濒危植物中甸刺玫种子休眠及其生态学意义
2019-04-27潘丽蛟关文灵李懿航
潘丽蛟,关文灵,李懿航
(1.昆明医科大学资产管理处,云南 昆明 650201;2.云南农业大学园林园艺学院,云南 昆明 650201;3.昆明冶金高等专科学校后勤管理处,云南 昆明 650201)
中甸刺玫Rosa praelucens为蔷薇科蔷薇属落叶灌木,是中国云南的特有种,仅分布在云南中甸高原,是典型的狭域分布种。中甸刺玫花大色艳,有香味,开花繁茂,是蔷薇属野生资源中的大花类型,具有极高的观赏性,是十分珍贵的高山花卉资源,是高海拔地区园林绿化的树种,也是月季杂交育种的重要亲本材料,其果实富含营养。因此,中甸刺玫具有极高的保护和开发价值。该物种野生种群分布点有限且分散,生境脆弱,天然更新困难,乱挖乱采现象严重,分别被《中国物种红色名录》、《中国生物多样性红色名录》列为易危物种和极危物种[1]。目前,对中甸刺玫的研究主要在种群分布特征[2]、引种驯化[3]、杂交起源[4]等方面,对其种子休眠和萌发方面的报道非常少。
濒危植物的种子休眠特性是导致植株濒危的一个重要原因[5]。海南龙血树Dracaena cambodiana种子萌发对果皮、温度、光照强度等微环境的依赖性较强,原有生境破坏导致种子萌发微环境的改变可能是其处于濒危状态的主要原因之一[6]。羽叶丁香Syringa pinnatifolia致濒的内部原因是其种子含有水溶性萌发抑制物,种群缺少实生幼苗[7]。珙桐Davidia involucrata、翅果油树Elaeagnus mollis、裂叶沙参Adenophora lobophylla等濒危植物的致濒机制主要是其果实的特殊构造或果实构造在萌发中不占优势[8—13]。因此,在保护珍稀濒危植物工作中,开展种子休眠和生态学特性研究有重要意义。
种子繁殖是被子植物最常规的繁殖方式之一。调查发现,中甸刺玫野外繁殖率低,居群内存在根蘖苗和小幼苗,说明其繁殖大部分是通过根蘖苗的形式来完成,实生苗繁殖较少[4]。为较好地利用中甸刺玫这一珍稀濒危植物,本文从种子结构、种子内抑制物活性等方面探索其休眠特性,探索中甸刺玫种子休眠的原因、类型及破解方法,从种子生物学角度了解中甸刺玫的生态适应机制及濒危原因,为种群的保护、恢复和可持续利用提供理论数据。
1 材料与方法
1.1 材料
中甸刺玫种子采自香格里拉县小中甸镇(27.499°N,99.732°E),该地区位于滇西地区横断山脉和滇中地区红色高原交接处,是印度支那半岛和青藏高原的转折区域。主要受西南季风和南支西风急流的交替控制,形成寒温带季风气候,年平均气温 5.8 ℃,最热月平均气温 11.7~24.1℃,极端最低气温-19.4 ℃,年日照时数1742.9~2186.6 h,年均降雨量268~945 mm,无霜期129~197 d。
2011年10月中旬采集新鲜种子后,自然晾干,人工剥离肉质果壳,放入瓶中室温储藏。研究中甸刺玫种子萌发抑制物选用白菜Brassica rapavar var. pekinensis种子,纯度≥95%,净度≥97%,发芽率≥82%,含水量≤8.0%。
采集香格里拉县与中甸刺玫同一生长海拔(3300~4300 m)的蔷薇科峨眉蔷薇Rosa omeienisis种子,自然晾干,人工剥离果壳,放入瓶中室温储藏。峨眉蔷薇生长于海拔750~4500 m,分布普遍,四川、湖北、陕西、宁夏、甘肃、青海、西藏地区均有发现,属广布种,适应性强,抗病抗逆性强,故选用峨眉蔷薇作为参照研究中甸刺玫种子形态。
1.2 方法
1.2.1 瘦果、种子形态观察 对10个样地的中甸刺玫假果进行取样,每个样地随机取10个假果。在采摘一周后,对其瘦果重量、瘦果直径、果实结籽率、种子直径、千粒重等5个指标进行测量。用相同方法测量峨眉蔷薇的果实和种子。
1.2.2 千粒重测定 参照《林木种子检验规程》(GB 2772-1999)相关规定,采用百粒重法测定种子千粒重,对10个样地的种子进行取样,每个样地随机取100粒种子,并分别用1/1000天平称取100粒种子的重量,测量后取平均值后乘以10。用相同方法测量峨眉蔷薇。
