影响及其与分蘖的相关关系
家畜对草地采食践踏等行为对植物构成了一定程度的损伤,这使放牧过程中成为一种逆境条件。为了减轻植物损伤,细胞内的内源激素会做出相应的调节从而影响植物的生理代谢[21]。放牧强度过大导致克隆整合的能量投入远远大于该特征的正向产出[22]。从表观来看,植物能够在草食动物采食之后能够进行补偿性再生生长,弥补动物采食所造成的损失。事实上,植物的补偿性效应是与碳水化合物和氮素在植物体各部分之间的转移,以及外界的环境干扰对植物体内源激素的产生及传递直接相关的[23]。王静研究放牧退化群落中冷蒿种群生物量资源分配的变化发现:载畜率的增加,可以使冷蒿体内生长素(IAA)的含量下降,从而消弱顶端优势、抑制植物细胞的伸长,增强分蘖[24]。这与高莹年模拟羊草种群放牧耐受性研究时得到的结论相反,即分蘖芽生长促进型激素(IAA)在载畜率增加时增加[23]。本试验的研究结果与高莹的研究结果一致,即放牧能够显著增加内源激素的浓度(P<0.05),但这种相关只存在于中、小株丛的短花针茅中。这是由于植物激素是植物响应环境胁迫的信号转导的主要成员,当植物受到放牧胁迫后,植物的生长和发育受到抑制,激素含量会发生变化,以缓解环境胁迫对植物的伤害[25]。如生长素(IAA)含量的增加抑制了花芽的分化作用以减少分蘖芽的生长。本试验研究结果表明细胞分裂素(CTK)与短花针茅的分蘖数量之间尚未发现相关关系。但细胞分裂素(CTK)与水稻分蘖的相关研究多为:细胞分裂素(CTK)直接产生作用促进水稻分蘖芽的发生[2]。此外刘杨在研究细胞分裂素对水稻分蘖相关基因表达的调控时发现:外界因素对分蘖芽生长的调控至少存在2条途径,其中关键的一条就是通过调控节和芽中细胞分裂素(CTK)含量,进而调控芽中OsTB1等基因表达[26]。
4 结论
放牧能够在一定程度上影响短花针茅植株个体的分蘖数量(P<0.05),载畜率影响短花针茅植株个体分蘖数量,特别是重度放牧下。
内源激素与载畜率之间存在显著相关关系,放牧能够显著增加内源激素的浓度(P<0.05),但这种相关只存在于中、小株丛的短花针茅中。
过高浓度的生长素(IAA)会抑制短花针茅的分蘖数量(P<0.01)。而细胞分裂素(CTK)与短花针茅的分蘖数量之间尚未发现相关关系。
