鲟鱼营养研究进展(一)
2019-04-26魏洪城薛敏
译/魏洪城 薛敏

北京市鲟鱼、鲑鳟鱼创新团队专栏

鲟鱼(Sturgeons)是一种具有重要生物学和经济意义的鱼类,具有体型大、性成熟晚、产卵期长、寿命长等特点,这些独特的生物学特性使它们容易受过度捕捞、栖息地和产卵场退化、水体和沉积物的污染的影响,因此大多数属于濒危物种。本文将已有的关于鲟鱼营养需求研究进行了概述,包括鲟鱼对能量、蛋白质、脂类、碳水化合物、维生素和矿物质的需求和利用,目的是希望通过这项工作,为今后的研究提供参考,使这些重要鱼类资源得以持续合理地管理和生产。未来的研究,迫切需要开发营养成分合适的饲料,以满足鲟鱼的生长需要;其次,应该更加系统的分析不同实验以及不同鲟鱼物种间的各种联系;最后,应将传统的生长生化试验和现代分子技术、计算机建模等结合,在生物学的各个层次上,采用多因素和多数据的系统方法进行试验设计。对鲟鱼营养需求和利用的机制更科学全面的了解,将有助于我们更好地管理野生鲟鱼资源,利于鲟鱼养殖业更持续高效地发展。
一、引言
鲟鱼是兼具重要生物学(进化、地理、形态、解剖和生理)和经济学(生态、娱乐和水产养殖)价值的物种,属于脊索门,硬骨鱼纲,辐鳍亚纲,鲟形目,鲟科。共有27个物种,其中4种已经灭绝(Birstein, 1993),现存23种分为4个属,其中鳇属2种,铲鲟属2种,拟铲鲟属3种,鲟属16种(Scott and Crossman, 1973)。这些鱼类于2亿5000万年前在侏罗纪时期进化而成,被认为是遗传学“活化石”(Birstein, 1993;Billard and Lecointre, 2001)。鲟鱼保持了许多原始特征,如异形尾鳍、软骨骨架、1根脊索和5排纵行的骨板(鳞)。鲟鱼十分独特,具有很高的杂交能力,分布在同区域的所有物种几乎都能杂交(Billard and Lecointre, 2001),主要生活在北半球的温带水域(从亚热带到亚北极)。有些在海水或咸水中生长和性成熟,但迁移到淡水中产卵,而另一些整个生命周期都在淡水中度过(Billard and Lecointre, 2001)。
欧洲鳇是最大的淡水鱼,生活在黑海、里海、阿波罗海中,最大重量可达1000公斤,生活在威海的铲鲟(P. kaufmanni, P. hermanni, and P.fedtschenkoi)则是最小的鲟鱼,最大重量仅有0.5公斤(Doroshov, 1985)。它们身体通常呈圆柱形,鼻子坚硬凸出,嘴长在腹部可伸出且有触须(Billard and Lecointre, 2001)。它们是优秀的底部猎食者,在鼻子的下侧有非常敏感的触须,用来探测底部动物,长而突出的嘴则是吸食猎物的利器。鲟鱼的胃肠道也是非常独特,它们的幽门异常肥大进化成一个类似胃的器官,成年鲟鱼肠道有功能性纤毛上皮,它们的后肠进化成了螺旋状(Buddington and Doroshov, 1986)。