1.2.3 种皮扫描电镜观察 将不经任何处理的干燥种子的正面、立面和剥除胚的种壳粘于样品台上,经喷金镀膜,在扫描电镜下观察种子的种脐、外种皮、内种皮等。置于KYKY-EM 3200扫描电镜观察并拍照,工作电压25 kV。
1.2.4 种子吸水性 将干燥种子,浓硫酸处理3、6、9、12、15、18、21、24、27、30 h的种子,剥离种皮的种子分别放入蒸馏水中,每天用吸水纸吸干种子表面水分后称重,直至恒重。每处理3次重复,取平均值。吸水率(%)=(Wx- W0)/ W0×100%。
1.2.5 果实结籽率 对10个样地的假果进行取样,每个样地随机取10个假果,分别观察种子数量,并记录。用相同方法测量峨眉蔷薇。结籽率=饱满种子数/(空瘪种子数+饱满种子数)。
1.2.6 赤霉素处理对胚的影响 采用不同浓度(100、500、1000 mg·kg-1)赤霉素处理胚,放置于20 ℃恒温箱中,注意保持蛭石湿润,定期观察,以胚根伸长3 mm作为萌发标准,分别记录胚的萌发情况。
1.2.7 果壳、种皮、胚乳粗提物制备与活性测定
1.2.7.1 粗提物制备 随机取种子 30粒,分离肉质果皮、种皮、胚乳,然后将各部分分别用 80%甲醇研碎、浸提、过滤之后,仍用80%甲醇浸提残渣,重复3次,合并滤液,减压浓缩蒸干,用蒸馏水洗脱,定容至5 mL。
1.2.7.2 粗提物对白菜籽萌发的影响 用清水和不同浓度浸提液25 ℃下浸泡白菜种子2 h,然后每培养皿放入40粒种子,在25 ℃光照培养箱中进行萌发培养,48 h后统计白菜籽发芽率,重复3次。
1.2.8 数据分析 用Microsoft Excel 2010软件统计数据,采用SPSS 19.0软件进行处理间多重比较分析。
2 结果与分析
2.1 种子形态与结籽率
中甸刺玫瘦果呈肉质,黄绿色,瘦果平均重量82.53 g,平均直径3.51 cm。种子表皮多数为浅褐色或褐色,少数呈黑色的种子不具胚或胚未发育完全。种子长7.54 mm,宽5.02 mm,厚3.96 mm。种子直径为3.51 mm。
中甸刺玫的结籽率为0.69,显著低于峨眉蔷薇的结籽率(0.96)(表1) ,多数果实中没有饱满的种子。结籽率较低可能是导致中甸刺玫濒危的原因之一。
2.2 千粒重
中甸刺玫种子的千粒重为86.37 g(表1)。然而10个样方的种子千粒重在67.07~122.65 g之间,样方间差异较大,这可能与各个样方的土壤、光照等因子的差异有关。

表1 峨眉蔷薇与中甸刺玫种子物理性状特征值Table 1 Physical value of seeds of Rosa praelucens and R. omeienisis
2.3 种皮解剖结构
2.3.1 种子各部分所占比例 中甸刺玫种子的外种皮重量占种子重量的21.38%,内种皮占2.11%,胚占76.42%,胚乳占0.09%。
2.3.2 种皮结构 中甸刺玫扫描电镜观察显示,外种皮光滑且坚硬,由排列紧密的多层厚壁细胞组成的排列紧密的表皮细胞(图2: A),再向内为坚硬致密的纤维状的内种皮,属于明显的栅栏组织,紧紧的包裹着胚(图2: B)。中甸刺玫的种子极面观为圆形(图2: C),赤道面观为椭圆形,外壁表面有细网状纹饰,并且有不规则突起,种脐比较明显。

图2 中甸刺玫种皮显微结构Fig. 2 Seed coat microstructure of Rosa praelucens
2.4 种皮吸水性
图3显示,在25 ℃时,中甸刺玫未处理的种子吸水率较低,吸水量增加不明显,13 d后增加量为18.82%。浓硫酸处理的种子吸水量迅速增加,且处理时间较长的种子吸水量快速增加。吸水24 h后去种皮处理和浓硫酸处理6 h的种子开始迅速吸水。至第3天后去种皮处理种子和浓硫酸处理6 h的种子吸水量基本保持不变,表明种子已经饱和。由此可见,中甸刺玫的种皮透水性较差,对种子有效吸收水分的影响较大,是阻碍种子萌发的因素之一。
2.5 赤霉素处理对胚的影响
