焦虑、风险决策及其关系的神经机制
2019-04-29
(1.西南大学心理学部,重庆 400715;2.美国密苏里大学圣路易斯分校教育学院,圣路易斯 63121;3.重庆城市职业学院,重庆 402160)
1 引 言
近年来,越来越多的研究者高度地关注焦虑对风险决策的影响,并进行了一系列有价值的研究,但所呈现的结果尚未一致。
如同研究所示,与低特质焦虑者相比,高特质焦虑者更加倾向于风险寻求(Robinson,Bond,& Roiser,2015)。但有研究却呈现了不一致的结果,即高焦虑的决策者更加偏好于风险规避(Charpentier,Aylward,Roiser,& Robinson,2017)。正如研究者所述:高焦虑者对环境中的潜在威胁或负性刺激较为敏感,对此会过度地警惕并产生注意偏向(Ortega,Ramírez,Colmenero,& García-Viedma,2017),且极大可能地将其解释为负性结果(Gu et al.,2017)。故而,风险规避或许是为了避免受到伤害或是负性情绪体验的结果。然而有研究却表明,短暂的焦虑情绪在未能影响决策者的风险偏好时却改变了其大脑评价机制的编码(Engelmann,Meyer,Fehr,& Ruff,2015)。
鉴于此,或许明晰焦虑影响风险决策的神经机制,是能够直观地阐述两者关系的途径之一。本文回顾了近几年有关焦虑与风险决策的神经机制的研究,试图探索了焦虑影响风险决策的神经机制,并借用风险敏感理论与认知资源理论,从情绪与认知双加工角度诠释了此神经机制。
2 焦虑与风险决策的神经机制
2.1 焦虑的神经机制
2.1.1 焦虑与潜在威胁
焦虑个体对所处环境中存在的潜在威胁比较敏感,进而会夸大其可能对自身造成的伤害(Ortega et al.,2017)。前人在探究大脑皮层对面孔威胁性变化时发现,腹内侧前额叶(VMPFC)能够识别面孔微小变化,而杏仁核(amygdala)能够识别中性面孔到恐惧面孔的变化(Achaibou,Loth,& Bishop,2016)。并且当个体置身于威胁中时,杏仁核将直接编码从感觉皮层检测到的威胁信息,随后终纹床核(BNST)、纹状体(STR)、腹侧海马(VHPC)以及内侧前额叶等会依据编码信号进行表达,同时内侧前额叶与腹侧海马也会将反应信号传送回杏仁核、纹状体与终纹床核,以此形成一个循环的网络结构(Lago,Davis,Grillon,& Ernst,2017)。
2.1.2 焦虑与负性预期、恐惧
高特质焦虑群体的核心特征是对将来不确定事件的过度负性预期(Grupe & Nitschke,2013)。而且在他们预期负性刺激时,前脑岛(AI)被过度地激活,且激活强度与特质焦虑水平呈正相关(Simmons et al.,2011)。然而,焦虑者对负性刺激的预期除了反映对潜在威胁的应对准备之外,同时也激活了与恐惧情绪唤醒有关的神经区域(内侧前扣带回(MACC)、纹状体、杏仁核、腹内侧前额叶、脑岛(insula)以及背外侧前额叶(DLPFC)等)。其中,杏仁核主管威胁刺激的加工,背外侧前额叶与腹外侧前额叶(VLPFC)主管注意指向加工、注意偏差与警觉维持;同时,腹外侧前额叶调解着背外侧前额叶对杏仁核的影响,而前额叶则控制杏仁核的恐惧反应程度,海马(hippocampus)则为杏仁核的恐惧反应提供场景(Buhle et al.,2014;Milad & Quirk,2012)。
2.1.3 焦虑障碍患者的脑机制