超过80%的鲟鱼物种濒危或濒临灭绝(IUCN Red List, 2017),因为野生鲟鱼的性成熟周期(3年到20年)和产卵周期(2年到5年以上)都比较长(Birstein, 1993;Billard and Lecointre,2001),此外,它们较大的体型(3kg~2000kg)和较长的寿命(3年~100年)容易被过度捕捞,加之生境和产卵场地的退化,水和沉积物的污染(Billard and Lecointre, 2001)。
近年来,许多鲟鱼经过人工培育释放到野外以增加自然种群数量,或者通过水产养殖供人类消费,以减轻其野生种群的捕捞压力(Billard and Lecointre, 2001;Bronzi et al., 2011)。目前,尽管鲟鱼养殖业受到了广泛关注,但鲟鱼的营养需求和利用,特别是鲟鱼常规营养(表1)的信息仍然很少,然而这些信息对于鲟鱼的成功养殖又具有重要意义。因此,本文的目的主要是概括目前的研究现状并提出一些建议(Hung, 1988, 1991a,b;M edale et al., 1995; Hung and Deng,2002; García-Gallego et al.,2009),为今后的研究提供参考,使这些重要的鱼类资源能够得到持续高效的管理和生产。
二、能量
目前仅有的一些能量研究是Medale and Kaushik(1991),Medale et al.(1991), Cui et al.(1996)和Guo et al.(2012)报道了鲟鱼能量收支的关系。Medale and Kaushik(1991)分别研究了40g、230g和1500g西伯利亚鲟对饲料的能量利用率,试验饲料中含50%粗蛋白、11%脂肪和21.5kJ/g能量,作者发现随着体重的增加,西伯利亚鲟的摄食率减少了,在1500g鲟鱼体内,能量保存率(55%体脂)高于40g和230g的鲟鱼,其能量主要保留在蛋白质内,鲟鱼禁食4周,内源氮每天损失为60mg/kgBW。正如作者所指出的,本研究的局限是比较捕杀和间接量热法之间的误差。如Gisbert et al.(2001)的研究,间接量热法应该和比较捕杀一样应用到将来的鲟鱼能量研究中。
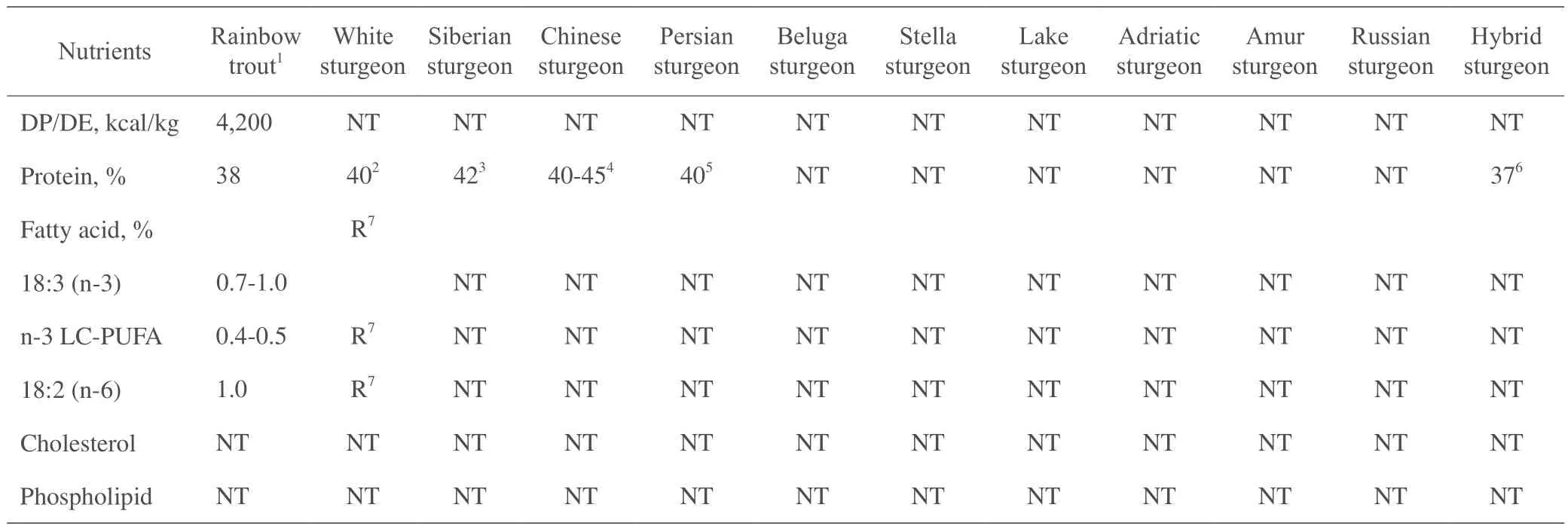
表1 不同鲟鱼的营养需要量