用不同浓度赤霉素处理中甸刺玫种胚,从胚的萌发率来看,各浓度处理间存在显著差异。由表2可以看出当赤霉素为100 mg·kg-1时,对胚培养的萌发起到最显著的作用,萌发率为63.87%,说明100 mg·kg-1的赤霉素预处理可以加快胚的萌发速率。
2.6 种子萌发抑制物活性
用中甸刺玫种子各部分粗提液处理白菜种子,观察其48 h后的萌发效果。结果表明,中甸刺玫粗提液对白菜种子萌发的抑制效果为:胚乳>种皮>果壳(表 3)。胚乳粗提液处理的白菜种子萌发率最低,种皮粗提液的抑制作用与果壳粗提液的抑制作用差异不显著,但都显著低于对照。说明中甸刺玫胚乳、种皮、果壳中存在萌发抑制物,与其种子休眠有关。
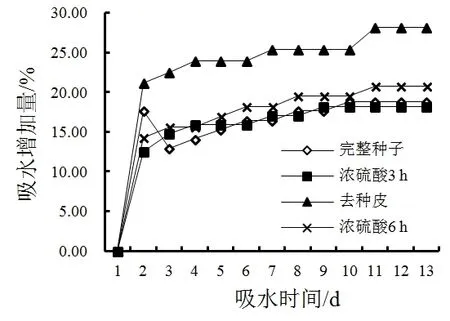
图3 不同处理对种子吸水率的影响Fig. 3 Effect of factor on water permeation

表2 不同浓度赤霉素处理对胚的影响Table 2 Germination of embryo treated with GA3

表3 果实与种子提取液处理的白菜种子萌发率Table 3 Germination rate of Brassica rapavar var.pekinensis seeds treated with methanol extracts of Rosa praelucens fruits and seeds
3 讨论
3.1 种子休眠类型
种子的休眠现象广泛存在于植物当中,Baskin等[16]将种子休眠划分为5种类型,即生理休眠、形态休眠、形态生理休眠、物理休眠和综合休眠。在本研究中,种子吸水试验显示,通过机械损伤预处理种子,在2 d内能明显提高种子的吸水率,可以达到种子萌发所需的含水量,完整种子在7 d内仍然无法达到种子萌发所需含水量。通过电镜观察,中甸刺玫外种皮光滑且十分坚硬,由多层排列紧密的厚壁细胞组成,内种皮坚硬致密,纤维状,属于明显的栅栏组织,紧紧地包裹着胚。物理休眠是由种皮或者果皮中一层或者多层不透水的栅栏细胞所引起的。机械或者化学损伤可以解除物理休眠[6]。因此,种皮不透水性是导致中甸刺玫种子萌发困难的原因之一,可能属于物理休眠和综合休眠。
生理休眠是最普遍的休眠类型,广泛存在于裸子植物和大多数被子植物的种子中[6]。中甸刺玫种皮、果壳、胚乳的甲醇粗提取液均对白菜籽萌发有明显的抑制作用,说明存在抑制种子萌发的内源抑制物,抑制了中甸刺玫种子萌发。
中甸刺玫种子具有物理休眠和生理休眠的特征,属于综合休眠。种皮吸水性较差,100 mg·kg-1赤霉素预处理可以明显提高胚的萌发率。
3.2 种子休眠的生态学意义
种子休眠是其对自然生态环境适应的一种方式[14],对植物的生存、繁殖有着重要的意义[15—16]。中甸刺玫在适应性进化过程中形成的种子休眠与萌发有以下三点意义:其一,中甸刺玫仅生长于云南省香格里拉县小中甸盆地硕多岗河流域内,生长地几乎为平缓的盆地及邻近山麓,果实于9~10月成熟,此时为深秋,土壤水分少,对中甸刺玫的幼苗生长极为不利,种子以休眠的方式度过即将到来的寒冷冬季[4]。为适应该环境,中甸刺玫种子在翌年春夏季温度上升且雨水充沛时解除休眠萌发,为种群补充后代。其二,中甸刺玫果实成熟时已至深秋,为旱季,种子难以完成后熟过程;另外,中甸刺玫果实为猪喜食[4],为适应该情况,肉质果表面有刺,种子形成坚硬致密的内外种皮保护胚和胚乳,其外种皮过于坚硬导致胚难以充分吸水,这可能是中甸刺玫致濒的重要原因。其三,中甸刺玫生长于高海拔环境,昆虫多样性和活动能力降低,有性繁殖的几率降低,而且种子萌发困难[5],为适应该环境,中甸刺玫主要是通过根孽苗方式进行营养繁殖。
近年来,在中甸刺玫分布的小中甸区域,人口迅速增加,道路、房屋修建以及水电站等建设对其生境和种群造成了严重的影响。中甸刺玫虽然进化出一系列适应恶劣生境的对策,但由于分布狭窄、数量稀少,导致种群数量越来越少。种子萌发困难,种群缺乏实生苗,可能降低基因交流、物种进化的几率,增加物种淘汰的可能性。