广泛性焦虑障碍(GAD)患者的左侧杏仁核与左背外侧前额叶的功能连接显著降低,但右侧杏仁核与右侧小脑、脑岛、颞上回的功能连接却显著增强,且增强强度与焦虑水平呈负相关(Liu et al.,2015)。而社交焦虑障碍(SAD)患者却呈现出眶额叶(OFC)对杏仁核情绪表达的自上而下监管机制的损坏,具体表现为二者功能连接的异常积极兴奋,且这种独特的连接方式促进了杏仁核的激活强度(Sladky et al.,2015)。此外,背侧前扣带回(dACC)和背外侧前额叶的激活强度与SAD患者的症状严重程度呈负相关(Heitmann et al.,2017)。
综上所述,高焦虑的神经网络主要由眶额叶、丘脑与扣带回组成,而低焦虑的神经网络主要包括默认网络(杏仁核、前额叶、脑岛、前脑岛、纹状体等)(Takagi et al.,2017),其中杏仁核或许充当着这些脑区间运输信息的枢纽。基于此,本文推测焦虑的产生可能存在一个高度敏感的“神经回路”(如图1):杏仁核直接编码从感觉皮层检测到的信息,以自身为枢纽将编码信号分别传送至威胁加工与预期奖赏系统进行深度加工与解释,其中脑岛与扣带回负责辨别与处理刺激信息,背外侧前额叶与腹外侧前额叶负责精细加工与结果解释,而前额叶与眶额叶负责监管杏仁核对加工结果的情绪表达,海马则为杏仁核的情绪反应提供场景。但在此过程中,刺激线索的上层传递(皮层)与下层传递(脑干)或许存在解体与协调缺陷,致使各个通路的协调作用将信息结果错误地传达至杏仁核,使其高度激活,进而促使核心网络间的功能连接异常,产生焦虑。
图1 焦虑的脑功能网络图
→刺激传入通路←┄→与焦虑产生相关的脑网络┄→监控与场景作用机制
2.2 风险决策的神经机制
2.2.1 决策选项的分布
决策选项的分布(选项的变异范围、期望值与参照点的关系)会影响决策者的选择(Mishra,2014)。其中,变异范围与参照点主要涉及背内侧前额叶(DMPFC)、眶额叶、腹侧纹状体与前扣带回等(Blankenstein,Peper,Crone,& van Duijvenvoorde,2017)。眶额叶作为“计算中枢”处理着刺激与结果的价值信息;内侧前额叶作为“执行中枢”,将刺激与结果的联结信息汇总形成执行策略并保存,然后参照新的信息进行策略更新(黄文强,杨沙沙,&于萍,2016);前扣带回既预测决策结果的价值信息又将整合的奖赏信息发送至运动区(Hyman,Whitman,Emberly,Woodward,& Seamans,2013)。而期望值则调节以内外侧前额叶为中枢的价值敏感性网络的有效连接,使其密集的连接并迅速的形成最短中心路径,以便快速计算(Minati,Grisoli,Seth,& Critchley,2012)。
2.2.2 风险容忍与风险态度
研究表明,腹内侧前额叶的损伤会改变对预期收益与损失的相对风险容忍度,即腹内侧前额叶病变的患者在损失情景中倾向于风险寻求,但在收益情景中倾向于风险规避(Pujara,Wolf,Baskaya,& Koenigs,2015)。并且个体在预期不确定性收益时,激活了伏核(NAc)与前脑岛,但在预期不确定性损失时,只激活了前脑岛(Knutson et al.,2014)。这暗示着,腹内侧前额叶与前脑岛或许是风险容忍度的神经基础。
此外,在风险寻求的个体中,腹外侧前额叶的信号值随着风险而增大,但在风险规避的个体中却呈现了相反现象,这由此反映了风险态度(B?mmel et al.,2014)。并且个体较多的风险寻求也与内外侧眶额叶中较强的激活有关(Blankenstein et al.,2017)。故而,风险态度的神经表征主要涉及到腹内外侧前额叶、左侧眶额叶(LOFC)、内侧眶额叶(MOFC)与后扣带回(PCC)(Losecaat Vermeer,Boksem,& Sanfey,2014)。