Medale et al.(1991)研究了西伯利亚鲟(初始体重[IBW]49g)对2种饲料(51%粗蛋白和22kJ/g能量)非蛋白能量的利用,两种饲料中分别含有21.8%脂质和9.9%粗淀粉,12.5%脂质和20%的易消化淀粉。8周生长试验后,饲喂含21.8%脂质和9.9%粗淀粉的饲料处理组特定生长率(SGR)和饲料效率(FE)低于另外一组,SGR=(lnFBW-lnIBW)/Days,IBW and FBW分别为初体重和末体重,FE(%)=100×体重增量/饲喂量。作者认为,含有低脂肪和较高可消化碳水化合物的饲料处理组,能量吸收更高,能量吸收的增加是由于鲟鱼体内脂肪的增加,鲟鱼粪便的能量也更高,而2个处理鲟鱼尿和鳃的能量没有明显差异。这些结果表明,在西伯利亚鲟饲料中,除饲料蛋白外,饲料脂质是比碳水化合物更优质的能量物质。这项研究使用的2种饲料,不仅脂质含量不同,还用了不同含量的2种不同淀粉,因此,得出的结论往往比较笼统。未来的研究应该采用单因素饲料脂肪、蛋白质或者碳水化合物剂量梯度实验方法,如Guo et al.(2012)用到的实验方法,便于明确鲟鱼饲料的各能量组分和表型之间的因果关系。

Cui et al.(1996)研究了饲料对不同体重(2.4g~22.5g)高首鲟的影响。据作者报道,无论鲟鱼体重多少,SGR随着饲喂量的增加而线性增加,并且FE在饱食投喂时最高,饱食投喂时,鲟鱼SGR随个体的增大而降低,吸收的能量在进食总能量的94.8%~96.4%之间,尿和鳃能量也随饲喂量的增加而降低,吸收的能量64.9%用于新陈代谢,35.1%用于生长。然而,由于在实验室环境中培养的高首鲟使用活水丝蚓饲喂,具有连续进食的独特摄食习惯(Cui et al., 1997),使得进食能量难以精确计算,不能控制进食能量和蛋白质的比例,这让实验具有一定的局限性。此外,如作者所言,鲟鱼生长与饲喂水丝蚓的线性关系和鲟鱼生长与饲喂人工饲料的曲线关系不同(Hung and Lutes, 1987),这种差异进一步扩大了研究的复杂性。因此在鲟鱼实际养殖中,有必要对饲料能量准确预算,未来的研究应该使用含有不同蛋白和脂质水平且在水中更稳定的饲料,如Hung et al.(1998)的研究。
Guo et al.(2012)研究了7种蛋白质水平的饲料对初重25g的杂交鲟(西伯利亚鲟×俄罗斯鲟)能量收支的影响。8周生长试验表明,当饲料蛋白质从25%增加到40%,SGR随之增加,试验鱼吸收的能量随饲料蛋白质的增加而降低,但是鱼体蛋白沉积随饲料蛋白质的增加而增加。另一方面,尿能量,作为摄入能量的一部分,随着饲料蛋白质水平的增加而增加。作者根据实验结果提出了杂交鲟的能量收支公式:摄入能量(100%)=粪便能量(35.08%)+尿能量(4.53%)+代谢能(34.1%)+生长净能(26.23%)。作者还建议,在使用高蛋白饲料时,应考虑到经济和环境的影响。然而,鲟鱼能量学研究目前只有3个,需要更多的相关研究,以更好地了解鲟鱼能量利用的生物学机制。