综上所述,风险决策涉及的特定脑区有前额叶、眶额叶、前扣带回、纹状体、脑岛与杏仁核等(Blankenstein et al.,2017;Claus et al.,2017)。其中,眶额叶与伏核编码了风险决策中的概率与风险等信息;而眶额叶与内侧前额叶参与了风险决策的策略形成与转换,且杏仁核、腹侧纹状体与伏核等参与了策略保持、价值判断,影响了决策偏向与行动强度(黄文强et al.,2016)。而且风险决策由刺激编码、比较与行为选择组成。故本文推测风险决策的形成可能存在这样的一个神经环路(图2):首先刺激编码系统通过对外界刺激信息的加工而形成指令评价编码,随后预期奖赏系统对此编码进行深度解释而产生不确定性预期奖赏信号,然后行为选择系统参照此奖赏信号而形成风险态度指令行为编码,产生相应的决策行为。

图2 风险决策的脑功能网络图
→刺激输入路径←┄→与风险决策相关的脑网络
3 焦虑影响风险决策的神经机制
3.1 影响因素涉及的脑区
3.1.1 框架效应
罗寒冰,徐富明,李彬,张慧,和李燕(2015)综述道:框架效应的产生可能源自以杏仁核为代表的情绪加工,而以前扣带回与前额叶为代表的认知加工可能对此起到抑制与监控作用。当决策与框架效应相一致时,特质焦虑与杏仁核和腹内侧前额叶的功能连接呈正相关;但当决策与框架效应背道而驰时,特质焦虑与腹内侧前额叶和背侧前扣带回的功能连接呈负相关(Xu et al.,2013)。这暗示着,框架效应与焦虑反映间存在着显著重叠的神经回路,其中焦虑的产生主要依赖于杏仁核与腹内侧前额叶间的情绪神经回路,而决策评价主要依赖于前扣带回与前额叶间的认知神经回路,则这三个脑区间构成的神经回路可能是焦虑通过加工框架信息而影响风险决策的神经基础。
3.1.2 主观评价
研究证实,在没有显著的行为偏好的风险决策中,短暂的焦虑干扰了腹内侧前额叶与腹侧纹状体的正值编码,使其主观值减小,但同时又促进了脑岛的负值编码,使其主观值增大,即短暂的焦虑能够把决策选项的神经值的焦点从可能的正性结果转向为预期的负性结果(Engelmann et al.,2015)。这表明,状态焦虑在改变低水平的大脑评价系统时有可能未改变更加高级的“慎重”过程(例如:价值的整合与比较)。同时,这也暗示着大脑评价系统或许是焦虑影响风险决策的神经基础。
3.1.3 损失反感
Galván和Peris(2014)的研究发现,在损失情景中进行风险决策时,焦虑者呈现出较强的杏仁体与脑岛的衔接,而在收益情景中腹侧纹状体的激活强度显著降低。这就表明,与收益情景相比,焦虑者的杏仁核和腹侧纹状体对损失情景的反应更加强烈,即损失反感(Canessa et al.,2013)。此外,损失反感与双侧脑岛、左内侧额叶的灰质体积呈负相关(Markett,Heeren,Montag,Weber,& Reuter,2016),也与左腹侧纹状体、右后侧脑岛的激活强度高度相关(Canessa et al.,2016)。这暗示着,损失反感主要涉及的脑区有杏仁核、腹侧纹状体与脑岛。
综上所述,焦虑影响风险决策的因素主要涉及的脑区包括杏仁核、腹内侧前额叶、前扣带回、腹侧纹状体、前脑岛与脑岛等。基于此,本文探索了这些脑区在焦虑影响风险决策过程中所扮演的角色。
3.2 焦虑影响风险决策涉及的脑区及其功能
3.2.1 前额叶及其认知灵活性
焦虑的消极影响会干扰个体的认知功能,特别是认知灵活性与决策。然而,前额叶的完整性是认知功能的基础(Park & Moghaddam,2017)。在焦虑环境中,前额叶通过配对神经元的相关性以及同步的θ振荡,与杏仁核相互作用,调节着个体的条件性恐惧反应与冒险行为(Karalis et al.,2016;Likhtik,Stujenske,Topiwala,Harris,& Gordon,2014)。而在转换任务中,焦虑的变化与前额叶神经元对任务规则的反应缺陷有关,同时其也抑制了前额叶神经元的自动激活(Park,Wood,Bondi,Del,& Moghaddam,2016)。但前额叶神经元激活的持续变化可能与焦虑相关的神经化学物质的变化有关(如:多巴胺与去甲肾上腺素的释放),并在此延长时间尺度上,焦虑诱发了前额叶神经元自发的异常激活状态(Park & Moghaddam,2017)。这暗示着,焦虑可能通过损伤认知灵活性区域而影响着风险决策。
3.2.2 纹状体的注意偏向与腹侧纹状体的规避动机
在个体关注任务时,携带多巴胺神经递质的纹状体通过调节前额叶、顶叶与扣带回的激活强度而影响着注意力(van Schouwenburg,den Ouden,& Cools,2015)。而且与高奖励相比较,焦虑者更加倾向于低奖励,且低奖励较强的激活了纹状体。这或许是焦虑者对低奖励的相对负性值较为敏感,进而产生了注意偏向(Benson,Guyer,Nelson,Pine,& Ernst,2015)。此外,当行为被抑制的青少年察觉到他们的决策将直接影响到预期结果时,其的腹侧纹状体呈现出超活化状态(Guyer et al.,2012)。并且个体在预期损失时,焦虑促进了腹侧纹状体的激活(Gorka,Fuchs,Grillon,& Ernst,2017)。这暗示着,焦虑者偏好于规避负性结果的可能性要归结为有助于风险规避的高动机,而这种高异常地动机可能是腹侧纹状体的功能障碍所致(Benson et al.,2015;Lago et al.,2017)。
3.2.3 前扣带回与前脑岛及其损失反感
Fukunaga,Brown,和Bogg(2012)的研究发现,随着气球爆破概率的增大而被试选择放弃继续打气球时,前扣带回与前脑岛呈现出较强的激活状态,其中前扣带回较强的激活可能代表着损失反感(Flagan,Aupperle,Simmons,B.Stein,& Paulus,2011)。然而有研究表明,焦虑不但促进了背侧前扣带回的损失编码(Gorka et al.,2017),而且高特质焦虑与较强的脑岛激活密切相关(Flagan et al.,2011),这可能反映了脑岛对风险强烈的反感内感受性。因此,反感感应加工的神经机制(杏仁核、脑岛与前扣带回)或许能够解释焦虑者规避潜在损失的产生与功能紊乱(Flagan et al.,2011)。
3.2.4 脑区间的神经环路及其功能
Lago等人(2017)综述道:焦虑的神经环路由杏仁核、内侧前额叶、脑岛、纹状体、终纹床核和海马等间的功能连接构成。并且在风险决策期间,高焦虑者借助框架效应的神经环路(杏仁核(情绪系统)-腹内侧前额叶(整合信息)-背侧前扣带回(DACC)(分析系统))对风险信息进行评估,其中评估的脑网络主要由腹内侧前额叶、腹侧纹状体、前脑岛与前扣带回等构成(Fukunaga et al.,2012;Heekeren,Schulreich,Mohr,& Morawetz,2017;Xu et al.,2013),而对评估的结果产生损失反感的神经环路由杏仁核、前扣带回与前脑岛间的功能连接构成(Flagan et al.,2011)。
综上所述,焦虑影响风险决策主要涉及的脑区有杏仁核、前额叶、腹内侧前额叶、脑岛、前脑岛、纹状体、腹侧纹状体与前扣带回等。然而,焦虑对风险决策的影响不但涉及情绪加工,而且也涉及认知加工,特别是认知灵活性转换、注意偏向等高级加工过程。基于此,本文推测焦虑影响风险决策可能存在这样的一个神经回路(图3):以杏仁核为交汇点,以前额叶区域为桥梁形成情绪与认知双加工的主干路径,其中框架效应的神经环路或许是情绪与认知加工间的一条捷径。焦虑者通过调节环境中的情绪信息,进而协助认知加工系统对输入大脑评价系统中的信息进行全面整合与综合比较,从而指导行为决策。若主干路径受损(主要指前额叶区域的病变),杏仁核会借助脑岛加工冗余信息的特点,及时与其搭建一个简单的信息处理平台,以此协助个体应对信息的变化。
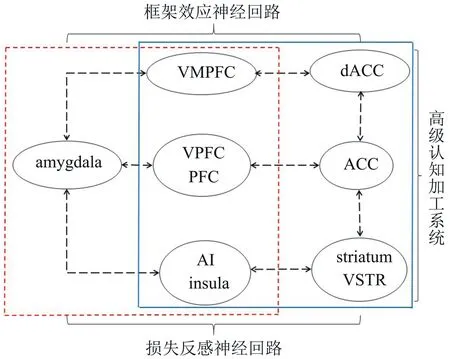
图3 焦虑影响风险决策的脑功能网络图
←┄→焦虑影响风险决策的脑网络
情绪加工系统 □认知加工系统
4 总结与展望
本文通过综述有关焦虑与风险决策的神经机制发现,二者涉及的脑区间存在着高度重叠。而这种独特的重叠不仅暗示着焦虑影响风险决策神经机制的普遍性,而且也暗示着其的复杂性。如上所述,焦虑影响风险决策的过程不但涉及到情绪与认知的双加工,而且可能涉及到焦虑的三个阶段。在短暂焦虑状态时,焦虑可能更多改变了决策者内部的神经编码而未能影响行为偏好;在焦虑的潜伏期,焦虑可能过多地干扰了决策者的神经评价,抑制了其的认知灵活性转换,进而逐渐地削弱了神经评价系统的功能,此时或许会呈现行为偏好;但在焦虑的病变期,焦虑严重地损伤了决策者的大脑机制,致使其呈现出诸多地功能障碍,从而高频率的产生异常行为。基于此,本文借用风险敏感理论与认知资源分配理论诠释了此过程。
风险敏感理论的核心观点为:“需要”驱动决策行为(宋云嫱,徐瑞珩,&邢采,2017)。在风险决策情景中,由于风险的不确定性诱发了个体的焦虑情绪,进而提高了个体对所处环境中潜在风险的敏感性,并对此产生了注意偏向且优先加工,对加工的结果更倾向于负性解释。故而为了避免受到伤害或是负性情绪体验以此满足自身安全的“需要”,短暂的焦虑可能促进了个体的风险规避。但长期的焦虑情绪会削弱认知资源的分配,进而抑制个体的认知加工,致使其因认知资源的紧缺而没有能力做出正确判断时,会迫切地选择能提供一定机会而满足自身需要的高风险选项。
然而,未来的研究不但需要验证焦虑影响风险决策的神经机制,还需要探索神经元的连接状况以及神经递质的流向。故而可以考虑从以下几个方面进行:
第一,遗传基因的研究。研究发现,5-HTTLPR与青少年的焦虑障碍和损失规避有关,而且遗传基因通过神经递质的传导影响着人类的神经活动以及相应的决策行为(Ernst et al.,2014)。第二,神经递质的研究。采用正电子断层扫描(PET)技术对风险决策背后的神经递质活动进行分子水平研究,以此探索焦虑影响风险决策的脑神经回路信息传递状况(如:传递了何种信息、传递的信息量、信息传递的方向)。第三,开展焦虑影响风险决策的发展性研究。利用近红外脑功能成像技术(fNIRS)生态学效度高的特点,可考虑对婴儿群体开展研究,以揭示人类早期的焦虑加工与风险决策及其关系的发展特点,还可开展焦虑症患者的风险决策发展特征与前额叶高级认知功能间关系的研究,以揭示焦虑影响风险决策的认知脑机制。
